
Identifying Meteorologic and Oceanic Conditions Contributing to a Unique Phytoplankton Bloom Occurrence in Micronesia during October 2013

Abstract
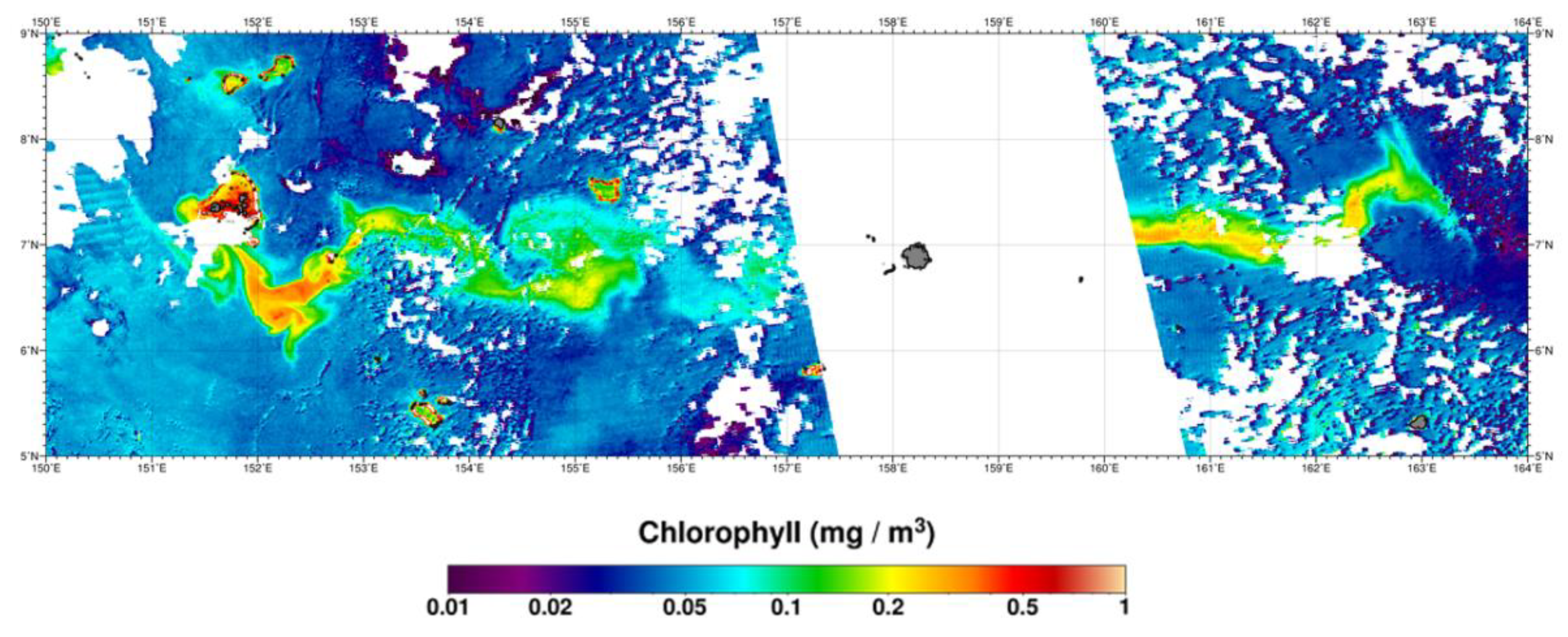
:1. Introduction
2. Materials and Methods
- Japanese Meteorological Agency (JMA) Atmospheric Motion Vectors;
- European Organisation for the Exploitation of Meteorological Satellites (EUMETSAT) Atmospheric Motion Vectors;
- National Oceanic and Atmospheric Administration (NOAA) Geostationary Operational Environmental Satellite (GOES) Atmospheric Motion Vectors;
- NOAA/EUMETSAT Advanced Very High-Resolution Radiometer (AVHRR) Atmospheric Motion Vectors;
- MODIS winds;
- Special Sensor Microwave Imager (SSM/I) and Special Sensor Microwave Imager/Sounder (SSMIS) Wind Speed—the use of these data ended on 29 October 2013;
- Advanced Scatterometer (ASCAT) surface winds.
3. Results
3.1. MERRA-2 Wind Data and SSkT Data
3.2. MERRA-2 Windspeed Variability Filter Test
- (a)
- “Trigger” event: windspeed exceeding 9 m/s;
- (b)
- “Sustainer” event: windspeed exceeding 8 m/s at least 5 days after the initiation “trigger” event;
- (c)
- “Lull/Zero” event; windspeed less than 2 m/s, with the average of the following 6 or more hours less than 1 m/s;
- (d)
- “Peak” event; windspeed exceeding 10 m/s. Note that the windspeed of some “Trigger” events is higher than the “Peak” event—this nomenclature was used because this event in the sequence we examined was both the highest windspeed and had a short duration.
3.3. MERRA-2 Wind Direction Data
4. Discussion
4.1. Description of Wind Event Sequence
4.2. Duration of Bloom
4.3. Significance of Wind Direction
4.4. Examination of Cloud Cover
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behrenfeld, M.J.; Boss, E.S. Student’s tutorial on bloom hypotheses in the context of phytoplankton annual cycles. Glob. Change Biol. 2018, 24, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Brenes, C.L.; Coen, J.E.; Chelton, D.B.; Enfield, D.B.; Leon, S.; Ballestero, D. Wind driven upwelling in the Gulf of Nicoya, Costa Rica. Int. J. Remote Sens. 2003, 24, 1127–1133. [Google Scholar] [CrossRef]
- Kubryakov, A.A.; Zatsepin, A.G.; Stanichny, S.V. Anomalous summer-autumn phytoplankton bloom in 2015 in the Black Sea caused by several strong wind events. J. Marine Sys. 2019, 194, 11–24. [Google Scholar] [CrossRef]
- Stanichny, S.V.; Kubryakova, E.A.; Kubryakov, A.A. Quasi-tropical cyclone caused anomalous autumn coccolithophore bloom in the Black Sea. Biogeosciences 2021, 18, 3173–3188. [Google Scholar] [CrossRef]
- Chow, C.H.; Cheah, W.; Tai, J.-H.; Liu, S.-F. Anomalous wind triggered the largest phytoplankton bloom in the oligotrophic North Pacific Subtropical Gyre. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef]
- Chen, X.; Pan, D.; Bai, Y.; He, X.; Chen, C.-T.A.; Hao, Z. Episodic phytoplankton bloom events in the Bay of Bengal triggered by multiple forcings. Deep Sea Res. Part I Oceanogr. Res. Pap. 2013, 73, 17–30. [Google Scholar] [CrossRef]
- Furnas, M.J.; Mitchell, A.W.; Gilmartin, M.; Revelante, N. Phytoplankton biomass and primary production in semi-enclosed reef lagoons of the central Great Barrier Reef, Australia. Coral Reefs 1990, 9, 1–10. [Google Scholar] [CrossRef]
- Gelaro, R.; McCarty, W.; Suarez, M.J.; Todling, R.; Molod, A.; Takacs, L.; Randles, C.A.; Darmenov, A.; Bosilovich, M.G.; Reichle, R.; et al. The Modern-Era Retrospective Analysis for Research and Applications, Version 2 (MERRA-2). J. Clim. 2017, 30, 5419–5454. [Google Scholar] [CrossRef] [PubMed]
- NASA. Geospatial Interactive Online Visualization ANd aNalysis Infrastructure (Giovanni). Available online: https://giovanni.gsfc.nasa.gov (accessed on 15 June 2022).
- McCarty, W.; Coy, L.; Gelaro, R.; Huang, A.; Merkova, D.; Smith, E.B.; Sienkiewicz, M.; Wargan, K. MERRA-2 Input Observations: Summary and Assessment; Technical Report Series on Global Modeling and Data Assimilation; NASA/TM–2016-104606/; Randal, D.K., Ed.; NASA: Greenbelt, MD, USA, 2016; Volume 46, p. 64.
- Akella, S.; Todling, R.; Suarez, M. Assimilation for skin SST in the NASA GEOS atmospheric data assimilation system. Q. J. R. Meteorol. Soc. 2017, 143, 1032–1046. [Google Scholar] [CrossRef] [PubMed]
- Donlon, C.J.; Martin, M.; Stark, J.; Roberts-Jones, J.; Fiedler, E.; Wimmer, W. The operational sea surface temperature and sea ice analysis (OSTIA) system. Remote Sens. Environ. 2012, 116, 140–158. [Google Scholar] [CrossRef]
- World Weather Online. Available online: https://www.worldweatheronline.com/ (accessed on 24 June 2022).
- SeaDAS: The Official NASA/OB.DAAC Data Analysis Software. Available online: https://seadas.gsfc.nasa.gov/ (accessed on 26 August 2022).
- NASA Worldview. Available online: https://worldview.earthdata.nasa.gov (accessed on 23 August 2021).
- Joint Typhoon Warning Center. Available online: https://www.metoc.navy.mil/jtwc/jtwc.html (accessed on 19 July 2021).
- Joint Typhoon Warning Center Annual Tropical Cyclone Report 2013, “Typhoon Danas”, Page 39. Available online: https://www.metoc.navy.mil/jtwc/products/atcr/2013atcr.pdf (accessed on 19 July 2021).
- National Weather Service Climate Prediction Center, “Cold and Warm Episodes by Season”. Available online: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (accessed on 19 July 2021).
- Acker, J.; Lyon, P.; Hoge, F.; Shen, S.; Roffer, M.; Gawlikowski, G. Interaction of Hurricane Katrina with optically complex water in the Gulf of Mexico: Interpretation using satellite-derived inherent optical properties and chlorophyll concentration. IEEE Geosci. Rem. Sens. Lett. 2009, 6, 209–213. [Google Scholar] [CrossRef]
- Sunda, W. Feedback interactions between trace metal nutrients and phytoplankton in the ocean. Front. Microbiol. 2012, 3, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bif, M.B.; Siqueira, L.; Hansell, D.A. Warm events induce loss of resilience in organic carbon production in the northeast Pacific Ocean. Glob. Biogeochem. Cycles 2019, 33, 1174–1186. [Google Scholar] [CrossRef]
- Bosilovich, M.G. MERRA-2: Initial Evaluation of the Climate. NASA/TM2015104606. 2015; Volume 43, p. 139. Available online: https://gmao.gsfc.nasa.gov/pubs/docs/Bosilovich803.pdf (accessed on 11 August 2022).
- Image of Chuuk Lagoon, Used Under a Creative Commons License Granted by Island Research & Education Initiative (iREi) and Water and Environmental Research Institute of the Western Pacific (WERI). The Dataset was Extracted from the 2020 General Bathymetric Chart of the Oceans (GEBCO) by the GEBCO Compilation Group (2020) as Part of the Seabed 2030 Project. Known as the GEBCO 2020 Grid, this is a Continuous Terrain Model for Ocean and Land with a Spatial Resolution of 15 Arc Seconds. It is an Updated Version of Previous GEBCO Grids, First Published in January 2009. Island Research & Educa-tion Initiative (iREi) (2020) Extracted a Subset of the Original Dataset and Formatted it for Use in this Atlas. Available online: https://islandatlas.org/chuuk/reefs/ (accessed on 26 August 2022).
- Kahru, M.; Gille, S.T.; Murtugudde, R.; Strutton, P.G.; Manzano-Sarabia, M.; Wang, H.; Mitchell, B.G. Global correlations between winds and ocean chlorophyll. J. Geophys. Res. Atmos. 2010, 115, 1–11. [Google Scholar] [CrossRef]
- Wolanski, E.; Colin, P.; Naithani, J.; Deleersnijder, E.; Golbuu, Y. Large amplitude, leaky, island-generated, internal waves around Palau, Micronesia. Estuar. Coast. Shelf Sci. 2004, 60, 705–716. [Google Scholar] [CrossRef]
- Simpson, J.H.; Tett, P.B. Island stirring effects on phytoplankton growth. In Tidal Mixing and Plankton Dynamics; Bowman, J., Yentsch, M., Peterson, W.T., Eds.; Springer: Berlin, Germany, 2012; Volume 17, pp. 41–77. [Google Scholar]
- Simpson, J.H.; Tett, P.B.; Argotte-Spinoza, M.L.; Edwards, M.; Jones, K.J.; Savidge, G. Mixing and phytoplankton growth around an island in a stratified sea. Cont. Shelf Res. 1982, 1, 15–31. [Google Scholar] [CrossRef]
- Messié, M.; Petrenko, A.; Doglioli, A.M.; Martinez, E.; Alvain, S. Basin-scale biogeochemical and ecological impacts of islands in the tropical Pacific Ocean. Nat. Geosci. 2022, 15, 469–474. [Google Scholar] [CrossRef]
- Acker, J.; Spatial and Temporal Coherence of SeaWiFS Chlorophyll Concentration Anomalies in the North Atlantic Bloom (1998–2005) Examined with Giovanni. Proc. Amer. Geophysical Union Spring Meeting, Baltimore, Maryland. 2006. Available online: https://docserver.gesdisc.eosdis.nasa.gov/public/project/publications/01_jan06_AGU_2006_poster_anomalies_1_acker.jpg (accessed on 26 August 2022).
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Start Date | Trigger Wind Speed (m/s) |
|---|---|
| 15 Jun 1982 20:30 | 10.5 |
| 15 Oct 1983 15:30 | 9.4 |
| 23 Aug 1986 0:30 | 9.4 |
| 7 Sep 1991 13:30 | 9.6 |
| 1 Nov 1992 1:30 | 13.2 |
| 1 Mar 1993 8:30 | 10.8 |
| 15 Oct 1994 3:30 | 9.3 |
| 9 Dec 1994 20:30 | 10.2 |
| 12 Nov 1995 16:30 | 9.2 |
| 8 Mar 1997 20:30 | 9.2 |
| 20 Aug 1997 14:30 | 9.2 |
| 8 Dec 2001 12:30 | 9.5 |
| 1 Feb 2004 21:30 | 9.0 |
| 13 Mar 2004 2:30 | 10.9 |
| 7 Apr 2004 23:30 | 9.0 |
| 8 Jun 2004 10:30 | 9.0 |
| 2 Jan 2005 17:30 | 10.2 |
| 16 Sep 2013 8:30 | 9.1 |
| 10 Jan 2014 11:30 | 9.1 |
| 10 Jun 2015 8:30 | 10.4 |
| 28 Jan 2017 0:30 | 9.3 |
| 19 Nov 2018 19:30 | 9.6 |
| 3 Jun 2019 07:30 | 10.3 |
| 22 Nov 2019 18:30 | 9.5 |
| Local Time | Average Wind Speed (km/h: m/s) | Gust Speed (km/h: m/s) |
|---|---|---|
| 0000 | 6: 1.7 | 9: 2.5 |
| 0300 | 7: 1.9 | 10: 2.8 |
| 0600 | 13: 3.6 | 25: 6.9 |
| 0900 | 4: 1.1 | 5: 1.4 |
| 1200 | 4: 1.1 | 5: 1.4 |
| 1500 | 3: 0.8 | 4: 1.1 |
| 1800 | 1: 0.3 | 2: 0.6 |
| 2100 | 4: 1.1 | 6: 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acker, J.; Hunzinger, A.; Kuring, N. Identifying Meteorologic and Oceanic Conditions Contributing to a Unique Phytoplankton Bloom Occurrence in Micronesia during October 2013. Earth 2022, 3, 951-974. https://doi.org/10.3390/earth3030056
Acker J, Hunzinger A, Kuring N. Identifying Meteorologic and Oceanic Conditions Contributing to a Unique Phytoplankton Bloom Occurrence in Micronesia during October 2013. Earth. 2022; 3(3):951-974. https://doi.org/10.3390/earth3030056
Chicago/Turabian StyleAcker, James, Alexis Hunzinger, and Norman Kuring. 2022. "Identifying Meteorologic and Oceanic Conditions Contributing to a Unique Phytoplankton Bloom Occurrence in Micronesia during October 2013" Earth 3, no. 3: 951-974. https://doi.org/10.3390/earth3030056
APA StyleAcker, J., Hunzinger, A., & Kuring, N. (2022). Identifying Meteorologic and Oceanic Conditions Contributing to a Unique Phytoplankton Bloom Occurrence in Micronesia during October 2013. Earth, 3(3), 951-974. https://doi.org/10.3390/earth3030056