
Gut Microbiota and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD): Emerging Pathogenic Mechanisms and Therapeutic Implications


 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Gut Microbiota Dysbiosis and Microbial Signatures Associated with MASLD
3. Mechanisms of Microbiome Modulation
4. Microbiota-Based Biomarkers and Therapeutic Implications
5. Future Perspectives and Research Needs to Exploit/Target the Gut Microbiota in the Context of MASLD
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the Management of Non-Alcoholic Fatty Liver Disease. Obes. Facts 2016, 9, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Q.; Singal, A.G.; Kono, Y.; Tan, D.J.H.; El-Serag, H.B.; Loomba, R. Changing global epidemiology of liver cancer from 2010 to 2019: NASH is the fastest growing cause of liver cancer. Cell Metab. 2022, 34, 969–977.e2. [Google Scholar] [CrossRef] [PubMed]
- Riazi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2022, 7, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. Hepatology 2023, 78, 1966–1986. [Google Scholar] [CrossRef] [PubMed]
- Shea, S.; Lionis, C.; Kite, C.; Atkinson, L.; Lagojda, L.; Chaggar, S.S.; Kyrou, I.; Randeva, H.S. Challenges in the Management of Non-Alcoholic Fatty Liver Disease (NAFLD): Towards a Compassionate Approach. Livers 2023, 3, 434–447. [Google Scholar] [CrossRef]
- Lonardo, A.; Bril, F.; Caldwell, S.H.; Eslam, M.; Fan, J.G.; Gish, R.G.; Gronbaek, H.; Sanal, M.G.; Stefan, N.; Suzuki, A.; et al. Researchers call for more flexible editorial conduct rather than abruptly adopting only the new MASLD nomenclature. J. Hepatol. 2024, 80, e192–e194. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Paik, J.M.; Stepanova, M.; Ong, J.; Alqahtani, S.; Henry, L. Clinical profiles and mortality rates are similar for metabolic dysfunction-associated steatotic liver disease and non-alcoholic fatty liver disease. J. Hepatol. 2024, 80, 694–701. [Google Scholar] [CrossRef]
- Abdelhameed, F.; Kite, C.; Lagojda, L.; Dallaway, A.; Chatha, K.K.; Chaggar, S.S.; Dalamaga, M.; Kassi, E.; Kyrou, I.; Randeva, H.S. Non-invasive Scores and Serum Biomarkers for Fatty Liver in the Era of Metabolic Dysfunction-associated Steatotic Liver Disease (MASLD): A Comprehensive Review from NAFLD to MAFLD and MASLD. Curr. Obes. Rep. 2024, 13, 510–531. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Mejia, M.M.; Jimenez-Gutierrez, C.; Eslam, M.; George, J.; Mendez-Sanchez, N. Breaking new ground: MASLD vs. MAFLD-which holds the key for risk stratification? Hepatol. Int. 2024, 18, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Wong, V.W. Implications of the new nomenclature of steatotic liver disease and definition of metabolic dysfunction-associated steatotic liver disease. Aliment. Pharmacol. Ther. 2024, 59, 150–156. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, B. Unraveling the pathogenesis of non-alcoholic fatty liver diseases through genome-wide association studies. J. Gastroenterol. Hepatol. 2023, 38, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Vilar, L.; Oliveira, C.P.; Faintuch, J.; Mello, E.S.; Nogueira, M.A.; Santos, T.E.; Alves, V.A.; Carrilho, F.J. High-fat diet: A trigger of non-alcoholic steatohepatitis? Preliminary findings in obese subjects. Nutrition 2008, 24, 1097–1102. [Google Scholar] [CrossRef]
- Zelber-Sagi, S.; Nitzan-Kaluski, D.; Goldsmith, R.; Webb, M.; Blendis, L.; Halpern, Z.; Oren, R. Long term nutritional intake and the risk for non-alcoholic fatty liver disease (NAFLD): A population based study. J. Hepatol. 2007, 47, 711–717. [Google Scholar] [CrossRef]
- Rumgay, H.; Murphy, N.; Ferrari, P.; Soerjomataram, I. Alcohol and Cancer: Epidemiology and Biological Mechanisms. Nutrients 2021, 13, 3173. [Google Scholar] [CrossRef] [PubMed]
- Ha, N.B.; Yao, F. Alcohol and Hepatocellular Carcinoma. Clin. Liver Dis. 2024, 28, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Liu, J.; Dalamaga, M. What are the key points in the association between the gut microbiome and nonalcoholic fatty liver disease? Metabol. Open 2019, 1, 9–10. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Obesity and Obesity-Associated Metabolic Disorders: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.G.; Kounatidis, D.; Psallida, S.; Vythoulkas-Biotis, N.; Adamou, A.; Zachariadou, T.; Kargioti, S.; Karampela, I.; Dalamaga, M. NAFLD/MASLD and the Gut-Liver Axis: From Pathogenesis to Treatment Options. Metabolites 2024, 14, 366. [Google Scholar] [CrossRef]
- Vallianou, N.; Christodoulatos, G.S.; Karampela, I.; Tsilingiris, D.; Magkos, F.; Stratigou, T.; Kounatidis, D.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Non-Alcoholic Fatty Liver Disease: Current Evidence and Perspectives. Biomolecules 2021, 12, 56. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Hrncir, T.; Hrncirova, L.; Kverka, M.; Hromadka, R.; Machova, V.; Trckova, E.; Kostovcikova, K.; Kralickova, P.; Krejsek, J.; Tlaskalova-Hogenova, H. Gut Microbiota and NAFLD: Pathogenetic Mechanisms, Microbiota Signatures, and Therapeutic Interventions. Microorganisms 2021, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, A.A.; Zheng, D.; Shibolet, O.; Elinav, E. The role of the microbiome in NAFLD and NASH. EMBO Mol. Med. 2019, 11, e9302. [Google Scholar] [CrossRef]
- Betrapally, N.S.; Gillevet, P.M.; Bajaj, J.S. Gut microbiome and liver disease. Transl. Res. 2017, 179, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Kostovcikova, K.; Coufal, S.; Galanova, N.; Fajstova, A.; Hudcovic, T.; Kostovcik, M.; Prochazkova, P.; Jiraskova Zakostelska, Z.; Cermakova, M.; Sediva, B.; et al. Diet Rich in Animal Protein Promotes Pro-inflammatory Macrophage Response and Exacerbates Colitis in Mice. Front. Immunol. 2019, 10, 919. [Google Scholar] [CrossRef] [PubMed]
- Fajstova, A.; Galanova, N.; Coufal, S.; Malkova, J.; Kostovcik, M.; Cermakova, M.; Pelantova, H.; Kuzma, M.; Sediva, B.; Hudcovic, T.; et al. Diet Rich in Simple Sugars Promotes Pro-Inflammatory Response via Gut Microbiota Alteration and TLR4 Signaling. Cells 2020, 9, 2701. [Google Scholar] [CrossRef] [PubMed]
- Hrncirova, L.; Hudcovic, T.; Sukova, E.; Machova, V.; Trckova, E.; Krejsek, J.; Hrncir, T. Human gut microbes are susceptible to antimicrobial food additives in vitro. Folia Microbiol. 2019, 64, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Hrncirova, L.; Machova, V.; Trckova, E.; Krejsek, J.; Hrncir, T. Food Preservatives Induce Proteobacteria Dysbiosis in Human-Microbiota Associated Nod2-Deficient Mice. Microorganisms 2019, 7, 383. [Google Scholar] [CrossRef]
- Rabot, S.; Membrez, M.; Bruneau, A.; Gerard, P.; Harach, T.; Moser, M.; Raymond, F.; Mansourian, R.; Chou, C.J. Germ-free C57BL/6J mice are resistant to high-fat-diet-induced insulin resistance and have altered cholesterol metabolism. FASEB J. 2010, 24, 4948–4959. [Google Scholar]
- Le Roy, T.; Llopis, M.; Lepage, P.; Bruneau, A.; Rabot, S.; Bevilacqua, C.; Martin, P.; Philippe, C.; Walker, F.; Bado, A.; et al. Intestinal microbiota determines development of non-alcoholic fatty liver disease in mice. Gut 2013, 62, 1787–1794. [Google Scholar] [CrossRef]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, K.; Cohen, T.S. Can You Trust Your Gut? Implicating a Disrupted Intestinal Microbiome in the Progression of NAFLD/NASH. Front Endocrinol. 2020, 11, 592157. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clement, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergstrom, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Backhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Seksik, P.; Furet, J.P.; Firmesse, O.; Nion-Larmurier, I.; Beaugerie, L.; Cosnes, J.; Corthier, G.; Marteau, P.; Dore, J. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm. Bowel Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Stepanova, M.; Ong, J.; Trimble, G.; AlQahtani, S.; Younossi, I.; Ahmed, A.; Racila, A.; Henry, L. Nonalcoholic Steatohepatitis Is the Most Rapidly Increasing Indication for Liver Transplantation in the United States. Clin. Gastroenterol. Hepatol. 2021, 19, 580–589.e585. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat. Commun. 2020, 11, 4982. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Demir, M.; Martin, A.; Jiang, L.; Zhang, X.; Duan, Y.; Gao, B.; Wisplinghoff, H.; Kasper, P.; Roderburg, C.; et al. Intestinal Virome Signature Associated with Severity of Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 159, 1839–1852. [Google Scholar] [CrossRef] [PubMed]
- Demir, M.; Lang, S.; Hartmann, P.; Duan, Y.; Martin, A.; Miyamoto, Y.; Bondareva, M.; Zhang, X.; Wang, Y.; Kasper, P.; et al. The fecal mycobiome in non-alcoholic fatty liver disease. J. Hepatol. 2022, 76, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Zazueta, A.; Valenzuela-Perez, L.; Ortiz-Lopez, N.; Pinto-Leon, A.; Torres, V.; Guinez, D.; Aliaga, N.; Merino, P.; Sandoval, A.; Covarrubias, N.; et al. Alteration of Gut Microbiota Composition in the Progression of Liver Damage in Patients with Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). Int. J. Mol. Sci. 2024, 25, 4387. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, N.; Kamiya, T.; Kawada, N. Recent updates on the role of the gut-liver axis in the pathogenesis of NAFLD/NASH, HCC, and beyond. Hepatol. Commun. 2023, 7, e0241. [Google Scholar] [CrossRef]
- Miele, L.; Valenza, V.; La Torre, G.; Montalto, M.; Cammarota, G.; Ricci, R.; Masciana, R.; Forgione, A.; Gabrieli, M.L.; Perotti, G.; et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology 2009, 49, 1877–1887. [Google Scholar] [CrossRef]
- Volynets, V.; Kuper, M.A.; Strahl, S.; Maier, I.B.; Spruss, A.; Wagnerberger, S.; Konigsrainer, A.; Bischoff, S.C.; Bergheim, I. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD). Dig. Dis. Sci. 2012, 57, 1932–1941. [Google Scholar] [CrossRef]
- Luther, J.; Garber, J.J.; Khalili, H.; Dave, M.; Bale, S.S.; Jindal, R.; Motola, D.L.; Luther, S.; Bohr, S.; Jeoung, S.W.; et al. Hepatic Injury in Nonalcoholic Steatohepatitis Contributes to Altered Intestinal Permeability. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K.; Desai, C.; Iyer, S.S.; Thorn, N.E.; Kumar, P.; Liu, Y.; Smith, T.; Neish, A.S.; Li, H.; Tan, S.; et al. Loss of Junctional Adhesion Molecule A Promotes Severe Steatohepatitis in Mice on a Diet High in Saturated Fat, Fructose, and Cholesterol. Gastroenterology 2016, 151, 733–746.E12. [Google Scholar] [CrossRef]
- Johnson, R.J.; Rivard, C.; Lanaspa, M.A.; Otabachian-Smith, S.; Ishimoto, T.; Cicerchi, C.; Cheeke, P.R.; Macintosh, B.; Hess, T. Fructokinase, Fructans, Intestinal Permeability, and Metabolic Syndrome: An Equine Connection? J. Equine Vet. Sci. 2013, 33, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Spruss, A.; Bergheim, I. Dietary fructose and intestinal barrier: Potential risk factor in the pathogenesis of nonalcoholic fatty liver disease. J. Nutr. Biochem. 2009, 20, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Bergheim, I.; Weber, S.; Vos, M.; Kramer, S.; Volynets, V.; Kaserouni, S.; McClain, C.J.; Bischoff, S.C. Antibiotics protect against fructose-induced hepatic lipid accumulation in mice: Role of endotoxin. J. Hepatol. 2008, 48, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Gabele, E.; Dostert, K.; Hofmann, C.; Wiest, R.; Scholmerich, J.; Hellerbrand, C.; Obermeier, F. DSS induced colitis increases portal LPS levels and enhances hepatic inflammation and fibrogenesis in experimental NASH. J. Hepatol. 2011, 55, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Horst, A.K.; Neumann, K.; Diehl, L.; Tiegs, G. Modulation of liver tolerance by conventional and nonconventional antigen-presenting cells and regulatory immune cells. Cell Mol. Immunol. 2016, 13, 277–292. [Google Scholar] [CrossRef]
- Doherty, D.G. Antigen-specific immune tolerance in the liver. Nat. Biomed. Eng. 2019, 3, 763–765. [Google Scholar] [CrossRef]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARgamma-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Festuccia, W.T.; Crisma, A.R.; Alves, V.S.; Martins, A.R.; Amaral, C.L.; Fiamoncini, J.; Hirabara, S.M.; Sato, F.T.; et al. Tributyrin attenuates obesity-associated inflammation and insulin resistance in high-fat-fed mice. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E272–E282. [Google Scholar] [CrossRef] [PubMed]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhanel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M.; et al. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef]
- Ge, H.; Li, X.; Weiszmann, J.; Wang, P.; Baribault, H.; Chen, J.L.; Tian, H.; Li, Y. Activation of G protein-coupled receptor 43 in adipocytes leads to inhibition of lipolysis and suppression of plasma free fatty acids. Endocrinology 2008, 149, 4519–4526. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406.e1-10. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Hwang, S.J.; O’Donnell, C.J.; Ellison, R.C.; Vasan, R.S.; D’Agostino, R.B., Sr.; Liang, T.J.; Fox, C.S. Parental obesity and offspring serum alanine and aspartate aminotransferase levels: The Framingham heart study. Gastroenterology 2008, 134, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, L.F.; Sun, J.; Singh, B.K.; Zhou, J.; Kaddai, V.A.; Lanni, A.; Yen, P.M.; Sinha, R.A. Short chain fatty acids induce UCP2-mediated autophagy in hepatic cells. Biochem. Biophys. Res. Commun. 2016, 480, 461–467. [Google Scholar] [CrossRef]
- Zhai, S.; Qin, S.; Li, L.; Zhu, L.; Zou, Z.; Wang, L. Dietary butyrate suppresses inflammation through modulating gut microbiota in high-fat diet-fed mice. FEMS Microbiol. Lett. 2019, 366, fnz153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Mehedint, M.G.; Zeisel, S.H. Choline’s role in maintaining liver function: New evidence for epigenetic mechanisms. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 339–345. [Google Scholar] [CrossRef]
- Flessa, C.M.; Nasiri-Ansari, N.; Kyrou, I.; Leca, B.M.; Lianou, M.; Chatzigeorgiou, A.; Kaltsas, G.; Kassi, E.; Randeva, H.S. Genetic and Diet-Induced Animal Models for Non-Alcoholic Fatty Liver Disease (NAFLD) Research. Int. J. Mol. Sci. 2022, 23, 15791. [Google Scholar] [CrossRef]
- Dumas, M.E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511–12516. [Google Scholar] [CrossRef]
- Tang, W.H.; Hazen, S.L. Microbiome, trimethylamine N-oxide, and cardiometabolic disease. Transl. Res. 2017, 179, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Liu, Y.; Zhou, R.F.; Chen, X.L.; Wang, C.; Tan, X.Y.; Wang, L.J.; Zheng, R.D.; Zhang, H.W.; Ling, W.H.; et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci. Rep. 2016, 6, 19076. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Van de Wiele, T.; De Bodt, J.; Marzorati, M.; Gewirtz, A.T. Dietary emulsifiers directly alter human microbiota composition and gene expression ex vivo potentiating intestinal inflammation. Gut 2017, 66, 1414–1427. [Google Scholar] [CrossRef]
- Barrea, L.; Annunziata, G.; Muscogiuri, G.; Di Somma, C.; Laudisio, D.; Maisto, M.; de Alteriis, G.; Tenore, G.C.; Colao, A.; Savastano, S. Trimethylamine-N-oxide (TMAO) as Novel Potential Biomarker of Early Predictors of Metabolic Syndrome. Nutrients 2018, 10, 1971. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.S.; Obeid, S.; Wang, Z.N.; Hazen, B.J.; Li, L.; Wu, Y.P.; Hurd, A.G.; Gu, X.D.; Pratt, A.; Levison, B.S.; et al. Trimethyllysine, a trimethylamine N-oxide precursor, provides near- and long-term prognostic value in patients presenting with acute coronary syndromes. Eur. Heart J. 2019, 40, 2700–2709. [Google Scholar] [CrossRef]
- Boyer, J.L. Bile formation and secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [PubMed]
- Ferslew, B.C.; Xie, G.; Johnston, C.K.; Su, M.; Stewart, P.W.; Jia, W.; Brouwer, K.L.; Barritt, A.S.t. Altered Bile Acid Metabolome in Patients with Nonalcoholic Steatohepatitis. Dig. Dis. Sci. 2015, 60, 3318–3328. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zheng, M.; Liu, J.; Luo, Y.; Yang, W.; Yang, J.; Liu, J.; Zhou, J.; Xu, C.; Zhao, F.; et al. Ratio of Conjugated Chenodeoxycholic to Muricholic Acids is Associated with Severity of Nonalcoholic Steatohepatitis. Obesity 2019, 27, 2055–2066. [Google Scholar] [CrossRef]
- Fiorucci, S.; Biagioli, M.; Zampella, A.; Distrutti, E. Bile Acids Activated Receptors Regulate Innate Immunity. Front. Immunol. 2018, 9, 1853. [Google Scholar] [CrossRef] [PubMed]
- Weickert, M.O.; Hattersley, J.G.; Kyrou, I.; Arafat, A.M.; Rudovich, N.; Roden, M.; Nowotny, P.; von Loeffelholz, C.; Matysik, S.; Schmitz, G.; et al. Effects of supplemented isoenergetic diets varying in cereal fiber and protein content on the bile acid metabolic signature and relation to insulin resistance. Nutr. Diabetes 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Belka, M.; Gostynska-Stawna, A.; Stawny, M.; Krajka-Kuzniak, V. Activation of Nrf2 and FXR via Natural Compounds in Liver Inflammatory Disease. Int. J. Mol. Sci. 2024, 25, 11213. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, L.; Yi, Q.; Luo, L.; Xiong, Y. Regulation of bile acids and their receptor FXR in metabolic diseases. Front. Nutr. 2024, 11, 1447878. [Google Scholar] [CrossRef]
- Tang, Y.; Fan, Y.; Wang, Y.; Wang, D.; Huang, Q.; Chen, T.; Cao, X.; Wen, C.; Shen, X.; Li, J.; et al. A Current Understanding of FXR in NAFLD: The multifaceted regulatory role of FXR and novel lead discovery for drug development. Biomed. Pharmacother. 2024, 175, 116658. [Google Scholar] [CrossRef]
- Wu, D.; van de Graaf, S.F.J. Maladaptive regeneration and metabolic dysfunction associated steatotic liver disease: Common mechanisms and potential therapeutic targets. Biochem. Pharmacol. 2024, 227, 116437. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Tu, W.; Huang, G. Regulating bile acids signaling for NAFLD: Molecular insights and novel therapeutic interventions. Front. Microbiol. 2024, 15, 1341938. [Google Scholar] [CrossRef] [PubMed]
- Almeqdadi, M.; Gordon, F.D. Farnesoid X Receptor Agonists: A Promising Therapeutic Strategy for Gastrointestinal Diseases. Gastro Hep Adv. 2024, 3, 344–352. [Google Scholar] [CrossRef]
- Hu, L.; Cheng, Z.; Chu, H.; Wang, W.; Jin, Y.; Yang, L. TRIF-dependent signaling and its role in liver diseases. Front. Cell Dev. Biol. 2024, 12, 1370042. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhu, L.; Tao, Y.; Lu, W.; Cheng, H. Role of targeting TLR4 signaling axis in liver-related diseases. Pathol. Res. Pract. 2023, 244, 154410. [Google Scholar] [CrossRef]
- Hoyles, L.; Fernandez-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 2018, 24, 1070–1080. [Google Scholar] [CrossRef]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef]
- Yu, Q.; Jiang, Z.; Zhang, L. Bile acid regulation: A novel therapeutic strategy in non-alcoholic fatty liver disease. Pharmacol. Ther. 2018, 190, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Swamikkannu, D.M.; Dasarapu, S.; Siva, R.P.V.; Nallam, J.; Pabba, S. The gut-liver nexus: Exploring gut microbiota dysbiosis in non-alcoholic fatty liver disease and its therapeutic implications. Egypt. Liver J. 2024, 14, 28. [Google Scholar] [CrossRef]
- Ma, J.L.; Zhou, Q.H.; Li, H.K. Gut Microbiota and Nonalcoholic Fatty Liver Disease: Insights on Mechanisms and Therapy. Nutrients 2017, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Yki-Jarvinen, H.; Neuschwander-Tetri, B.A. Metabolic dysfunction-associated steatotic liver disease: Heterogeneous pathomechanisms and effectiveness of metabolism-based treatment. Lancet Diabetes Endocrinol. 2025, 13, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Armandi, A.; Bugianesi, E. Dietary and pharmacological treatment in patients with metabolic-dysfunction associated steatotic liver disease. Eur. J. Intern. Med. 2024, 122, 20–27. [Google Scholar] [CrossRef]
- Ciardullo, S.; Muraca, E.; Vergani, M.; Invernizzi, P.; Perseghin, G. Advancements in pharmacological treatment of NAFLD/MASLD: A focus on metabolic and liver-targeted interventions. Gastroenterol. Rep. 2024, 12, goae029. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, B.; Rao, G.; Tang, Y.; Rodriguez, A.; Glass, L.C.; Hartman, M.L. Incretin-based investigational therapies for the treatment of MASLD/MASH. Diabetes Res. Clin. Pract. 2024, 211, 111675. [Google Scholar] [CrossRef] [PubMed]
- Chianelli, D.; Rucker, P.V.; Roland, J.; Tully, D.C.; Nelson, J.; Liu, X.D.; Bursulaya, B.; Hernandez, E.D.; Wu, J.N.; Prashad, M.; et al. Nidufexor (LMB763), a Novel FXR Modulator for the Treatment of Nonalcoholic Steatohepatitis. J. Med. Chem. 2020, 63, 3868–3880. [Google Scholar] [CrossRef]
- Traussnigg, S.; Halilbasic, E.; Hofer, H.; Munda, P.; Stojakovic, T.; Fauler, G.; Kashofer, K.; Krssak, M.; Wolzt, M.; Trauner, M. Open-label phase II study evaluating safety and efficacy of the non-steroidal farnesoid X receptor agonist PX-104 in non-alcoholic fatty liver disease. Wien. Klin. Wochenschr. 2021, 133, 441–451. [Google Scholar] [CrossRef]
- Patel, K.; Harrison, S.A.; Elkhashab, M.; Trotter, J.F.; Herring, R.; Rojter, S.E.; Kayali, Z.; Wong, V.W.; Greenbloom, S.; Jayakumar, S.; et al. Cilofexor, a Nonsteroidal FXR Agonist, in Patients with Noncirrhotic NASH: A Phase 2 Randomized Controlled Trial. Hepatology 2020, 72, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Lopez, P.; Lawitz, E.J.; Lucas, K.J.; Loeffler, J.; Kim, W.; Goh, G.B.B.; Huang, J.F.; Serra, C.; Andreone, P.; et al. Tropifexor for nonalcoholic steatohepatitis: An adaptive, randomized, placebo-controlled phase 2a/b trial. Nat. Med. 2023, 29, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Ratziu, V.; Loomba, R.; Rinella, M.; Anstee, Q.M.; Goodman, Z.; Bedossa, P.; Geier, A.; Beckebaum, S.; Newsome, P.N.; et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: Interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet 2019, 394, 2184–2196. [Google Scholar] [CrossRef] [PubMed]
- Aishwarya, T.S.; Mounika, N.; Vishwakarma, G.; Adela, R. Effect of obeticholic acid in non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH) patients: A systematic review and meta-analysis. RPS Pharm. Pharmacol. Rep. 2022, 1, rqac001. [Google Scholar] [CrossRef]
- Lin, X.; Mai, M.; He, T.; Huang, H.; Zhang, P.; Xia, E.; Guo, H. Efficiency of ursodeoxycholic acid for the treatment of nonalcoholic steatohepatitis: A systematic review and meta-analysis. Expert. Rev. Gastroenterol. Hepatol. 2022, 16, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Newsome, P.N.; Buchholtz, K.; Cusi, K.; Linder, M.; Okanoue, T.; Ratziu, V.; Sanyal, A.J.; Sejling, A.S.; Harrison, S.A.; Investigators, N.N. A Placebo-Controlled Trial of Subcutaneous Semaglutide in Nonalcoholic Steatohepatitis. N. Engl. J. Med. 2021, 384, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Ling, L.; Beuers, U.; DePaoli, A.M.; Lieu, H.D.; Harrison, S.A.; Hirschfield, G.M. Potent suppression of hydrophobic bile acids by aldafermin, an FGF19 analogue, across metabolic and cholestatic liver diseases. JHEP Rep. 2021, 3, 100255. [Google Scholar] [CrossRef]
- Harrison, S.A.; Ruane, P.J.; Freilich, B.; Neff, G.; Patil, R.; Behling, C.; Hu, C.; Shringarpure, R.; de Temple, B.; Fong, E.; et al. A randomized, double-blind, placebo-controlled phase IIa trial of efruxifermin for patients with compensated NASH cirrhosis. JHEP Rep. 2023, 5, 100563. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Samajdar, S.S.; Das, S. Effects of saroglitazar in the treatment of non-alcoholic fatty liver disease or non-alcoholic steatohepatitis: A systematic review and meta-analysis. Clin. Res. Hepatol. Gastroenterol. 2023, 47, 102174. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Bedossa, P.; Guy, C.D.; Schattenberg, J.M.; Loomba, R.; Taub, R.; Labriola, D.; Moussa, S.E.; Neff, G.W.; Rinella, M.E.; et al. A Phase 3, Randomized, Controlled Trial of Resmetirom in NASH with Liver Fibrosis. N. Engl. J. Med. 2024, 390, 497–509. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Velayudham, A.; Dolganiuc, A.; Ellis, M.; Petrasek, J.; Kodys, K.; Mandrekar, P.; Szabo, G. VSL#3 probiotic treatment attenuates fibrosis without changes in steatohepatitis in a diet-induced nonalcoholic steatohepatitis model in mice. Hepatology 2009, 49, 989–997. [Google Scholar]
- Paolella, G.; Mandato, C.; Pierri, L.; Poeta, M.; Di Stasi, M.; Vajro, P. Gut-liver axis and probiotics: Their role in non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 15518–15531. [Google Scholar] [CrossRef]
- Govender, M.; Choonara, Y.E.; Kumar, P.; du Toit, L.C.; van Vuuren, S.; Pillay, V. A review of the advancements in probiotic delivery: Conventional vs. non-conventional formulations for intestinal flora supplementation. AAPS PharmSciTech 2014, 15, 29–43. [Google Scholar] [PubMed]
- Alisi, A.; Bedogni, G.; Baviera, G.; Giorgio, V.; Porro, E.; Paris, C.; Giammaria, P.; Reali, L.; Anania, F.; Nobili, V. Randomised clinical trial: The beneficial effects of VSL#3 in obese children with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2014, 39, 1276–1285. [Google Scholar] [PubMed]
- Derosa, G.; Guasti, L.; D’Angelo, A.; Martinotti, C.; Valentino, M.C.; Di Matteo, S.; Bruno, G.M.; Maresca, A.M.; Gaudio, G.V.; Maffioli, P. Probiotic Therapy With VSL#3® in Patients With NAFLD: A Randomized Clinical Trial. Front. Nutr. 2022, 9, 846873. [Google Scholar]
- Duseja, A.; Acharya, S.K.; Mehta, M.; Chhabra, S.; Shalimar; Rana, S.; Das, A.; Dattagupta, S.; Dhiman, R.K.; Chawla, Y.K. High potency multistrain probiotic improves liver histology in non-alcoholic fatty liver disease (NAFLD): A randomised, double-blind, proof of concept study. BMJ Open Gastroenterol. 2019, 6, e000315. [Google Scholar] [CrossRef]
- Mohamad Nor, M.H.; Ayob, N.; Mokhtar, N.M.; Raja Ali, R.A.; Tan, G.C.; Wong, Z.; Shafiee, N.H.; Wong, Y.P.; Mustangin, M.; Nawawi, K.N.M. The Effect of Probiotics (MCP® BCMC® Strains) on Hepatic Steatosis, Small Intestinal Mucosal Immune Function, and Intestinal Barrier in Patients with Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 3192. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.L.; Laight, D.; Aspinall, R.J.; Higginson, A.; Cummings, M.H. A randomised placebo controlled trial of VSL#3® probiotic on biomarkers of cardiovascular risk and liver injury in non-alcoholic fatty liver disease. BMC Gastroenterol. 2021, 21, 144. [Google Scholar]
- Silva-Sperb, A.S.; Moraes, H.A.; Barcelos, S.T.A.; de Moura, B.C.; Longo, L.; Michalczuk, M.T.; Cerski, C.T.S.; Uribe-Cruz, C.; da Silveira, T.R.; Alvares-da-Silva, M.R.; et al. Probiotic supplementation for 24 weeks in patients with non-alcoholic steatohepatitis: The PROBILIVER randomized clinical trial. Front. Nutr. 2024, 11, 1362694. [Google Scholar] [CrossRef] [PubMed]
- Bomhof, M.R.; Parnell, J.A.; Ramay, H.R.; Crotty, P.; Rioux, K.P.; Probert, C.S.; Jayakumar, S.; Raman, M.; Reimer, R.A. Histological improvement of non-alcoholic steatohepatitis with a prebiotic: A pilot clinical trial. Eur. J. Nutr. 2019, 58, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Tsigalou, C.; Dalamaga, M. Probiotics, Prebiotics, Synbiotics, Postbiotics, and Obesity: Current Evidence, Controversies, and Perspectives. Curr. Obes. Rep. 2020, 9, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, W.; Rupasinghe, H.P.V. Proanthocyanidins-Based Synbiotics as a Novel Strategy for Nonalcoholic Fatty Liver Disease (NAFLD) Risk Reduction. Molecules 2024, 29, 709. [Google Scholar] [CrossRef] [PubMed]
- Philips, C.A.; Pande, A.; Shasthry, S.M.; Jamwal, K.D.; Khillan, V.; Chandel, S.S.; Kumar, G.; Sharma, M.K.; Maiwall, R.; Jindal, A.; et al. Healthy Donor Fecal Microbiota Transplantation in Steroid-Ineligible Severe Alcoholic Hepatitis: A Pilot Study. Clin. Gastroenterol. Hepatol. 2017, 15, 600–602. [Google Scholar] [CrossRef] [PubMed]
- van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; de Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.; Tijssen, J.G.; et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Kassam, Z.; Fagan, A.; Gavis, E.A.; Liu, E.; Cox, I.J.; Kheradman, R.; Heuman, D.; Wang, J.; Gurry, T.; et al. Fecal microbiota transplant from a rational stool donor improves hepatic encephalopathy: A randomized clinical trial. Hepatology 2017, 66, 1727–1738. [Google Scholar] [CrossRef]
- Kelly, C.R.; Kim, A.M.; Laine, L.; Wu, G.D. The AGA’s Fecal Microbiota Transplantation National Registry: An Important Step Toward Understanding Risks and Benefits of Microbiota Therapeutics. Gastroenterology 2017, 152, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojarvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef]
- Xue, L.; Deng, Z.; Luo, W.; He, X.; Chen, Y. Effect of Fecal Microbiota Transplantation on Non-Alcoholic Fatty Liver Disease: A Randomized Clinical Trial. Front. Cell Infect. Microbiol. 2022, 12, 759306. [Google Scholar] [CrossRef] [PubMed]
- Stols-Goncalves, D.; Mak, A.L.; Madsen, M.S.; van der Vossen, E.W.J.; Bruinstroop, E.; Henneman, P.; Mol, F.; Scheithauer, T.P.M.; Smits, L.; Witjes, J.; et al. Faecal Microbiota transplantation affects liver DNA methylation in Non-alcoholic fatty liver disease: A multi-omics approach. Gut Microbes 2023, 15, 2223330. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Chen, M. Commentary: Effect of fecal microbiota transplantation on non-alcoholic fatty liver disease: A randomized clinical trial. Front. Cell Infect. Microbiol. 2022, 12, 1056394. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, S.; Zhang, L.; Zhao, W.; Qin, Y.; Liu, M. The effect of gut microbiome-targeted therapies in nonalcoholic fatty liver disease: A systematic review and network meta-analysis. Front. Nutr. 2024, 11, 1470185. [Google Scholar] [CrossRef] [PubMed]
- Ayob, N.; Muhammad Nawawi, K.N.; Mohamad Nor, M.H.; Raja Ali, R.A.; Ahmad, H.F.; Oon, S.F.; Mohd Mokhtar, N. The Effects of Probiotics on Small Intestinal Microbiota Composition, Inflammatory Cytokines and Intestinal Permeability in Patients with Non-Alcoholic Fatty Liver Disease. Biomedicines 2023, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.; Won, G.L.; Chim, A.M.; Chu, W.C.; Yeung, D.K.; Li, K.C.; Chan, H.L. Treatment of nonalcoholic steatohepatitis with probiotics. A proof-of-concept study. Ann. Hepatol. 2013, 12, 256–262. [Google Scholar] [CrossRef]
- Sepideh, A.; Karim, P.; Hossein, A.; Leila, R.; Hamdollah, M.; Mohammad, E.G.; Mojtaba, S.; Mohammad, S.; Ghader, G.; Seyed Moayed, A. Effects of Multistrain Probiotic Supplementation on Glycemic and Inflammatory Indices in Patients with Nonalcoholic Fatty Liver Disease: A Double-Blind Randomized Clinical Trial. J. Am. Coll. Nutr. 2016, 35, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Aller, R.; De Luis, D.A.; Izaola, O.; Conde, R.; Gonzalez Sagrado, M.; Primo, D.; De La Fuente, B.; Gonzalez, J. Effect of a probiotic on liver aminotransferases in nonalcoholic fatty liver disease patients: A double blind randomized clinical trial. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1090–1095. [Google Scholar] [PubMed]
- Ahn, S.B.; Jun, D.W.; Kang, B.K.; Lim, J.H.; Lim, S.; Chung, M.J. Randomized, Double-blind, Placebo-controlled Study of a Multispecies Probiotic Mixture in Nonalcoholic Fatty Liver Disease. Sci. Rep. 2019, 9, 5688. [Google Scholar] [CrossRef] [PubMed]
- Escouto, G.S.; Port, G.Z.; Tovo, C.V.; Fernandes, S.A.; Peres, A.; Dorneles, G.P.; Houde, V.P.; Varin, T.V.; Pilon, G.; Marette, A.; et al. Probiotic Supplementation, Hepatic Fibrosis, and the Microbiota Profile in Patients with Nonalcoholic Steatohepatitis: A Randomized Controlled Trial. J. Nutr. 2023, 153, 1984–1993. [Google Scholar] [CrossRef] [PubMed]
- Reshef, N.; Gophna, U.; Reshef, L.; Konikoff, F.; Gabay, G.; Zornitzki, T.; Knobler, H.; Maor, Y. Prebiotic Treatment in Patients with Nonalcoholic Fatty Liver Disease (NAFLD)-A Randomized Pilot Trial. Nutrients 2024, 16, 1571. [Google Scholar] [CrossRef]
- Scorletti, E.; Afolabi, P.R.; Miles, E.A.; Smith, D.E.; Almehmadi, A.; Alshathry, A.; Childs, C.E.; Del Fabbro, S.; Bilson, J.; Moyses, H.E.; et al. Synbiotics Alter Fecal Microbiomes, But Not Liver Fat or Fibrosis, in a Randomized Trial of Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1597–1610.E7. [Google Scholar] [CrossRef]
- Mofidi, F.; Poustchi, H.; Yari, Z.; Nourinayyer, B.; Merat, S.; Sharafkhah, M.; Malekzadeh, R.; Hekmatdoost, A. Synbiotic supplementation in lean patients with non-alcoholic fatty liver disease: A pilot, randomised, double-blind, placebo-controlled, clinical trial. Br. J. Nutr. 2017, 117, 662–668. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Vacante, M.; Antic, T.; Giordano, M.; Chisari, G.; Acquaviva, R.; Mastrojeni, S.; Malaguarnera, G.; Mistretta, A.; Li Volti, G.; et al. Bifidobacterium longum with fructo-oligosaccharides in patients with non alcoholic steatohepatitis. Dig. Dis. Sci. 2012, 57, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Asgharian, A.; Askari, G.; Esmailzade, A.; Feizi, A.; Mohammadi, V. The Effect of Symbiotic Supplementation on Liver Enzymes, C-reactive Protein and Ultrasound Findings in Patients with Non-alcoholic Fatty Liver Disease: A Clinical Trial. Int. J. Prev. Med. 2016, 7, 59. [Google Scholar] [PubMed]
- Abhari, K.; Saadati, S.; Yari, Z.; Hosseini, H.; Hedayati, M.; Abhari, S.; Alavian, S.M.; Hekmatdoost, A. The effects of Bacillus coagulans supplementation in patients with non-alcoholic fatty liver disease: A randomized, placebo-controlled, clinical trial. Clin. Nutr. ESPEN 2020, 39, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Sayari, S.; Neishaboori, H.; Jameshorani, M. Combined effects of synbiotic and sitagliptin versus sitagliptin alone in patients with nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2018, 24, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Witjes, J.J.; Smits, L.P.; Pekmez, C.T.; Prodan, A.; Meijnikman, A.S.; Troelstra, M.A.; Bouter, K.E.C.; Herrema, H.; Levin, E.; Holleboom, A.G.; et al. Donor Fecal Microbiota Transplantation Alters Gut Microbiota and Metabolites in Obese Individuals With Steatohepatitis. Hepatol. Commun. 2020, 4, 1578–1590. [Google Scholar] [CrossRef]
- Craven, L.; Rahman, A.; Nair Parvathy, S.; Beaton, M.; Silverman, J.; Qumosani, K.; Hramiak, I.; Hegele, R.; Joy, T.; Meddings, J.; et al. Allogenic Fecal Microbiota Transplantation in Patients With Nonalcoholic Fatty Liver Disease Improves Abnormal Small Intestinal Permeability: A Randomized Control Trial. Am. J. Gastroenterol. 2020, 115, 1055–1065. [Google Scholar] [CrossRef]
- Abdel-Razik, A.; Mousa, N.; Shabana, W.; Refaey, M.; Elzehery, R.; Elhelaly, R.; Zalata, K.; Abdelsalam, M.; Eldeeb, A.A.; Awad, M.; et al. Rifaximin in nonalcoholic fatty liver disease: Hit multiple targets with a single shot. Eur. J. Gastroenterol. Hepatol. 2018, 30, 1237–1246. [Google Scholar] [CrossRef]
- Neuschwander-Tetri, B.A.; Loomba, R.; Sanyal, A.J.; Lavine, J.E.; Van Natta, M.L.; Abdelmalek, M.F.; Chalasani, N.; Dasarathy, S.; Diehl, A.M.; Hameed, B.; et al. Farnesoid X nuclear receptor ligand obeticholic acid for non-cirrhotic, non-alcoholic steatohepatitis (FLINT): A multicentre, randomised, placebo-controlled trial. Lancet 2015, 385, 956–965. [Google Scholar] [CrossRef]
- Harrison, S.A.; Rinella, M.E.; Abdelmalek, M.F.; Trotter, J.F.; Paredes, A.H.; Arnold, H.L.; Kugelmas, M.; Bashir, M.R.; Jaros, M.J.; Ling, L.; et al. NGM282 for treatment of non-alcoholic steatohepatitis: A multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2018, 391, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Targher, G.; Byrne, C.D.; Cao, Y.Y.; Zheng, M.H. Current status and future trends of the global burden of MASLD. Trends Endocrinol. Metab. 2024, 35, 697–707. [Google Scholar] [CrossRef]
- Fianchi, F.; Liguori, A.; Gasbarrini, A.; Grieco, A.; Miele, L. Nonalcoholic Fatty Liver Disease (NAFLD) as Model of Gut-Liver Axis Interaction: From Pathophysiology to Potential Target of Treatment for Personalized Therapy. Int. J. Mol. Sci. 2021, 22, 6485. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, S.R.; Schnabl, B.; Knight, R.; Loomba, R. Current Concepts, Opportunities, and Challenges of Gut Microbiome-Based Personalized Medicine in Nonalcoholic Fatty Liver Disease. Cell Metab. 2021, 33, 21–32. [Google Scholar] [CrossRef]
- Bourganou, M.V.; Chondorgianni, M.E.; Kyrou, I.; Flessa, C.-M.; Chatzigeorgiou, A.; Oikonomou, E.; Randeva, H.S.; Kassi, E. Unraveling Metabolic Dysfunction-Associated Steatotic Liver Disease Through the Use of Omics Technologies. Int. J. Mol. Sci. 2025, 26, 1589. [Google Scholar] [CrossRef]
- Kyrou, I.; Randeva, H.S.; Tsigos, C.; Kaltsas, G.; Weickert, M.O. Clinical Problems Caused by Obesity. In Endotext [INTERNET]; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. [Google Scholar]
- Chew, N.W.S.; Mehta, A.; Goh, R.S.J.; Zhang, A.; Chen, Y.; Chong, B.; Chew, H.S.J.; Shabbir, A.; Brown, A.; Dimitriadis, G.K.; et al. Cardiovascular-Liver-Metabolic Health: Recommendations in Screening, Diagnosis, and Management of Metabolic Dysfunction-Associated Steatotic Liver Disease in Cardiovascular Disease via Modified Delphi Approach. Circulation 2025, 151, 98–119. [Google Scholar] [CrossRef] [PubMed]
- Puengel, T.; Tacke, F. Pharmacotherapeutic options for metabolic dysfunction-associated steatotic liver disease: Where are we today? Expert. Opin. Pharmacother. 2024, 25, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, A.; Tsioulos, G.; Kassi, E.; Chatzigeorgiou, A. Current and experimental pharmacotherapy for the management of non-alcoholic fatty liver disease. Hormones 2024, 23, 621–636. [Google Scholar] [CrossRef]
- Zhi, Y.; Dong, Y.; Li, X.; Zhong, W.; Lei, X.; Tang, J.; Mao, Y. Current Progress and Challenges in the Development of Pharmacotherapy for Metabolic Dysfunction-Associated Steatohepatitis. Diabetes Metab. Res. Rev. 2024, 40, e3846. [Google Scholar] [CrossRef] [PubMed]
- Ivancovsky Wajcman, D.; Byrne, C.J.; Dillon, J.F.; Brennan, P.N.; Villota-Rivas, M.; Younossi, Z.M.; Allen, A.M.; Crespo, J.; Gerber, L.H.; Lazarus, J.V. A narrative review of lifestyle management guidelines for metabolic dysfunction-associated steatotic liver disease. Hepatology 2024, 10, 1058. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines on the Management of Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). Obes. Facts 2024, 17, 374–444. [Google Scholar] [CrossRef]
- Allen, A.M.; Charlton, M.; Cusi, K.; Harrison, S.A.; Kowdley, K.V.; Noureddin, M.; Shubrook, J.H. Guideline-based management of metabolic dysfunction-associated steatotic liver disease in the primary care setting. Postgrad. Med. 2024, 136, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Zelber-Sagi, S.; Grinshpan, L.S.; Ivancovsky-Wajcman, D.; Goldenshluger, A.; Gepner, Y. One size does not fit all; practical, personal tailoring of the diet to NAFLD patients. Liver Int. 2022, 42, 1731–1750. [Google Scholar] [CrossRef] [PubMed]
- Thakral, N.; Desalegn, H.; Diaz, L.A.; Cabrera, D.; Loomba, R.; Arrese, M.; Arab, J.P. A Precision Medicine Guided Approach to the Utilization of Biomarkers in MASLD. Semin. Liver Dis. 2024, 44, 273–286. [Google Scholar] [CrossRef]
- Mullins, V.A.; Bresette, W.; Johnstone, L.; Hallmark, B.; Chilton, F.H. Genomics in Personalized Nutrition: Can You “Eat for Your Genes”? Nutrients 2020, 12, 3118. [Google Scholar] [CrossRef] [PubMed]
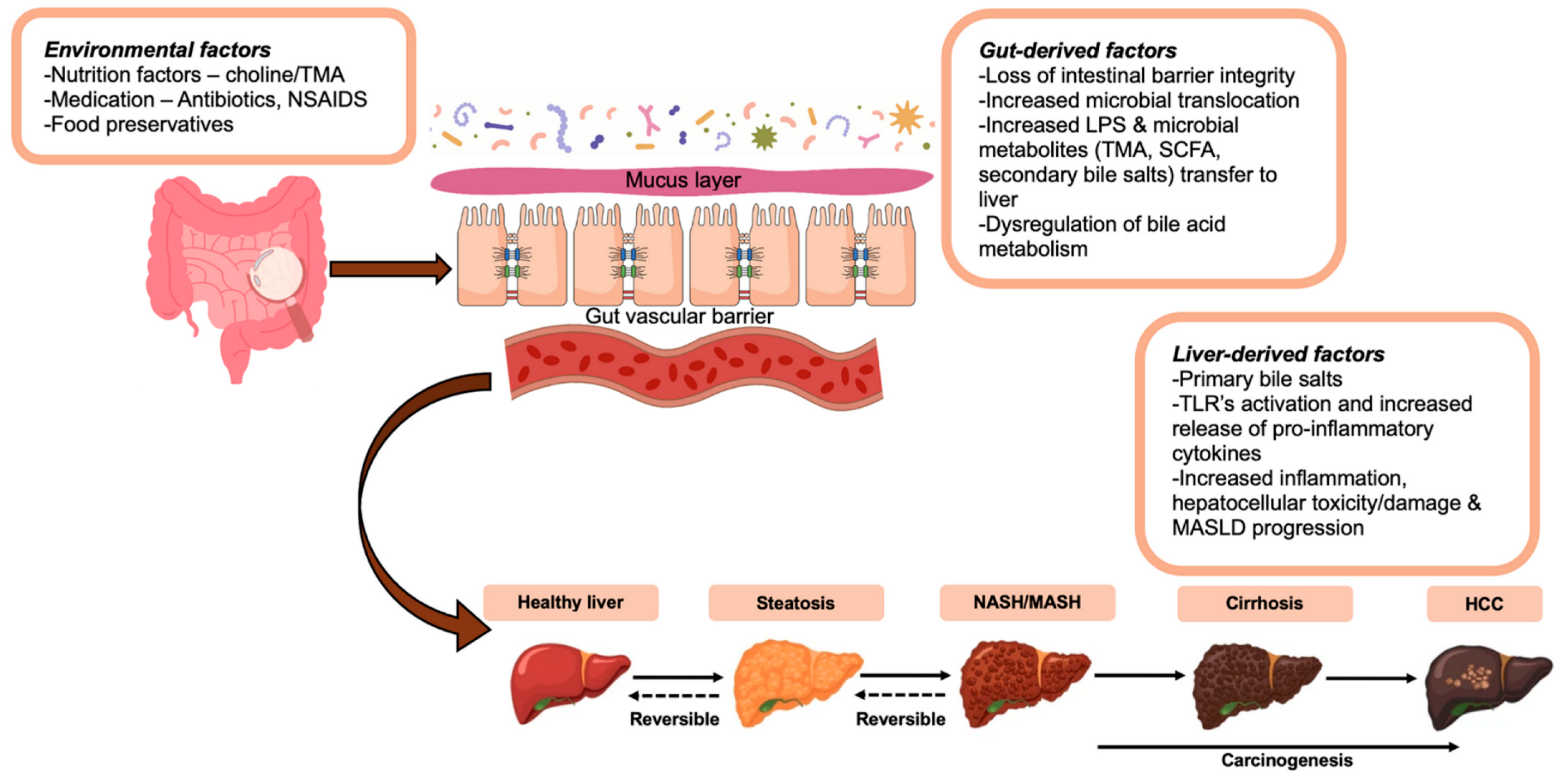
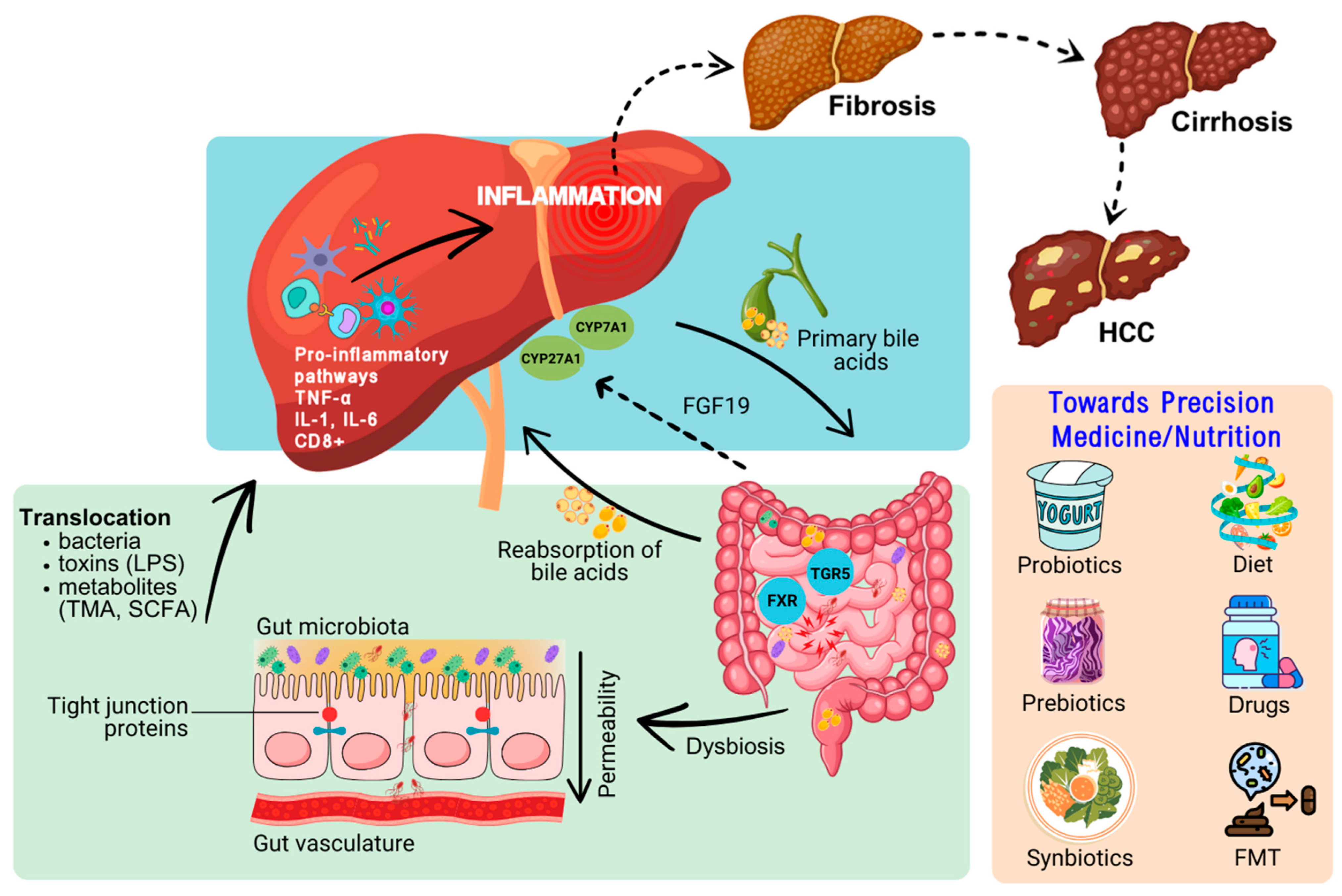

{kind=link}
{kind=link}
| Drug [Ref.] | Selected Key Clinical Evidence to Date | Drug Target and Mechanism of Action | |
|---|---|---|---|
| Cohort | Key Results | ||
| Nidufexor [98] | NASH patients | Improved NAFLD activity score and lower triglyceride levels | FXR agonists. FXR is a nuclear receptor primarily expressed in the liver. FXR activation regulates bile acid synthesis and improves lipid metabolism. This modulation of bile acids and cholesterol homeostasis has been shown to reduce hepatic inflammation and injury |
| PX-104 [99] | Non-diabetic NAFLD patients | Improved insulin sensitivity and liver enzymes (ALT and GGT) levels | |
| Cilofexor [100] | NASH patients | Reduction in elevated liver enzyme (GGT) levels, hepatic steatosis and serum bile acids | |
| Tropifexor [101] | NASH patients | Reduction in elevated liver enzyme (ALT and AST) levels, and hepatic steatosis | |
| Obeticholic acid [102,103] | NASH and NASH-related fibrosis patients | Improved NASH activity scores and fibrosis severity | |
| Ursodeoxycholic acid [104] | NASH patients | Reduction in elevated liver enzyme (ALT and GGT) levels | FXR antagonist. FXR antagonists indirectly modulate bile acid synthesis, and thereby reduce bile acid-mediated hepatic stress. They also reduce ceramide levels, which have been implicated in lipid-induced hepatocyte injury |
| Semaglutide [105] | NASH patients | Significant NASH resolution in patients on semaglutide compared to placebo | GLP-1 agonist. GLP-1 agonists act via GLP-1 receptors to enhance insulin secretion, delay gastric emptying, and reduce appetite. Reduces hepatic steatosis by improving hepatic insulin sensitivity and reducing systemic inflammation and lipotoxicity |
| Aldafermin [106] | NASH and NASH-related fibrosis patients | Dose-dependent reductions in serum bile acids and improvements in fibrogenesis markers | FGF19 analogue. FGF analogues reduce bile acid synthesis and improve lipid metabolism. This reduces hepatic bile acid toxicity and reduces systemic inflammation and lipotoxicity |
| Efruxifermin [107] | NASH-related fibrosis | Improvements in fibrosis stage and fibrogenesis markers | FGF21 analogue |
| Saroglitazar [108] | NAFLD patients | Reduction in elevated liver enzyme (ALT) levels and improved hepatic stiffness and metabolic parameters | PPAR agonist. PPAR agonists enhance fatty acid oxidation and reduce lipogenesis. They also suppress pro-inflammatory cytokine production, improving insulin sensitivity and reducing oxidative stress in the liver |
| Resmetirom [109] | NASH-related fibrosis | Significant NASH resolution and improvement in fibrosis stage compared to placebo | THR-β agonist. THR-β agonists activate the beta thyroid hormone receptors predominantly expressed in the liver. This enhances hepatic fatty acid oxidation, reduces lipogenesis, and lowers circulating triglycerides, resulting in reduced hepatic steatosis and improved glucose and lipid metabolic profiles |
| Study [Ref.] | Design and Duration | Study Intervention | Study Groups | Key Study Outcomes | Key Study Limitations |
|---|---|---|---|---|---|
| Derosa et al., 2022 [116] | Double-blind, placebo-controlled, RCT; 12 weeks | Probiotic group: 2 sachets daily in the morning of VSL#3 [a high-concentration multi-strain probiotic mix containing eight different strains: one strain of Streptococcus thermophilus BT01, three strains of Bifidobacteria (B. breve BB02; B. animalis subspecies [subsp.] lactis BL03, previously identified as B. longum BL03; and B. animalis subsp. lactis BI04, previously identified as B. infantis BI04), and four strains of Lactobacilli (L. acidophilus BA05, L. plantarum BP06, L. paracasei BP07, and L. helveticus BD08, previously identified as L. delbrueckii subsp. bulgaricus BD08)] Control group: placebo | Probiotic (VSL#3) group: 30 patients with NAFLD (mean age ± standard deviation: 55.7.0 ± 6.7 and 55.9 ± 7.5 years for males and females, respectively) Placebo control group: 30 patients with NAFLD (mean age: 55.8 ± 7.7 and 57.5 ± 7.9 years for males and females, respectively) | The VSL#3 group had significantly improved hepatic parameters, echography grading, inflammatory biomarkers and triglyceride levels, compared to the placebo | Small sample size. Short duration. |
| Chong et al., 2021 [119] | Double-blind, placebo-controlled, proof-of-concept RCT; 10 weeks | Probiotic group: VSL#3 Control group: placebo | VSL#3 group: 19 patients with NAFLD (mean age: 57.0 ± 8.0 years) Placebo control group: 16 patients with NAFLD (mean age: 58.0 ± 7.0 years) | No significant differences in biomarkers of cardiovascular risk and liver injury between the VSL#3 and placebo groups. | Small sample size. Short duration. |
| Duseja et al., 2019 [117] | Double-blind, placebo-controlled, RCT; 12 months | Probiotic group: 2 capsules three times daily (each capsule containing 112.5 billion live, lyophilised, lactic acid bacteria and bifidobacteria: Lactobacillus paracasei DSM 24733, L. plantarum DSM 24730, L. acidophilus DSM 24735, L. delbrueckii subsp. bulgaricus DSM 24734, Bifidobacterium longum DSM 24736, B. infantis DSM 24737, B. breve DSM 24732, Streptococcus thermophilus DSM 24731) Control group: placebo | Probiotic group: 19 patients with NASH (mean age: 38.0 ± 10.0 years) Placebo control group: 20 patients with NASH (mean age: 33.0 ± 6.0 years) | A significant improvement in liver histology, ALT, and cytokine profiles in the probiotic group compared to the placebo group. | Small initial sample size, with a further reduction in patients at follow-up for a repeat liver biopsy. At baseline, most patients had mild hepatic histology. Histopathology interpretation by a single pathologist at respective study centers. |
| Mohamad Nor et al., 2021 [118] | Double-blind, placebo-controlled, RCT; 6 months | Probiotic group: One 3 g sachet twice daily of HEXBIO® microbial cell preparation (B-Crobes Laboratory Sdn. Bhd; each sachet consists of 30 billion CFU with six probiotic strains: Lactobacillus acidophilus BCMC® 12,130 (107 mg), L. casei subsp. BCMC® 12,313 (107 mg), L. lactis BCMC® 12,451 (107 mg), Bifidobacterium bifidum BCMC® 02290 (107 mg), B. infantis BCMC® 02129 (107 mg) and B. longum BCMC® 02120 (107 mg)] Control group: placebo | Probiotic group: 17 patients with NAFLD (mean age: 54.70 ± 10.19 years) Placebo control group: 22 patients with NAFLD (mean age: 52.47 ± 16.73 years) | Probiotics did not significantly improve clinical outcomes in NAFLD patients compared to placebo, but may help to stabilize the mucosal immune function and protect against increased intestinal permeability. | Small sample size. The unanticipated reduction in total fat intake in the probiotics group may have influenced the study outcomes. |
| Ayob et al., 2023 [133] | Double-blind, placebo-controlled, RCT; 6 months | Probiotic group: one 3 g sachet twice daily of HEXBIO® microbial cell preparation (B-Crobes Laboratory Sdn. Bhd) Control group: placebo | Probiotic group: 18 patients with NAFLD (mean age: 55.00 ± 11.07 years) Placebo control group: 22 patients with NAFLD (mean age: 49.95 ± 14.05 years) | Compared to placebo, probiotic supplementation ameliorated dysbiosis in patients with NAFLD, leading to stabilization of pro-inflammatory cytokine expression and modulation of mucosal immune function. | Small sample size. Due to the COVID-19 pandemic, some study patients experienced interruptions in receiving probiotics. |
| Wong et al., 2013 [134] | Open-label RCT; 6 months | Probiotic group: one 10 g sachet of Lepicol probiotic (200 million probiotic cultures) formula (Healthy Bowels Company Ltd., UK) twice daily. The Lepicol probiotic formula contains: Lactobacillus plantarum, L. deslbrueckii, L. acidophilus, L. rhamnosus, and Bifidobacterium bifidum, as well as 3 g of fructo-oligo-saccharides (prebiotics). Control group: usual care | Probiotic group: 10 patients with biopsy-proven NASH (mean age: 42.0 ± 9.0 years) Control group: 10 patients with biopsy-proven NASH (mean age: 55.0 ± 9.0 years) | Probiotic treatment may be more effective than usual care in reducing liver fat and AST levels in patients with NASH. | Small sample size. Open-label RCT. Results cannot be extrapolated directly to other probiotic formulae [the Lepicol probiotic formula contains also 3 g of fructo-oligo-saccharides (prebiotics)]. |
| Sepideh et al., 2016 [135] | Double-blind, placebo-controlled, RCT; 8 weeks | Probiotic group: 1 g daily of Lactocare (Zist-takhmir Co., Tehran, Iran) which contains 7 strains of naturally occurring bacteria (Lactobacillus casei, L. acidophilus, L. rhamnosus, L. bulgaricus, Bifidobacterium breve, Bifidobacterium longum, Streptococcus thermophiles) Control group: placebo | Probiotic group: 11 patients with NAFLD (mean age: 42.10 ± 1.99 years) Placebo control group: 11 patients with NAFLD (mean age: 47.33 ± 2.53 years) | The probiotic group exhibited a reduction in insulin resistance, glycemic and inflammatory markers compared to the placebo group. | Small sample size. Short duration. Significant dropout rate. |
| Aller et al., 2011 [136] | Double-blind, placebo-controlled, RCT; 12 weeks | Probiotic group: one tablet daily (500 million of Lactobacillus bulgaricus and Streptococcus thermophiles; Nutricion Medica, SL, Spain) Control group: placebo (one placebo tablet daily with 120 mg of starch) | Probiotic group: 14 patients with biopsy-diagnosed NAFLD (mean age: 49.4 ± 10.9 years) Placebo control group: 14 patients with biopsy-diagnosed NAFLD (mean age: 44.3 ± 15.1 years) | Significantly reduced liver enzymes in the probiotic group compared to the placebo group. | Small sample size. Short duration. |
| Ahn et al., 2019 [137] | Double-blind, placebo-controlled, RCT; 12 weeks | Probiotic group: mixture (109 CFU/1.4 g) of six probiotics [L. acidophilus CBT LA1, L. rhamnosus CBT LR5 isolated from Korean human feces, L. paracasei CBT LPC5 isolated from Korean fermented food (jeotgal), P. pentosaceus CBT SL4 isolated from a Korean fermented vegetable product (kimchi), B. lactis CBT BL3, and B. breve CBT BR3 isolated from Korean infant feces] Control group: placebo | Probiotic group: 30 patients with NAFLD (mean age: 41.7 ± 12.49 years) Placebo control group: 35 patients with NAFLD (mean age: 44.71 ± 13.31 years) | Significantly reduced levels of triglycerides and intrahepatic fat in the probiotic group compared to the placebo group. | Small sample size. Short duration. The controlled attenuation parameter was measured with the M FibroScan probe, while patients with BMI ≥ 28 kg/m2 should be assessed with an XL probe to increase the reliability of steatosis measurements and to reduce scan failures. Eight study patients were excluded due to unreliable measurement results. |
| Escouto et al., 2023 [138] | Double-blind, placebo-controlled, RCT; 24 weeks | Probiotic group: one capsule daily (1 × 109 CFU Lactobacillus acidophilus ATCC SD5221 and 1 × 109 CFU Bifidobacterium lactis HN019; Beneflora Caps; Biolab Sanus Farmaceutica Ltd.a.) Control group: placebo | Probiotic group: 23 patients with NASH (median age: 58.0 years) Placebo control group: 25 patients with NASH (median age: 57.0 years) | No statistically significant differences for liver fibrosis, steatosis, and inflammatory activity between the RCT groups. Significantly improved APRI score with probiotic supplementation compared to placebo. | Small sample size. Short duration. Inclusion of patients with liver cirrhosis. Most study participants were on statins and/or metformin treatment, which may have influenced relevant metabolic markers. |
| Bomhof et al., 2019 [121] | Single-blinded (participants blinded to treatment), placebo-controlled, RCT; 36 weeks | Prebiotic group: oligofructose (Orafti P95, Beneo-Orafti Inc., Tienen, Belgium) 8 g once daily for 12 weeks followed by 16 g daily for 24 weeks Control group: placebo | Prebiotic group: 8 patients with biopsy-confirmed NASH (mean age: 45.3 ± 5.6 years) Placebo control group: 6 patients with biopsy-confirmed NASH (mean age: 53.3 ± 4.8 years) | Significantly improved NAS score and steatosis in the probiotic group compared to the placebo group. | Small sample size. Single-blind RCT. The placebo group had poorer glycemic control compared to the prebiotic group. Following the NASH diagnosis, several study participants made lifestyle changes resulting in significant weight loss prior to randomization. |
| Reshef et al., 2024 [139] | Double-blind, placebo-controlled, RCT; 12 weeks | Prebiotic group: 8 g twice daily of inulin-type fructans (ITF) supplementation with inulin/oligofructose (75/25) Control group: placebo | Prebiotic group: 8 patients with NASH (median age: 47.8 years) Placebo control group: 11 patients with NASH (median age: 50.0 years) | The prebiotic group had a significant increase in the relative abundance of fecal Bifidobacterium; however, no significant improvement was observed in liver fat content or inflammatory biomarkers. | Small sample size. Short duration. Potential confounders, such as the use of metformin. |
| Scorletti et al., 2020 [140] | Double-blind, placebo-controlled, RCT; ≈12 months (10 months minimum to 14 months maximum) | Synbiotic group: synbiotic consisting of fructo-oligosaccharides with a polymerization degree < 10 (4 g twice daily; two sachets daily) plus Bifidobacterium animalis subsp. lactis BB-12 (10 billion CFU daily minimum; one capsule daily). Actilight®950P was the prebiotic component of the synbiotic Control group: placebo (4 g twice daily of maltodextrin; one capsule daily, plus two sachets daily) | Synbiotic group: 45 patients with NAFLD (mean age: 50.2 ± 12.4 years) Placebo control group: 44 patients with NAFLD (mean age: 51.6 ± 13.1 years) | Compared to placebo, the synbiotic supplementation induced changes in the fecal microbiome (growth of Faecalibacterium and Bifidobacterium at the expense of Oscillibacter and Alistipes). The gut microbiota composition changes were not associated with any primary outcome. No significant difference in hepatic fat reduction between the two RCT groups. | Small sample size. Selection of one strain of bacteria for the synbiotic. |
| Mofidi et al., 2017 [141] | Double-blind, placebo-controlled, RCT; 28 weeks | Synbiotic group: synbiotic supplementation capsule twice daily [Protexin; Probiotics International Ltd.; each capsule contained: 200 million bacteria (seven strains: Lactobacillus casei, L. rhamnosus, L. acidophilus, L. bulgaricus, Streptococcus thermophilus, Bifidobacterium breve, and B. longum); prebiotic (125 mg fructo-oligosaccharide; probiotic cultures (magnesium stearate); and hydroxyl-propylmethyl cellulose] Control group: placebo | Synbiotic group: 21 patients with NAFLD (mean age: 40.09 ± 11.44 years) Placebo control group: 21 patients with NAFLD (mean age: 44.61 ± 10.12 years) | Both RCT groups exhibited reductions in hepatic steatosis and fibrosis with the mean reduction being significantly greater in the synbiotic group compared to the placebo. | Small sample size. Only participants with BMI ≤ 25 kg/m2 were included in this RCT, thus these findings cannot be directly extrapolated for patients with NAFLD and overweightness or obesity. |
| Malaguarnera et al., 2012 [142] | Double-blind, placebo-controlled, RCT; 24 weeks | Synbiotic group: sachet of Bifidobacterium longum and fructo-oligosaccharides (Fos) (Zirfos, Alfa Wassermann, Bologna, Italy) 2.5 g, vitamin B1 (1.4 mg), vitamin B2 (1.6 mg), vitamin B6 (2.0 mg), and vitamin B12 (1.0 mg) plus lifestyle modification (diet and exercise) Control group: placebo plus lifestyle modification (diet and exercise) | Synbiotic group: 34 patients with NASH (mean age: 46.9 ± 5.4 years) Placebo control group: 32 patients with NASH (mean age: 46.7 ± 5.7 years) | Significantly reduced NASH activity index and steatosis, as well as serum endotoxin, AST, CRP, TNF-α levels and HOMA-IR, in the synbiotic group compared to the placebo group. | Small sample size. Short duration. Potential diet-related confounding. |
| Asgharian et al., 2016 [143] | Double-blind, placebo-controlled, RCT; 8 weeks | Synbiotic group: 500 mg capsule (Familact by Zisttakhmir) once daily containing seven probiotic bacteria species (Lactobacillus casei, L. acidophilus, L. rhamnosus, L. bulgaricus, Bifidobacterium breve, B. longum, and Streptococcus thermophilus) and fructo-oligosaccharides Control group: placebo | Synbiotic group: 38 patients with NAFLD (mean age: 46.57 ± 1.7 years) Placebo control group: 36 patients with NAFLD (mean age: 47.78 ± 1.7 years) | The synbiotic group demonstrated significantly reduced steatosis grade on ultrasound compared to baseline. No association of synbiotic supplementation with changes in ALT, AST and CRP levels. | Small sample size. Short duration. |
| Abhari et al., 2020 [144] | Double-blind, placebo-controlled, RCT; 12 weeks | Synbiotic group: one synbiotic capsule daily containing 109 spore of Bacillus coagulans (GBI-30) plus 0.4 g inulin Control group: placebo | Synbiotic group: 23 patients with NAFLD (mean age: 47.7 ± 11.4 years) Placebo control group: 22 patients with NAFLD (mean age: 46.7 ± 12.4 years) | Hepatic steatosis measured by the controlled attenuation parameter score in Fibroscan, serum ALT and GGT levels decreased significantly more in the synbiotic group compared to the placebo. Synbiotic supplementation significantly reduced serum TNF-α levels and nuclear factor-kB activity. | Small sample size. Short duration. |
| Sayari et al., 2018 [145] | Double-blind, placebo-controlled, RCT; 16 weeks | Sitagliptin plus Synbiotic group: Sitagliptin (50 mg once daily) plus synbiotic [500 mg once daily; each capsule (Familakt) contained 109 CFU of Lactobacillus casei, L. rhamnosus, L. acidophilus, L. bulgaricus, Bifidobacterium breve, B. longum, Streptococcus thermophiles; prebiotic (fructo-oligosaccharide); probiotic cultures (magnesium stearate); and hydroxyl-propyl methylcellulose] Sitagliptin plus placebo group: Sitagliptin (50 mg once daily) plus placebo (maltodexterin; once daily) | Sitagliptin plus Synbiotic group: 70 patients with NAFLD and BMI from 25 to 29.9 kg/m2 who had impaired fasting blood glucose and/or impaired glucose tolerance (mean age: 42.48 ± 11.41 years) Sitagliptin plus Placebo group: 70 patients with NAFLD and BMI from 25 to 29.9 kg/m2 and who had impaired fasting blood glucose and/or impaired oral glucose tolerance test (mean age: 43.42 ± 11.65 years) | The Sitagliptin [a dipeptidyl peptidase-4 (DPP-4) inhibitor] plus Synbiotic group had significantly greater improvements in AST, fasting glucose, cholesterol and LDL levels compared to the Sitagliptin plus Placebo group. | Small sample size. Short duration. |
| Xue et al., 2022 [129] | RCT; 4 weeks (RCT participants returned for re-examination 1 month post-treatment) | FMT group: allogenic FMT (heterologous from healthy donors via colonoscopy followed by three enemas over 3 days) Non-FMT control group: oral probiotics (a Bifidobacterium viable preparation and Lactobacillus acidophilus capsules) | FMT group: 47 patients with NAFLD (mean age: 57.3 ± 13.4 years) Non-FMT group: 28 patients with NAFLD (mean age: 60.2 ± 8.5 years) Healthy control group: 10 healthy undergraduate students (control group that received no intervention) | The mean hepatic fat attenuation values decreased from 278.3 to 263.9 dB/m in the FMT group (p = 0.049), while these increased from 265.5 to 282.5 dB/m in the Non-FMT group (p < 0.01). Compared to healthy controls, patients with NAFLD before the FMT had lower Chaol indices (p < 0.05), suggesting lower abundances of gut microbiota. Post-FMT, there was no statistical difference in the Chaol indices between the FMT NALFD patients and healthy controls. In the FMT group, better effects on the gut microbiota reconstruction were noted in lean patients with NAFLD, compared to those with obesity. | Small sample size. Short duration. Patients with NAFLD were randomly divided into the FMT and non-FMT RCT group based on the wishes of the participants. Age difference between the FMT/non-FMT groups and the healthy control group. Potential confounders relating to exercise/diet. |
| Witjes et al., 2020 [146] | Double-blind (all study participants, i.e., vegan donors and patients with NAFLD and all trial physicians/authors were blinded for the treatment until trial completion), proof-of-principle RCT; 24 weeks (FMT performed three times at 8-week intervals) | Allogenic FMT group: FMT with feces from healthy, lean, vegan donors three times at 8-week intervals (same donor for the three FMTs in each participant) Autologous FMT group: 8-week intervals Allogenic/Autologous FMT via gastroduodenoscopy at baseline, and via a duodenal tube via a CORTRAK enteral access (Medline Industries, Northfield, IL), at 8 weeks and 16 weeks | Allogenic FMT group: 10 omnivorous adults with treatment-naïve biopsy-proven NAFLD (hepatic steatosis on ultrasound) and obesity (mean age: 51.2 ± 6.6 years) Autologous FMT group: 11 omnivorous adults with treatment-naïve biopsy-proven NAFLD (hepatic steatosis on ultrasound) and obesity (mean age: 48.5 ± 10.2 years) | No statistically significant changes in the overall NAFLD activity score, steatosis grade and fibrosis scores. In the allogenic FMT group a trend for an improved necro-inflammation score was noted. Significant changes in the expression of hepatic genes involved in inflammation and lipid metabolism were also noted in the allogenic FMT group, compared with the autologous FMT. Intestinal microbial community structure changed following allogenic FMT, and was associated with changes in plasma metabolites and markers of steatohepatitis. | Underpowered study due to slow recruitment. Differences in baseline AST and ALT levels between the allogenic and autologous FMT groups. Study participants were relatively healthy, and, thus, the study sample was not representative of the entire spectrum of patients with NAFLD. |
| Craven et al., 2020 [147] | Double-blind, RCT (3:1 ratio to allogenic or autologous FMT); 24 weeks (follow-up appointments for 6 months post-FMT) | Allogenic FMT group: FMT with feces from healthy lean donors Autologous FMT group Allogenic/autologous FMT was delivered to the duodenum via endoscopy | Allogenic FMT group: 15 patients with NAFLD (mean age: 47.6 ± 14.9 years) Autologous FMT group: 6 patients with NAFLD (mean age: 57.5 ± 13.0 years) | No significant changes in hepatic proton density fat fraction or HOMA-IR in the allogenic or autologous FMT group. NAFLD patients (n = 7) with elevated small intestinal permeability at baseline had a significant reduction 6 weeks post allogenic FMT. | Small sample size. The two FMT groups differed regarding biochemical data, intestinal permeability. and fibrosis staging. |
| Abdel-Razik et al., 2018 [148] | Double-blind, placebo-controlled, RCT; 6 months | Rifaximin group: rifaximin 1100 mg daily (550 mg tablets 1 × 2 before meals) Control group: placebo | Rifaximin group: 25 patients with biopsy-proven NASH (mean age: 40.2 ± 9.88 years) Placebo control group: 25 patients with biopsy-proven NASH (mean age: 38.4 ± 9.21 years) | Significantly reduced NAFLD-liver fat score, HOMA-IR and toll-like receptor-4 expression, as well as ALT, AST, GGT, TNF-α, IL-6, CK-18 and endotoxin levels, were noted in the Rifaximin group, but not in the placebo group. | Small sample size. A second liver biopsy at 6 months post-treatment was not performed for any participants. The effect of rifaximin treatment on the gut flora was not assessed. |
| Neuschwander-Tetri et al., 2015 [149] | Double-blind, placebo-controlled, RCT; 72 weeks | Obeticholic acid group: 25 mg of 6-ethylchenodeoxycholic acid (obeticholic acid) once daily, per os (orally) Control group: placebo | Obeticholic acid group: 141 patients with biopsy-proven non-cirrhotic NASH (mean age: 52.0 ± 11.0 years) Placebo control group: 142 patients with biopsy-proven non-cirrhotic NASH (mean age: 51.0 ± 12.0 years) | More patients in the obeticholic acid group compared with placebo had improvement in the NAFLD activity score, fibrosis, hepatocellular ballooning, steatosis, and lobular inflammation. The mean change in the NAFLD activity score was also greater in the obeticholic acid group compared with placebo. The proportion of patients with resolution of NASH (change from baseline NASH diagnosis to non-NASH) did not differ between the two RCT groups. Compared with placebo, obeticholic acid treatment was associated more with pruritus and a temporary increase in total and LDL cholesterol with a modest decrease in HDL cholesterol. | Missing detailed tracking of interventions for the treatment of hyperlipidemia during the RCT, including dosing information. Smoking history was not captured until midway of the RCT. |
| Harrison et al., 2018 [150] | Double-blind, placebo-controlled, RCT; 12 weeks | NGM282 groups: (a) 3 mg NGM282 (an engineered FGF19 analogue) subcutaneously once daily (b) 6 mg NGM282 subcutaneously once daily Control group: Placebo | 3 mg NGM282 group: 27 patients with biopsy-proven NASH (mean age: 52.0 ± 7.1 years) 6 mg NGM282 group: 28 patients with biopsy-proven NASH (mean age: 56.4 ± 7.8 years) Placebo control group: 27 patients with biopsy-proven NASH (mean age: 52.8 ± 11.3 years) | Between baseline and 12 weeks, at least a 5% reduction in absolute hepatic fat content was noted for 74% and 79% of the participants in the 3 mg and 6 mg dose group, respectively (only for 7% in the placebo group). Adverse events were more frequent in the NGM282 groups compared to placebo, with injection site reactions, diarrhea, abdominal pain and nausea being the most commonly (≥10%) reported adverse events. | Small sample size. Short duration. Inclusion of few non-white participants and numerically more men (non-significantly) in the NGM282 groups, although, in post hoc analysis, sex and race had no effect on reductions in hepatic fat content. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelhameed, F.; Mustafa, A.; Kite, C.; Lagojda, L.; Dallaway, A.; Than, N.N.; Kassi, E.; Kyrou, I.; Randeva, H.S. Gut Microbiota and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD): Emerging Pathogenic Mechanisms and Therapeutic Implications. Livers 2025, 5, 11. https://doi.org/10.3390/livers5010011
Abdelhameed F, Mustafa A, Kite C, Lagojda L, Dallaway A, Than NN, Kassi E, Kyrou I, Randeva HS. Gut Microbiota and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD): Emerging Pathogenic Mechanisms and Therapeutic Implications. Livers. 2025; 5(1):11. https://doi.org/10.3390/livers5010011
Chicago/Turabian StyleAbdelhameed, Farah, Attia Mustafa, Chris Kite, Lukasz Lagojda, Alexander Dallaway, Nwe Ni Than, Eva Kassi, Ioannis Kyrou, and Harpal S. Randeva. 2025. "Gut Microbiota and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD): Emerging Pathogenic Mechanisms and Therapeutic Implications" Livers 5, no. 1: 11. https://doi.org/10.3390/livers5010011
APA StyleAbdelhameed, F., Mustafa, A., Kite, C., Lagojda, L., Dallaway, A., Than, N. N., Kassi, E., Kyrou, I., & Randeva, H. S. (2025). Gut Microbiota and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD): Emerging Pathogenic Mechanisms and Therapeutic Implications. Livers, 5(1), 11. https://doi.org/10.3390/livers5010011