1. Introduction
Hepatocellular carcinoma (HCC) represents a major health challenge worldwide as most patients suffer advanced disease conditions and carry poor prognoses [
1]. Src family kinase (SFK) is considered a potential target in HCC therapy as cellular Rous sarcoma oncogene cellular homolog (c-Src) is a critical component in various signaling pathways that regulate metastasis, invasion, proliferation, and angiogenesis [
1]. Further, c-Src interacts with other receptor kinases for downstream signaling by PI3K/PTEN/Akt, Ras/Raf/Mek, and STATs [
2]. Metastasis is also a primary cause of cancer-related death and can be treated by targeting the oncoprotein Src, which is found to be elevated in various cancer cells. Thus, it may possible to reduce the metastasis in cancer cells by inhibiting the expression of the Src protein [
3,
4]. Das, a Food and Drug Administration (FDA) approved drug, is currently in a phase II trial for the treatment of various cancers. It can be used for targeted therapy as it strongly binds to the SFK members and also inhibits BCR/ABL fusion protein [
5]. Several studies reported the anti-tumor activity of Das in combination with other chemotherapeutic drugs in the treatment of tumors [
6,
7]. In vitro studies reported that Das treatment can successfully suppress the migration, proliferation, and invasion in HCC cells by inhibiting the Src tyrosine kinase via affecting PI3K/PTEN/Akt pathways [
8]. In another report, Sayed et al. reported the synergistic anti-tumor effect of Das in combination with rosuvastatin in an in vitro HCC model and showed the involvement of the signaling pathway HCG/FAK/Src and its downstream pathways such as Ras/Raf/ERK and PI3K/Akt and molecules such as matrix metalloprotease 9 (MMP-9) and caspase-3 [
1]. Combined treatment of Das and temozolomide also causes synergistic effects and induces autophagic cell death and disruption to the cell cycle in glioma cells. In another study, Das in combination with cabozantinib was significantly effective against alveolar soft part sarcoma (ASPS) cells [
6].
Oncogenesis involves several metabolic changes that occur to fulfill the specific metabolic requirement of cancerous cells. These changes may enhance the rate of glucose transport associated with a high glycolysis rate, decrease pyruvate oxidation with increased lactate production, reduce fatty acid oxidation rate, and decrease the activity of the glycerol-3-phosphate shuttle system [
9,
10]. Pcar, C16:00, is synthesized from palmitic acid (PA) and represents ~80% of the total fatty acids present in cells. Various studies have reported the Pcar and carnitine levels in the tissues of humans and animals such as fish and rats [
11,
12,
13]. However, it is not clear how targeting mitochondrial oxidative phosphorylation can decrease the survival of cancerous cells. Several research groups demonstrated that the fatty acid-derived mitochondrial substrate Pcar proved toxic to prostate [
11] and colorectal cancerous cells [
14,
15]. Cancer cell metabolism is reported to have limited oxidative phosphorylation to minimize oxidative stress. Various available studies demonstrated that activation of oxidative phosphorylation promotes the reactive oxygen species (ROS) generation in cells. For example, activation of pyruvate dehydrogenase can redirect pyruvate towards oxidative phosphorylation which generates ROS and reduces proliferation in lung and tongue cancer cells [
16]. The supply of Pcar to cancerous cells in combination with carnitine increases mitochondrial respiration and induces apoptosis. Wenzel et al. also reported that the production of superoxide during Pcar oxidation is lethal to HT29 cancer cells but tolerated by preneoplastic NCOL-1 epithelial cells [
14]. Thus, this approach proves that Pcar oxidation may be less toxic to healthy cells as Pcar is a natural metabolite oxidized by mitochondria. The heterogeneous effect of Pcar on cell survival can be linked to inherent differences in glutathione redox buffering response [
15]. Pcar has also been reported to alter the activity of several enzymes and transporters located in a mitochondrial membrane that triggers the caspase activity in cells [
17].
Based on the aforementioned data, the present study aims to demonstrate the synergistic anti-tumor potential of Das and Pcar in an in vitro and in vivo HCC model by inducing HepG2-mediated liver cancer in nude mice. Several signaling pathways related to cancer cell survival, metastasis, and proliferation are also evaluated to elucidate the mechanism behind the anti-tumor activity of Das and Pcar. Results suggest that Das in combination with Pcar acts synergistically and inhibits the proliferation as well as migration of cancerous cells. The in vivo study also supports the observation that it suppresses tumor growth in an induced HCC mouse model.
2. Material and Method
2.1. Synthesis of Palmitoylcarnitine Chloride
First, 6 g of thionyl chloride and 50 g of palmitic acid were mixed and stirred for 3 h at 80 °C, followed by the addition of a solution containing 9.9 g of dl-carnitine chloride in 50 g of trichloroacetic acid at 40 °C. The final solution was stirred at 80 °C for 3 h and then poured into 200 mL of dry ether. The precipitate obtained was filtered, washed with ether, and dissolved in 50 mL of hot isopropyl alcohol. Turbid isopropyl alcohol solution (due to dl-carnitine chloride) was filtered, followed by the addition of 400 mL dry acetone to the filtrate. Pcar chloride was precipitated, filtered, washed with ether, and dried [
18]. The purity of synthesized Pcar was found to be approximately 99%. NMR spectra were obtained to confirm the synthesis of Pcar (
Figure S1).
The 1H NMR spectra were recorded with a Varian Mercury Plus 400 spectrometer operating at 400.1 MHz. The spectra were acquired at room temperature (298 K). The chemical shifts are reported in ppm for reference and are stated relative to external tetramethylsilane (TMS) for 1H NMR.
1H NMR [DMSO-d6, ppm]: δ 5.50, 5.48, 5.47, 5.45 (q, J = 6.4 Hz, 1H, 1O-CH), 3.88, 3.84 (dd, J = 14.4, 8.3 Hz, 1H, 1N-CH), 3.74, 3.71 (d, J = 13.7 Hz, 1H, 1N-CH), 3.16 (s, 9H, 3N-CH3), 2.70, 2.68 (d, J = 6.0 Hz, 2H, O=C-CH2), 2.34, 2.32, 2.30 (td, J = 7.5, 2.7 Hz, 2H, O=C-CH2), 1.57, 1.55, 1.53 (t, J = 6.8 Hz, 2H, CH2), 1.25 (m, 24H, CH2), 0.88, 0.87, 0.85 (t, J = 6.8 Hz, 3H, CH3).
2.2. Cell Culture
Liver cancer cell line HepG2 cells and non-cancerous cell line HEK293 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 1% antibiotic and 10% FBS. Cells were maintained in a humified atmosphere with 5% CO2 at 37 °C.
2.3. Cell Viability Assay
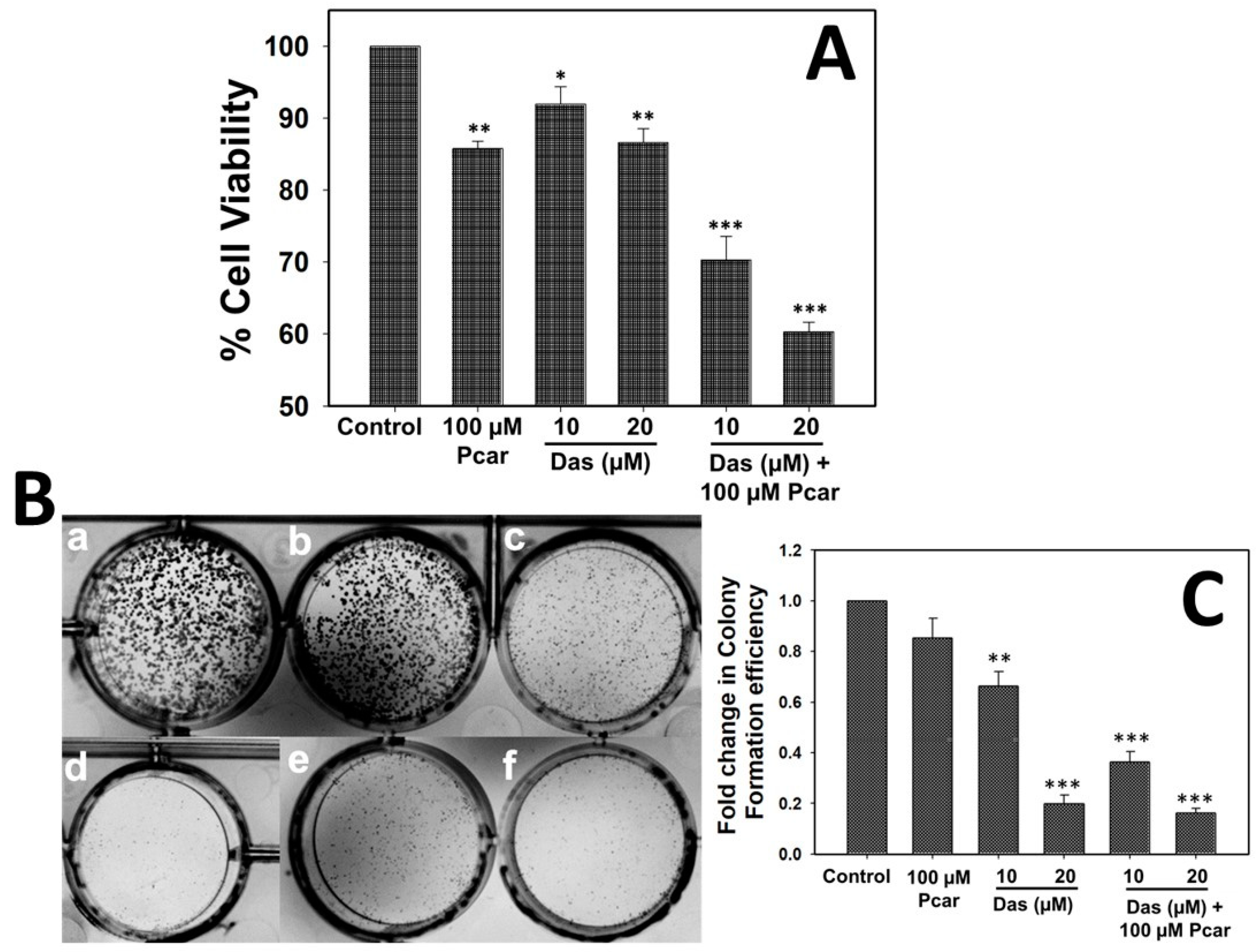
Cell viability was determined by mitochondrial 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide) (MTT) assay. In this assay, 1 × 104 cells were seeded in a 96-well plate and treated with different concentrations of Das (10 and 20 µM in DMEM media) and Pcar (100 µM in DMEM media) alone and in combination for 24 h. Following the completion of the exposure period, the media were withdrawn, and each well received 100 µL of MTT (0.5 mg/mL in media), followed by incubation for 3–4 h at 37 °C. Dimethyl sulfoxide (DMSO) was added after the MTT dye was discarded in order to solubilize the blue formazan. Color intensity was quantified in an iMark Bio-rad microplate reader by recording the absorbance at 590 nm.
2.4. Combination Index Analysis
In combination therapy, the interaction between two drugs is evaluated by combinational index (CI) analysis. It provides a quantitative measure of drug interaction [
19].
CA,X and CB,X are the concentrations of drugs A and B used to achieve X% effect in combination therapy. ICX,A and ICX,B are the concentrations of a single drug used to achieve the same effect. In general, CI < 0.9, CI = 0.9–1.1, and CI > 1.1 indicate synergism, additivity, and antagonism, respectively.
2.5. Clonogenic Assay
In order to investigate the colony-forming efficiency of cells, a clonogenic assay was performed. One thousand cells were seeded in a 6-well plate, followed by the addition of Das (10 and 20 µM) and Pcar (100 µM) alone and in combination. In order to form the colonies, cells were further incubated for 10 more days.
2.6. Migration Assay
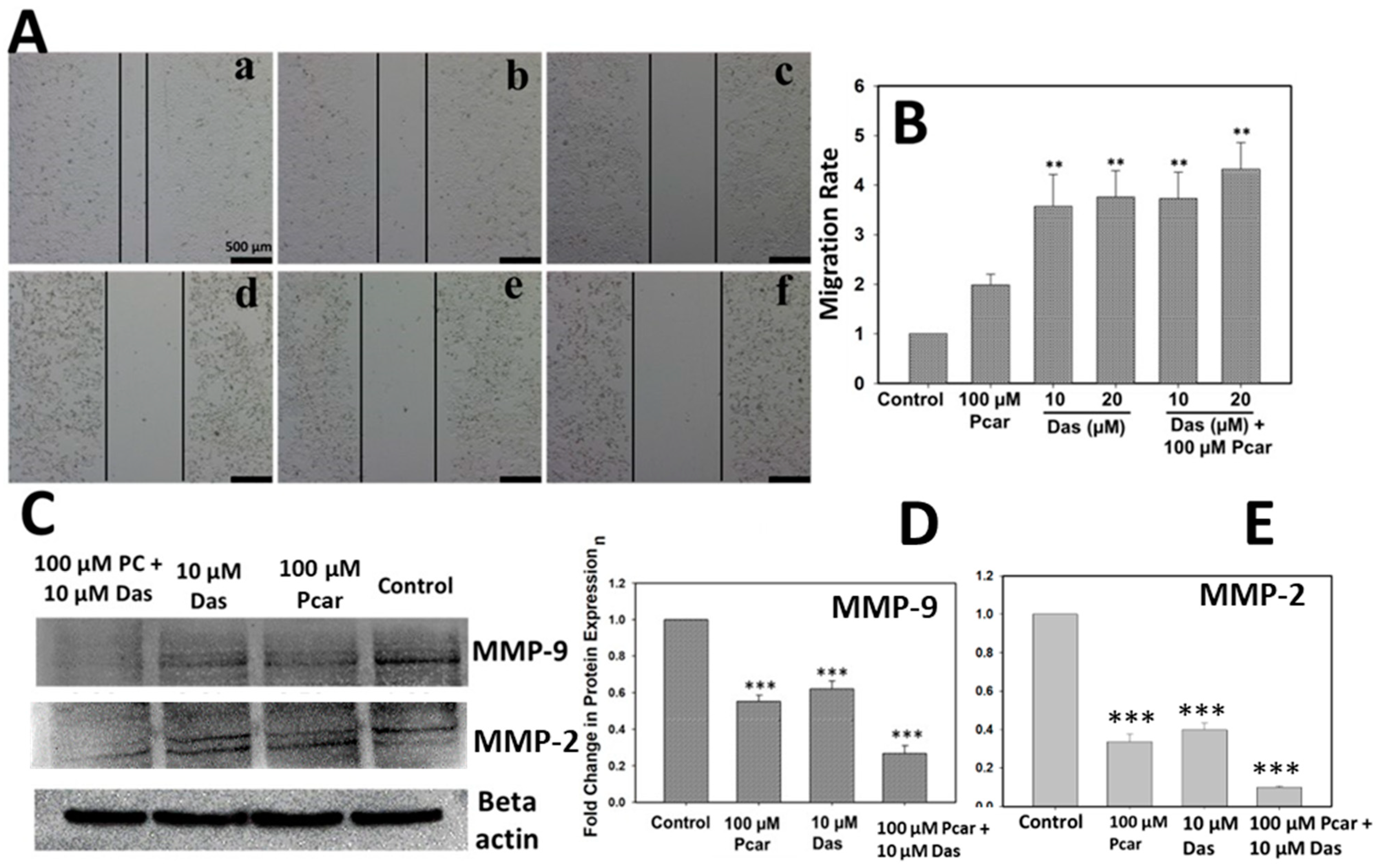
Cells were seeded in a 6-well plate and grown up to 90% confluency, and a linear wound in the cell monolayer was created using a 200 µL tip, followed by washing with phosphate buffer saline (PBS) to eliminate detached cells and exposure to 10 and 20 µM Das and 100 µM Pcar, alone and in combination, for 48 h. Edge movement of cells was then followed by using microscopy.
2.7. Nuclear Fragmentation
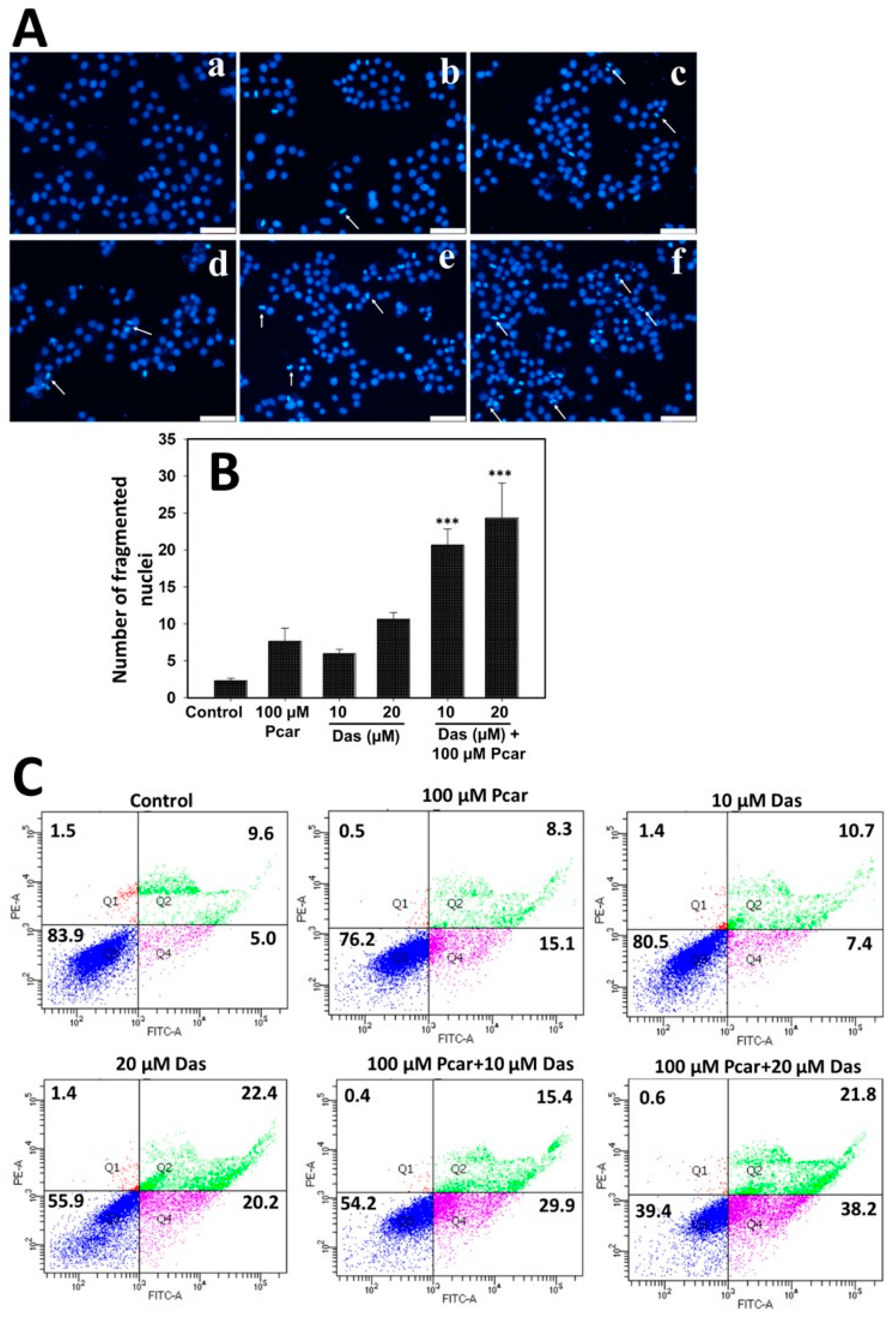
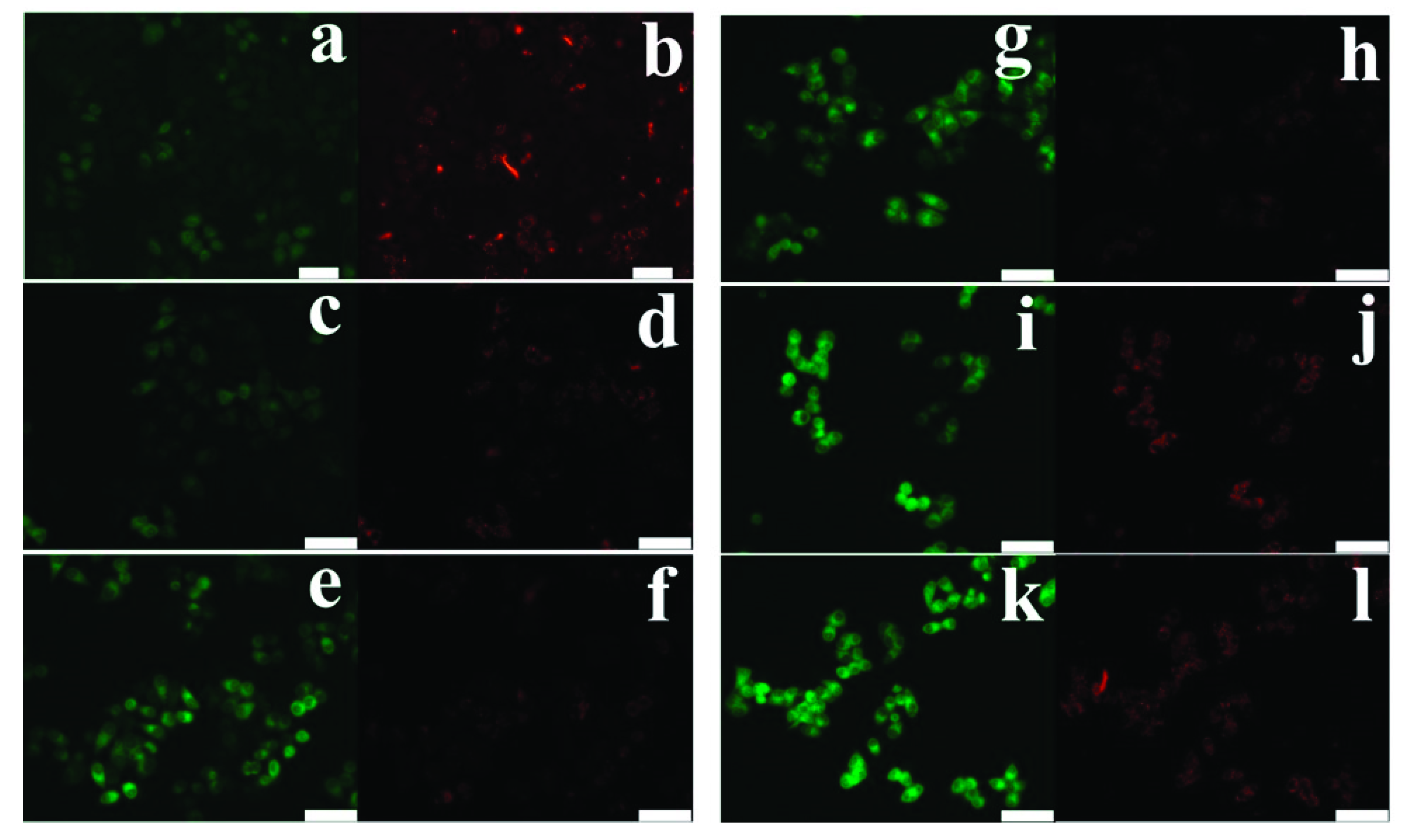
In cells treated with Das and Pcar, nuclear fragmentation was evaluated by DAPI staining. For this purpose, 5 × 104 cells were seeded in 12-well plates, followed by exposure to 10 and 20 µM Das and 100 µM Pcar, alone and in combination, for 24 h. Following the completion of the exposure period, the medium was withdrawn, and cells were washed in PBS and stained for 10 min with DAPI (1 g/mL in methanol). Fluorescence microscopy was used to capture images after the dye was removed, and cells were rinsed with PBS to remove background staining. By counting three independent areas, a graph was created.
2.8. Intracellular Reactive Oxygen Species Generation
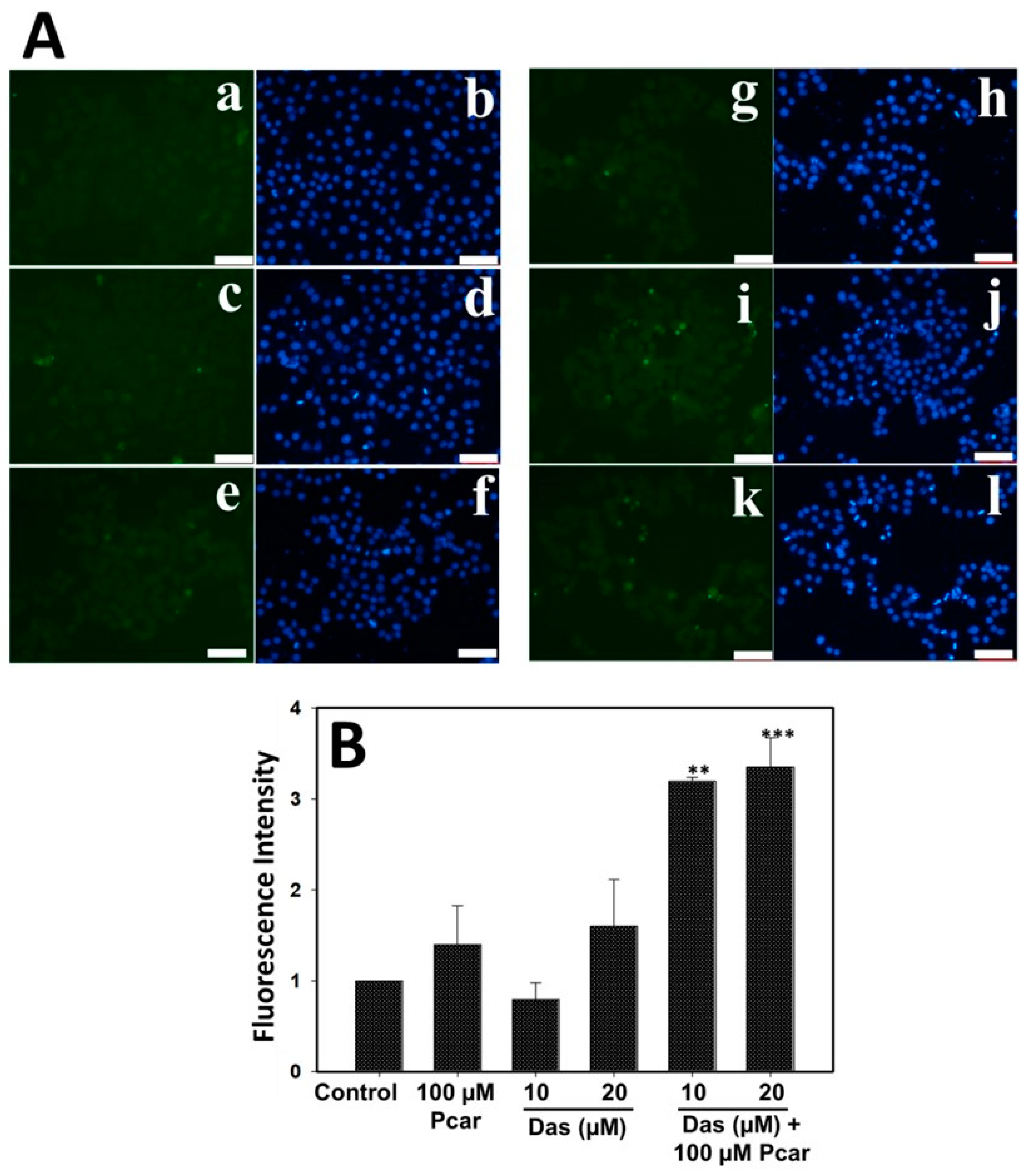
First, 5 × 104 cells were seeded in 12-well plates and incubated for 24 h, followed by exposure to 10 and 20 µM Das and 100 µM Pcar, alone and in combination, for 24 h. Following the completion of the exposure period, media were removed, and cells were washed with PBS and then stained with H2DCFDA dye (2, 7-dichlorofluorescein diacetate) (20 µM in PBS) for 30 min at 37 °C. In order to remove any more background staining, the dye was withdrawn and cells were rinsed with PBS before fluorescence microscopy was used to capture the images.
2.9. Mitochondrial Membrane Potential
JC-1 dye was used to estimate the mitochondrial membrane potential of cells. For this purpose, 5 × 104 cells were seeded in 12-well plates and incubated for 24 h, followed by exposure to 10 and 20 µM Das and 100 µM Pcar, alone and in combination, for 24 h. After completion of exposure time, media were removed, and cells were washed with PBS and then stained with JC-1 dye (20 µM in PBS) for 20 min at 37 °C. Cells were washed with PBS to remove background staining, and images were acquired using fluorescence microscopy.
2.10. Lysotracker
Lysotracker red was used to evaluate lysosomal accumulation. For this purpose, 5 × 104 cells were seeded in 12-well plates and incubated for 24 h, followed by exposure to 10 and 20 µM Das and 100 µM Pcar, alone and in combination, for 24 h. After the medium had been removed and the cells had been rinsed with PBS, Lysotracker red was used to stain them (75 nM). Images were taken using fluorescence microscopy after the cells were washed in PBS to remove any remaining background staining.
2.11. Apoptosis
Apoptosis in cancerous cells was detected by using an annexin V/fluorescein isothiocyanate (FITC) detection kit (BD Biosciences). For this purpose, 2.0 × 104 cells were seeded in a 6-well plate and treated with 10 and 20 µM Das and 100 µM Pcar for 24 h. After completion of exposure time, media were removed, trypsinized, washed with PBS, and incubated with 100 µL of binding buffer containing 5 µL PI and FITC for 15 min in the dark. The sample was supplemented with 400 µL of binding buffer, and the results were obtained using a flow cytometer with red and green channels.
2.12. Western Blot
For protein isolation, 2 × 105 cells were seeded in a 6-well plate and treated with 10 µM Das and 100 µM Pcar in combination or alone. Media were removed and cells were washed with ice-cold PBS, harvested, and lysed by sonication in the lysis buffer consisting of PMSF protease inhibitor. Isolated protein was quantified using the bicinchoninic acid (BCA) method (Pierce, BCA Protein Assay kit, Burlington, MA, USA). Protein (30 µg) was loaded on 12% SDS PAGE and transferred to a polyvinylidene difluoride (PVDF) membrane at 300 mV for 3 h. The membrane was blocked with 5% BSA, washed with Tris-buffered saline (TBST), and then incubated with specific primary antibodies overnight at 4 °C. Membranes were washed thrice with TBST and incubated with specific secondary antibodies for 2 h. Immobilon Western-HRP substrate (Millipore, Billerica, MA, USA) was used to develop the blots. MMP-9, MMP-2, cleaved caspase-3, cleaved poly (ADP-ribose) polymerase (PARP), cleaved caspase-6, pAKT, AKT, cleaved caspase-7, and phospho C-raf antibody antibodies were purchased from Cell Signaling Technology, with catalog numbers #3852, #4022, #9661, #9541, #9761, #9275, #4691, #9491, and #9421. Anti-LC3 antibody was purchased from Sigma-Aldrich, Burlington, MA, USA with catalog No. L7543-25UL. Anti-Src and anti-Src (phospho Y419) antibodies were obtained from Abcam, having catalog numbers ab47405 and ab185617. Primary antibodies were diluted at 1:1000 in the experiment. A secondary antibody, having catalog number #7074, was obtained from Cell Signaling Technology and diluted to 1:2000 in the experiment.
2.13. In Vivo Experiment
Male nude mice (4–6 weeks old) were used for the development of the ectopic HepG2 tumor models in the right flank of each mouse. The protocol was approved by the Animal Ethical Committee of Liaocheng University, China. HepG2 cells were mixed with matrigel in a 1:1
v:
v ratio and kept in ice until use, to prevent the gelation of matrigel. One hundred microliters of a solution containing 1 × 10
5 HepG2 cells was injected subcutaneously into the right flank of each mouse and observed daily to check the tumor development. Mouse tumor size was measured externally by using a digital caliper, and drug exposure was started when tumor size reached 50–100 mm
3. Mice were divided into four groups and given an intraperitoneal injection two times a week for 5 weeks with a solution containing (i) 0.9% saline buffer (control group), (ii) 2 mg/kg Pcar in PBS, (iii) 10 mg/kg DAS in PBS, or (iv) combined treatment of 2 mg/kg Pcar and 10 mg/kg DAS in PBS. At the end of the experiment, animals were euthanized by the cervical dislocation method. All efforts were made to minimize the suffering of animals. Tumor sizes were measured by using the formula (length) × (width)
2 × 1/2.
Table S1 shows the development of tumor in given time period.
2.14. Statistical Analysis
One-way ANOVA (GraphPad Prism 5.0) was used to perform the statistical analysis and determine the significance of the results of in vitro and in vivo experiments. Three independent experiments were performed in each case, and data are presented as the mean ± standard error (SE).
4. Discussion
The present study suggested that Pcar and Das acted synergistically to inhibit the growth of liver cancer cells in both in vitro and in vivo models. HCC is reported to be a hypervascular tumor type having an enhanced tendency for angiogenesis and neovascularization [
21]. In order to overcome the single-treatment resistance in cancer cells, combined therapy is used in this study to enhance the treatment effect and reduce the dose-responsive side effects [
22,
23]. In this regard, Xiao et al. demonstrated that Das in combination with paclitaxel significantly inhibited the proliferation and induced apoptosis in ovarian cancer cells. Further, an in vivo study also showed that combined treatment of Das and paclitaxel has a tumor growth inhibition rate of 76.7% and 58.5% for mice xenografted with A2780 and HO8910 cells, respectively [
24]. Other studies also supported the fact that Das in combination with other drugs provides an additive or synergistic anti-tumor effect [
1,
6,
25]. Similarly, acetylcarnitine in combination with butyrate has anticarcinogenic properties superior to those of butyrate alone and is shown to inhibit the survival of colon cancer in vitro [
26]. Based on these observations, the combined toxic effect of Pcar and Das was evaluated, and the result showed that the combined treatment exhibits enhanced anti-tumor effect in terms of apoptosis induction, cytotoxicity, membrane depolarization, metastasis, and tumor growth.
Various reports demonstrate the toxic effect of Das and Pcar on cancerous cells through different mechanisms. Thus, in the present study, the cytotoxic effect of Pcar and Das on HepG2 cells was first evaluated by MTT assay. The in vitro study showed that the growth of the HepG2 cell line was slightly inhibited by single treatments of Das and Pcar exposure, but a marked significant reduction was recorded in the Pcar and Das combination treatment group. Further, the synergistic interaction between Pcar and Das was confirmed by the CI value (0.83) being less than one, which clearly indicates the synergistic interaction between the two drugs. Thus, it can be seen that the combined treatment of Pcar and Das proved to be more toxic to liver cancer cells and induced more cell death in comparison to single-drug treatments. Similar to our result, combined treatment of acetylcarnitine and butyrate is also reported to inhibit the survival of SW480 colon cancer cells more significantly than single-drug exposure. The clonogenic assay is also an in vitro cell survival assay that shows the ability of single cells to grow and form a colony [
27]. Another report shows that Das treatment significantly inhibited the colony formation efficiency of ovarian cancer cells [
28]. Thus, to evaluate the combined effect of Das and Pcar on the colony formation efficiency of HepG2 cells, a clonogenic assay was performed. Results revealed that combined exposure to Pcar and Das inhibited the colony formation efficiency of HepG2 cells to a greater degree than single-drug treatment. Thus, the above results in combination with other observations suggest that Pcar and Das in combination act synergistically and can efficiently inhibit the growth and proliferation of cancerous cells.
Earlier it was reported that exposure of Pcar and carnitine to HT-29 causes O
2− production in the mitochondria of cancerous cells that promotes apoptosis initiation [
14]. Another report also suggests that cancerous cells are susceptible to Pcar exposure due to the production of H
2O
2 [
15]. Following the above observations, our study also showed that Pcar and Das combined exposure induces ROS generation in HepG2 cells which further leads to nuclear fragmentation and apoptosis in liver cancer cells. It has been well reported that increased H
2O
2 production in cancerous cells can trigger deleterious cellular effects such as caspase activation [
15]. Results also showed that combined exposure to Pcar and Das increased the expression of cleaved caspase-3, -7, and -6, which are apoptosis-related proteins. It has also been reported that during apoptosis, PARP is known to be cleaved by caspase, which is an important marker of caspase-dependent apoptosis [
29]. As expected, PARP cleavage was increased significantly upon Pcar and Das combined exposure in comparison to single-drug treatment, thus confirming its crucial role in the apoptotic process. Additionally, a number of chemotherapeutic treatments are known to change the integrity of the mitochondrial membrane, which results in a collapse in the membrane potential [
30]. Thus, in the present study, the membrane potential of HepG2 cells in response to Pcar and Das combined treatment was evaluated, and the result clearly showed that combined treatment induces membrane depolarization to a greater degree than single treatment. Thus, the crucial role of mitochondria in the apoptotic process and cell death induced by Pcar and Das is confirmed.
Das is also reported to induce autophagic cell death mediated by Beclin-1, AKT, and Bcl-2 in ovarian cancer cells. Das-induced autophagy is also reported to be responsible for the Das-induced inhibition of cell growth [
28]. Das treatment significantly increased the levels of autophagosomes and autophagolysosomes in the cytoplasm of cancerous cells [
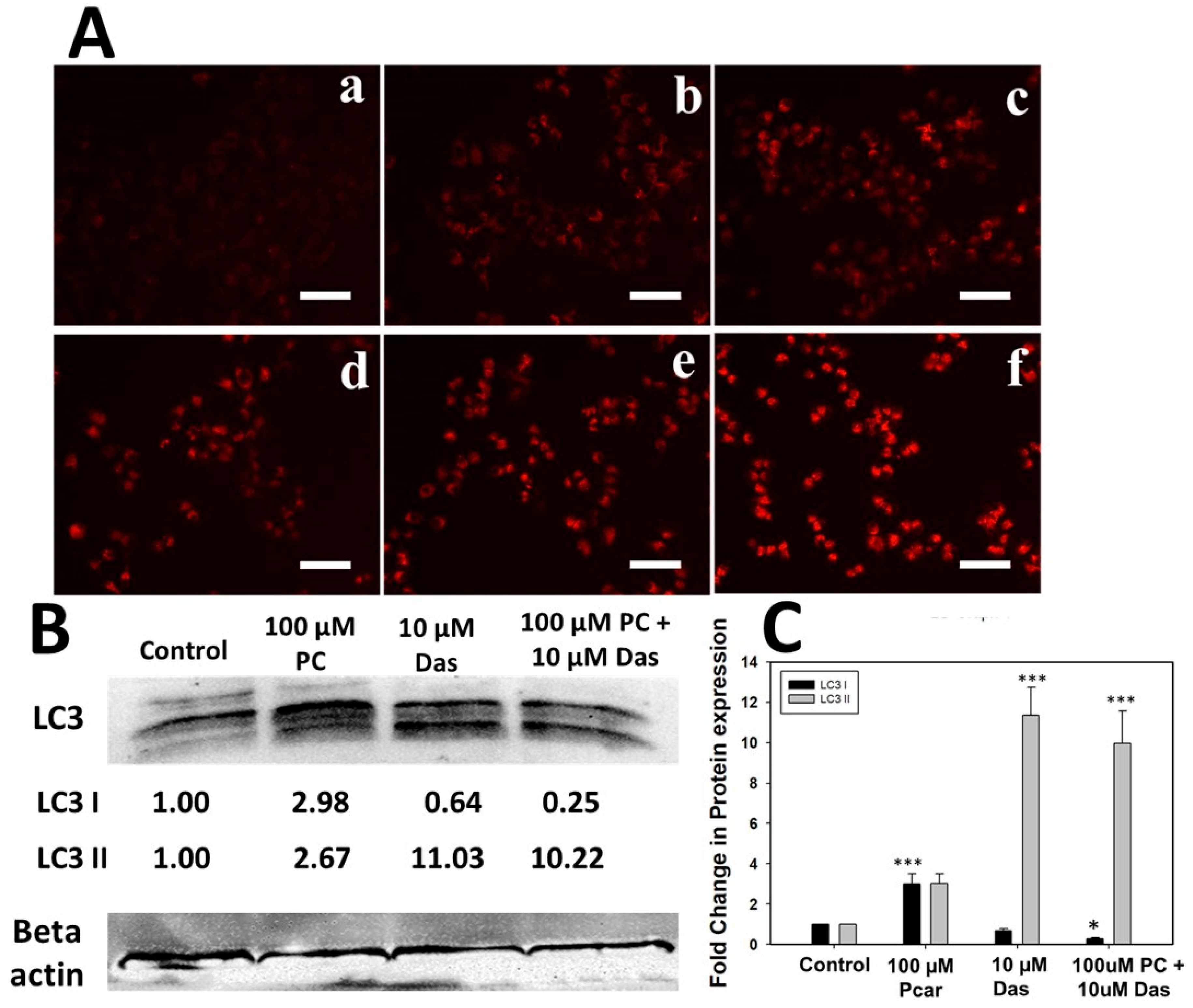
28]. Similar to this, our study also demonstrated autophagy induction in liver cancer cells, as shown by an accumulation of Lysotracker and overexpression of LC3 protein (
Figure 6). Feng Le reported that Das-induced autophagy partially depends on the AKT pathway and Bcl-2 expression [
28]. Thus, in the present study, the dependency of autophagy induced by Pcar and Das in liver cancer cells has been investigated. Results showed that combined treatment of Pcar and Das reduced the phosphorylation of Akt, which in accordance with the above result, could be considered as one of the factors that induce autophagy in cancerous cells.
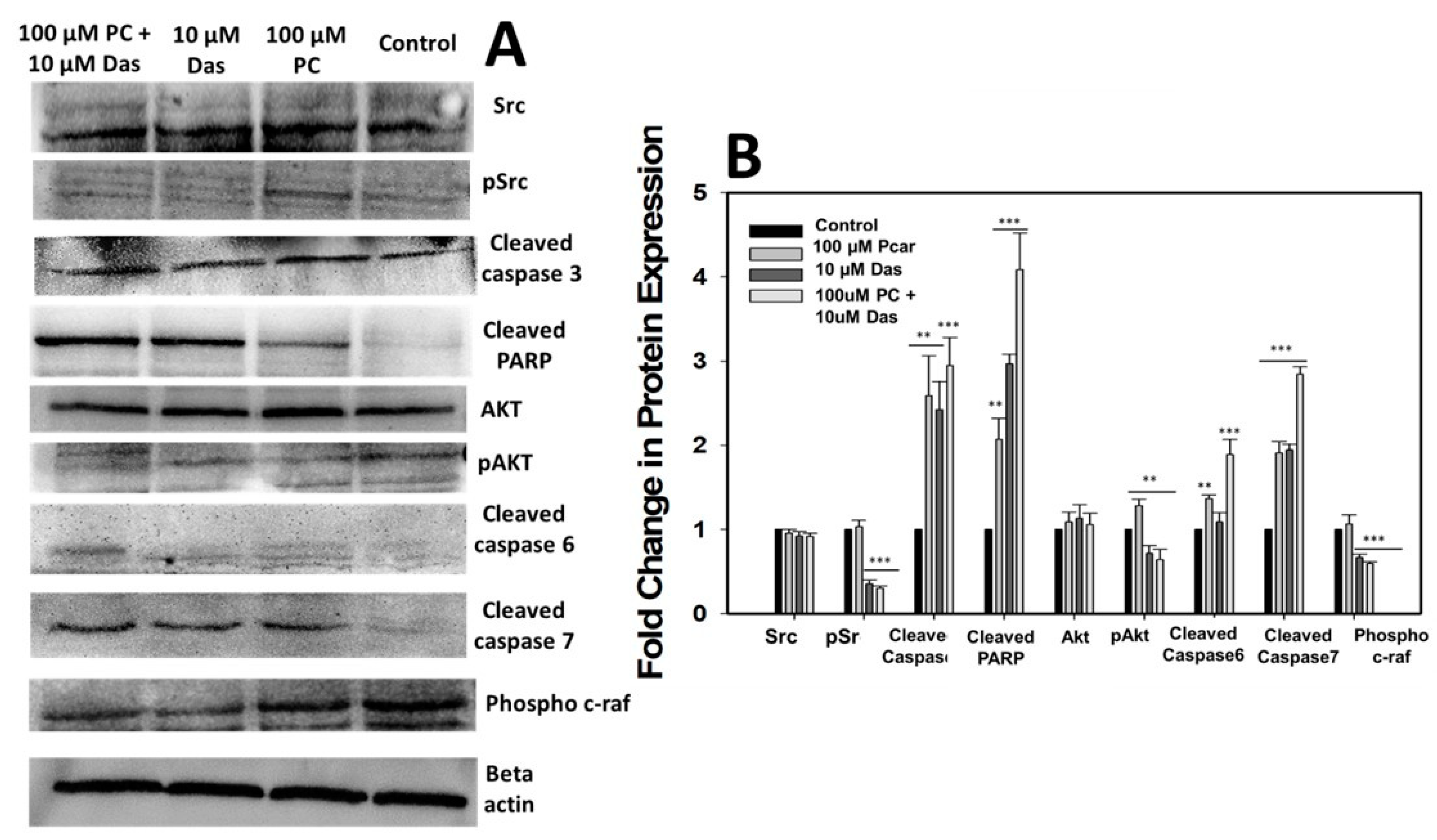
To reveal the further molecular mechanisms involved in the Pcar and Das anti-tumor effect, several proteins involved in cancer signaling pathways were also analyzed. Results showed that Pcar and Das downregulate several cancer-related proteins and upregulate the expression of apoptotic proteins. Mayor and Krop demonstrated that Src is overexpressed in numerous cancers and can regulate several signaling pathways that affect the proliferation, survival, invasion, migration, and angiogenesis in tumor cells [
31]. Further, Sayed et al. showed that Das in combination with rosuvastatin acts synergistically to suppress the growth of HCC by decreasing the expression of p-Src [
1]. Likewise, our result also suggests the decreased phosphorylation of Src upon Pcar and Das combined exposure. Other reports also showed that the chemotherapeutic effect of Das is primarily due to the inhibition of Src expression, which can also provide more sensitivity to oxaliplatin in cancer treatment. Contrary to this, cancer cells that did not show high Src expression failed to show the synergistic action between the two [
32]. Our finding also supports the idea that cancer cells highly dependent on Src signaling are more susceptible to Das in combination therapy (Src-targeted therapy).
Das is a Bcr-Abl TKI and SRC family tyrosine kinase inhibitor that has been approved for the treatment of chronic myelogenous leukemia, Philadelphia chromosome-positive acute lymphoblastic leukemia, and alveolar soft part sarcoma [
6,
7,
33]. In vivo studies also showed that Das exposure either in combination or alone reduces tumor size [
6,
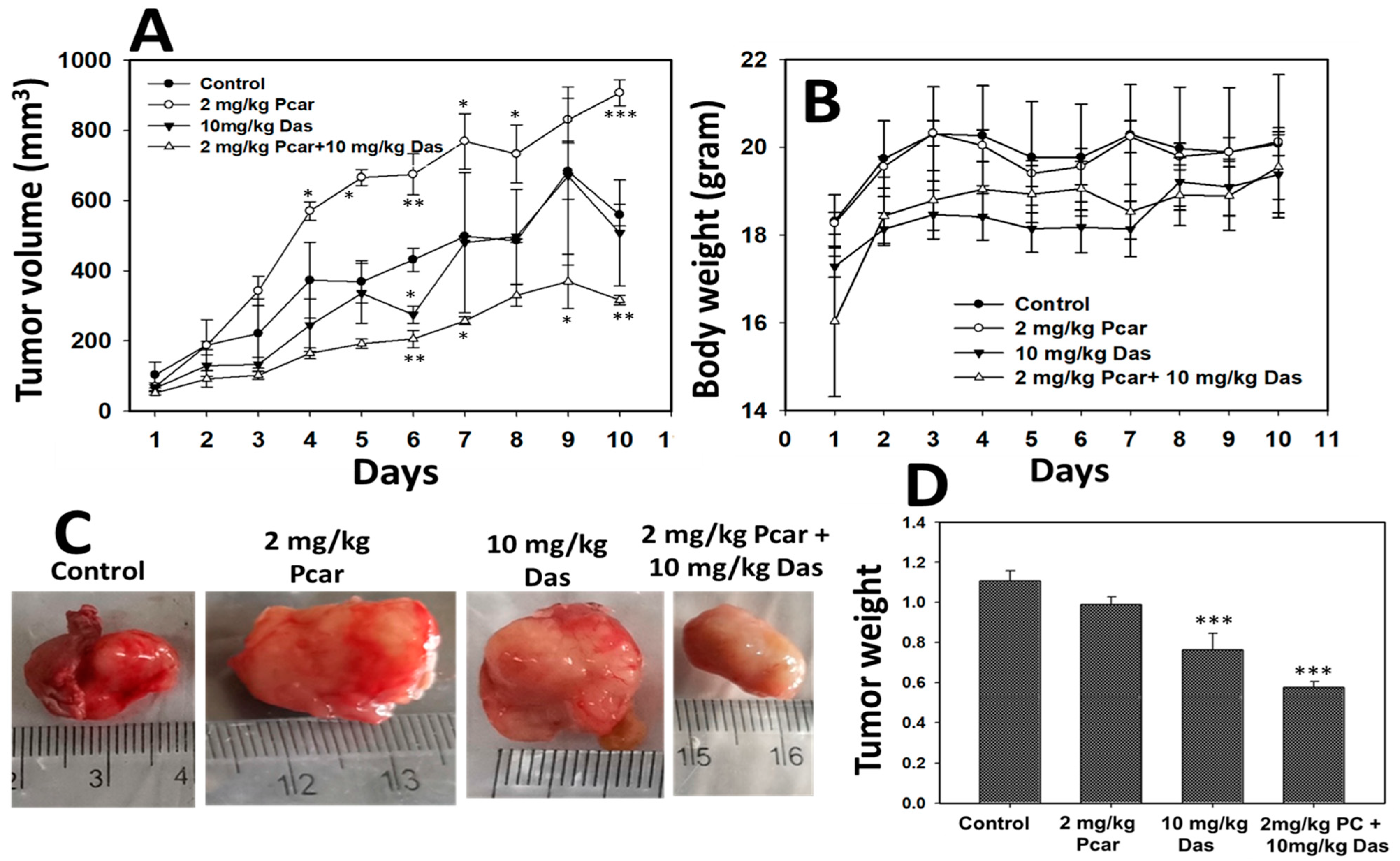
28]. Following the above observation, our result also showed that exposure of mice to Das and Pcar significantly reduces the tumor volume in comparison to a single treatment of Das.
Figures S4 and S5 shows the tumor migration in a mouse treated with only Das, which further leads to mouse death and mouse survival graph. The percentage of death in the group treated with only Das is recorded as 37.5%, whereas it is reduced to 20% for the Pcar and Das combination treatment group. This result also confirms that in comparison to single treatment, combined drug treatment shows more efficiency as an anti-tumor agent.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







