
Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
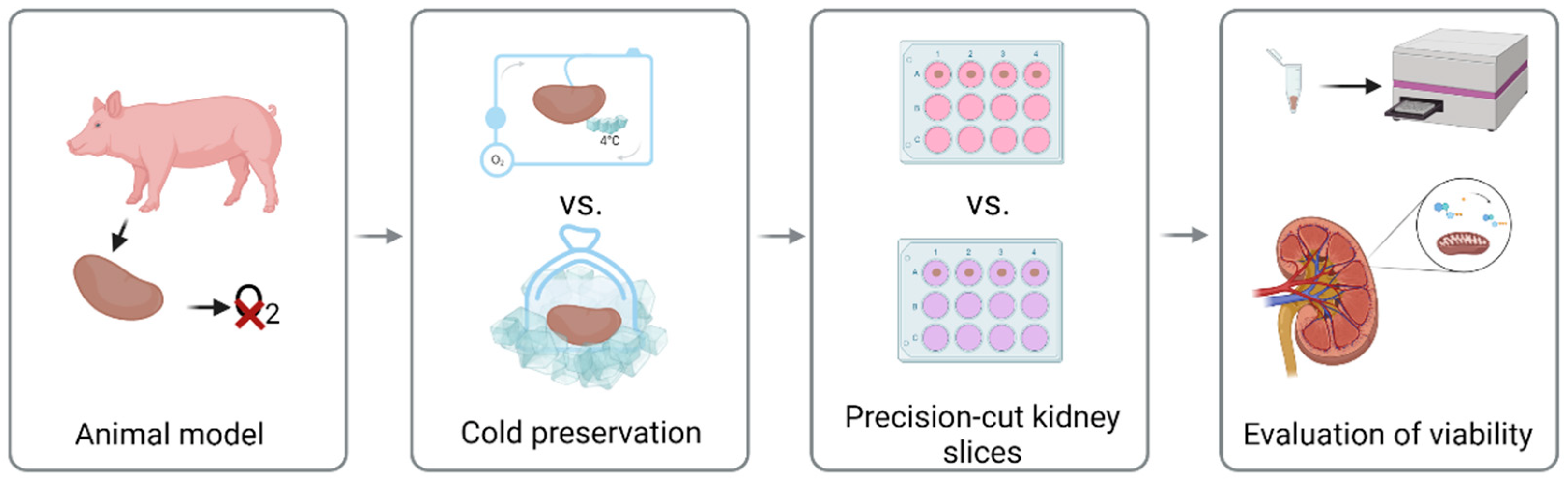
2.1. Experimental Design
2.2. Animal Model
2.3. Cold Preservation
2.4. Precision-Cut Kidney Slices
2.5. Evaluation of Viability
2.6. Histological Analysis
2.7. Determination of Fungal Infection
2.8. Statistical Analysis
3. Results
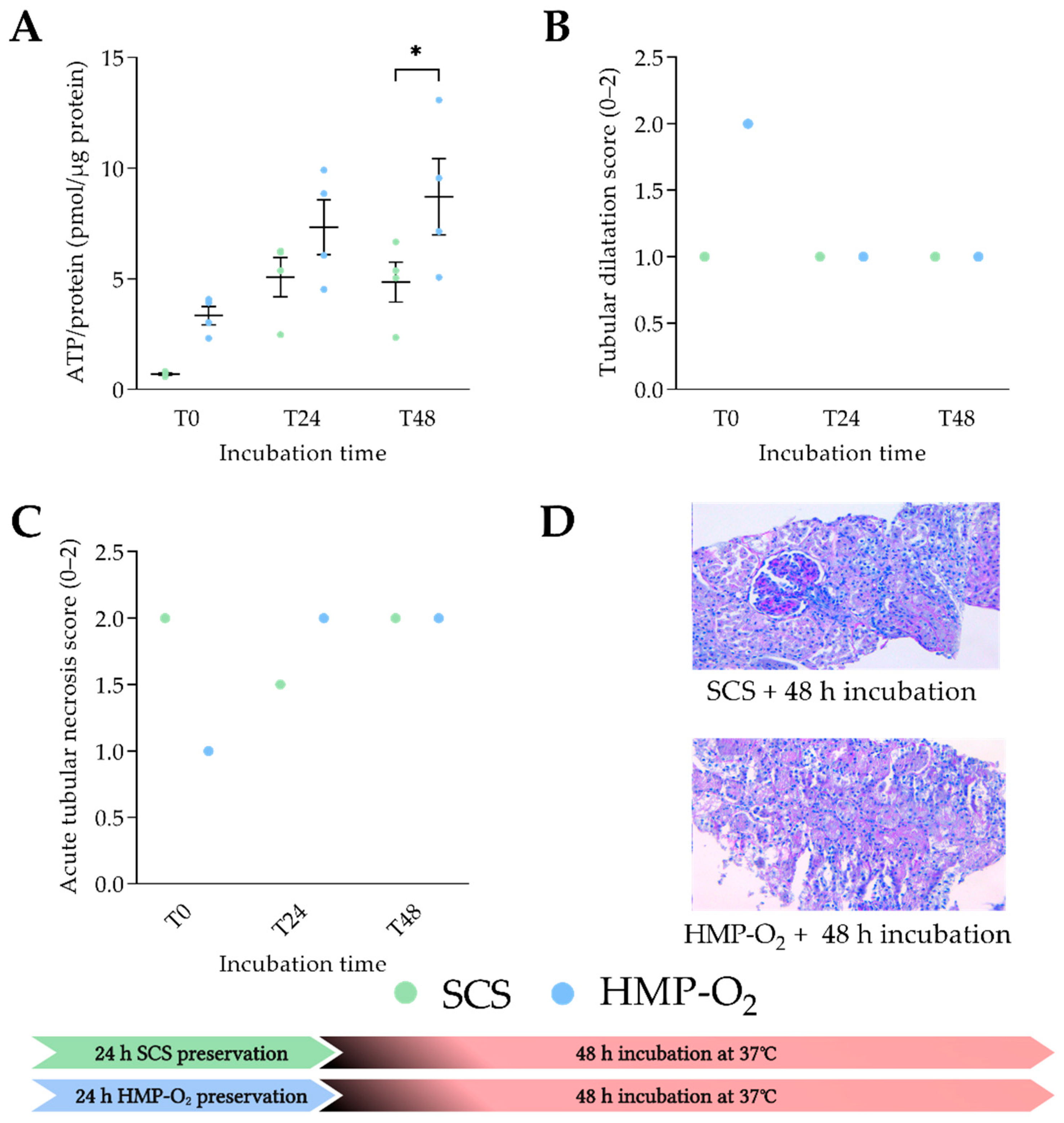
3.1. HMP-O2 as Cold Preservation Results in Significant Higher PCKS Viability
3.2. Porcine PCKS Remain Viable up to 72 h
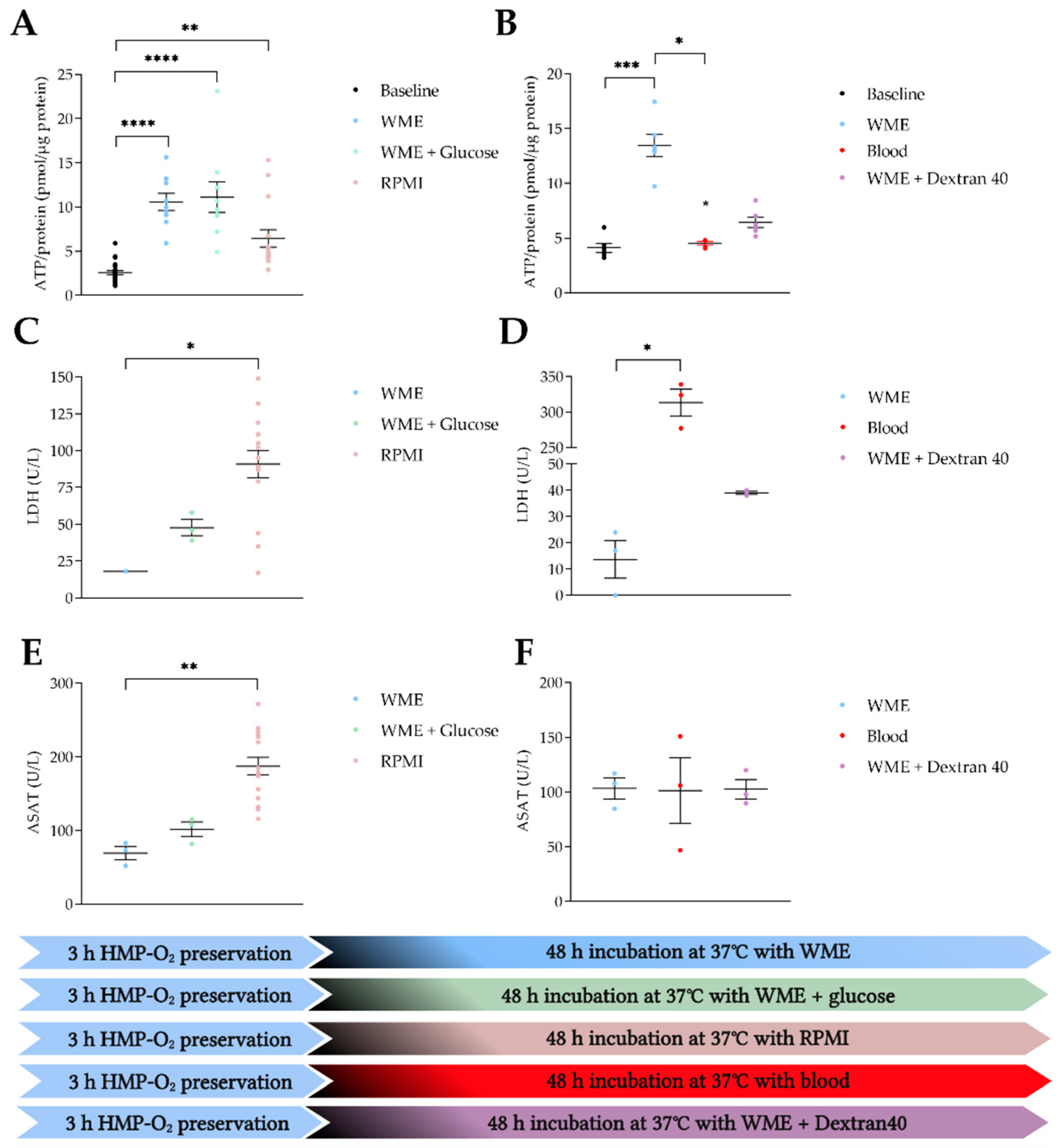
3.3. WME Provides the Most Beneficial Incubation Conditions
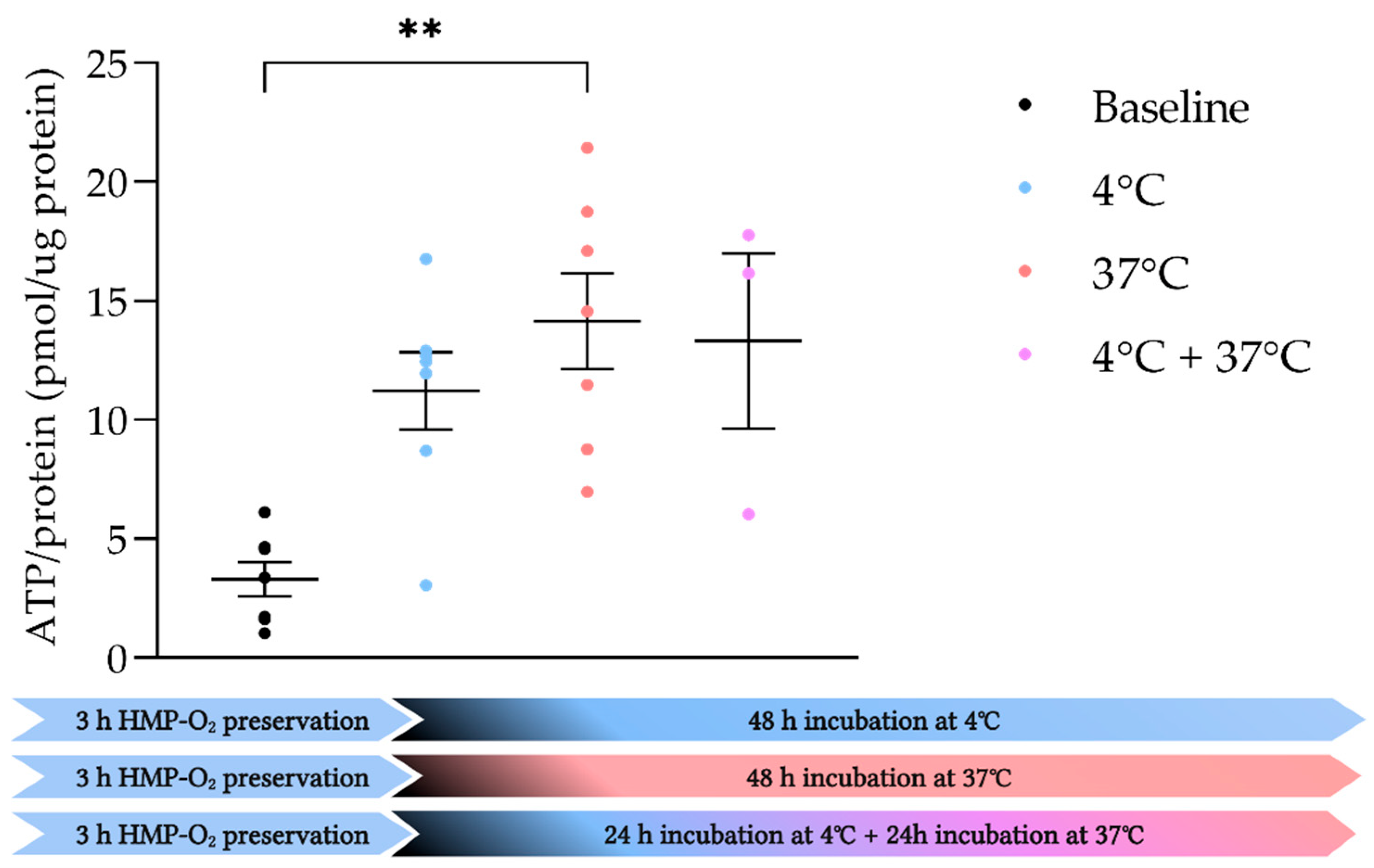
3.4. PCKS Remain Viable under Both Hypothermic and Normothermic Conditions
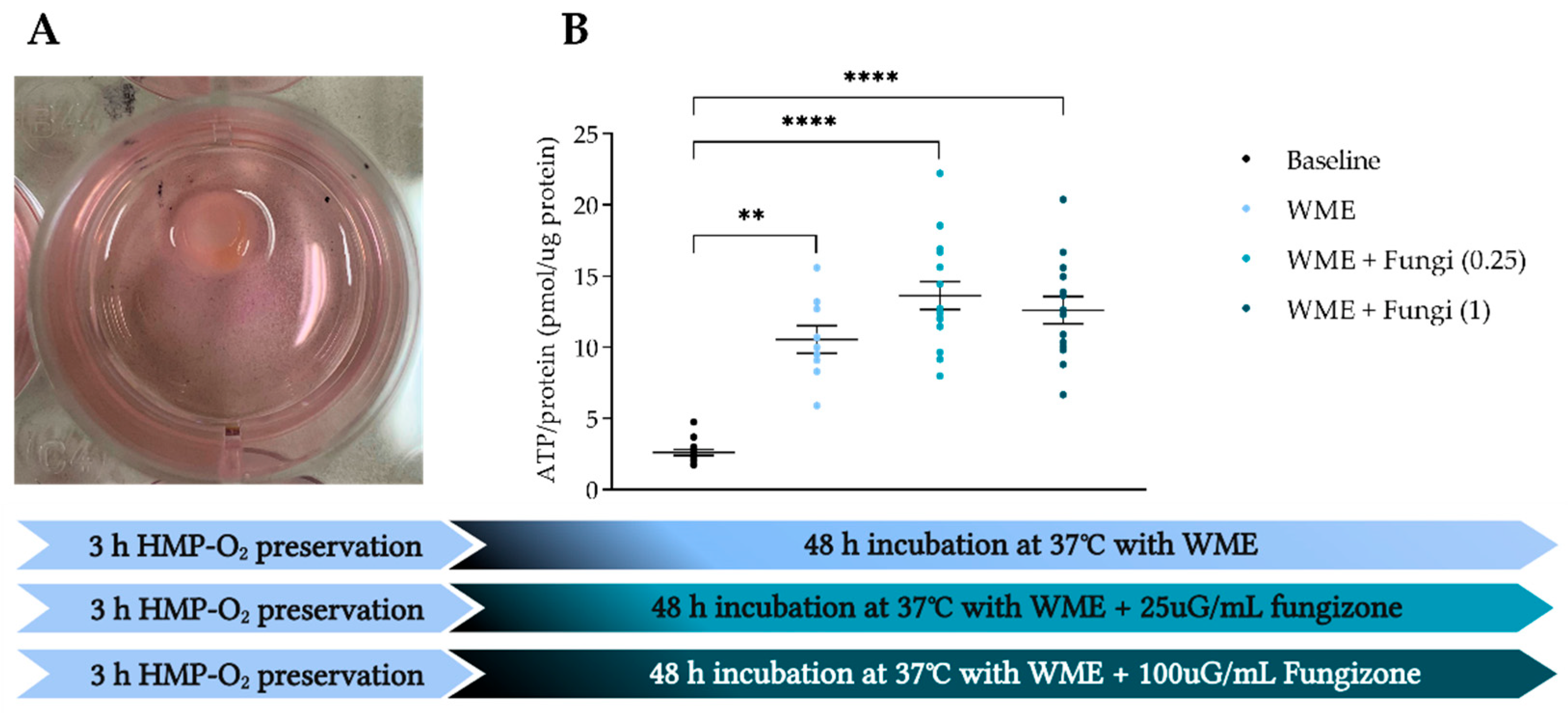
3.5. Amfotericine B Is Safe to Add to Prevent Fungal Infections
4. Discussion
4.1. Oxygenated HMP Provides an Excellent Foundation for Viable Porcine PCKS
4.2. WME Best Supports the Renal Metabolism of Porcine PCKS
4.3. Porcine PCKS Can Be Incubated under Both Hypo- and Normothermic Conditions
4.4. ATP as a Biomarker for Renal Tissue Viability
4.5. Advantages and Disadvantages of Porcine PCKS
4.6. Future Perspectives Porcine PCKS in Translation to NMP
4.7. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
Appendix A
| Perfusate Composition: | |
| Heparinized and leukocyte-depleted autologous blood | |
| Red blood cells: | 360 mL |
| Plasma: | 475 mL |
| Amoxicillin/Clavulanic acid (1000 mg/200 mg) | 10 mG |
| 8.4% Sodium bicarbonate (B. Braun) | 15.25 mL |
| 5% Glucose (Baxter) | 12.50 mL |
| Dexamethasone (Centrafarm) | 8.3 mG |
| Mannitol (Baxter) | 10 mG |
| Creatinine (Merck) | 135 mG |
| Sodium nitroprusside (Merck) | 2.7 mG |
| Aminoplasmal (B. Braun) | 90 mL |
| Insulin (100 IU/mL) (Novorapid) | 0.186 mL |
| 250 µg/mL Amfotericine B (Fungizone) (Merck) | 1 mL |
References
- Gavriilidis, P.; Inston, N.G. Recipient and allograft survival following donation after circulatory death versus donation after brain death for renal transplantation: A systematic review and meta-analysis. Transplant. Rev. 2020, 34, 100563. [Google Scholar] [CrossRef] [PubMed]
- Summers, D.M.; Watson, C.J.E.; Pettigrew, G.J.; Johnson, R.J.; Collett, D.; Neuberger, J.M.; Bradley, J.A. Kidney donation after circulatory death (DCD): State of the art. Kidney Int. 2015, 88, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, O.A.; Mohiuddin, S.S. Biochemistry, Oxidative Phosphorylation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Chaudhry, R.; Varacallo, M. Biochemistry, Glycolysis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Castaneda, M.; Swiatecka-Urban, A.; Mitsnefes, M.; Feuerstein, D.; Kaskel, F.; Tellis, V.; Devarajan, P. Activation of mitochondrial apoptotic pathways in human renal allografts after ischemiareperfusion injury. Transplantation 2003, 76, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Matas, A.J.; Gillingham, K.J.; Humar, A.; Kandaswamy, R.; Sutherland, D.E.R.; Payne, W.D.; Dunn, T.B.; Najarian, J.S. 2202 kidney transplant recipients with 10 years of graft function: What happens next? Am. J. Transplant. 2008, 8, 2410–2419. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Theruvath, T.; Kawai, T.; Tolkoff-Rubin, N.; Cosimi, A.B. Strategies to Improve Long-Term Outcomes after Renal Transplantation. N. Engl. J. Med. 2002, 346, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Hosgood, S.A.; Hoff, M.; Nicholson, M.L. Treatment of transplant kidneys during machine perfusion. Transpl. Int. 2021, 34, 224–232. [Google Scholar] [CrossRef]
- Resch, T.; Cardini, B.; Oberhuber, R.; Weissenbacher, A.; Dumfarth, J.; Krapf, C.; Boesmueller, C.; Oefner, D.; Grimm, M.; Schneeberger, S. Transplanting Marginal Organs in the Era of Modern Machine Perfusion and Advanced Organ Monitoring. Front. Immunol. 2020, 11, 631. [Google Scholar] [CrossRef]
- Moers, C.; Smits, J.M.; Maathuis, M.-H.J.; Treckmann, J.; Van Gelder, F.; Napieralski, B.P.; Van Kasterop-Kutz, M.; Van Der Heide, J.J.H.; Squifflet, J.-P.; Van Heurn, E.; et al. Machine perfusion or cold storage in deceased-donor kidney transplantation. N. Engl. J. Med. 2009, 360, 7–19. [Google Scholar] [CrossRef]
- Jochmans, I.; Moers, C.; Smits, J.M.; Leuvenink, H.G.D.; Treckmann, J.; Paul, A.; Rahmel, A.; Squifflet, J.-P.; van Heurn, E.; Monbaliu, D.; et al. Machine perfusion versus cold storage for the preservation of kidneys donated after cardiac death: A multicenter, randomized, controlled trial. Ann. Surg. 2010, 252, 756–762. [Google Scholar] [CrossRef]
- Patel, K.; Smith, T.B.; Neil, D.A.; Thakker, A.; Tsuchiya, Y.; Higgs, E.B.; Hodges, N.J.; Ready, A.R.; Nath, J.; Ludwig, C. The Effects of Oxygenation on Ex Vivo Kidneys Undergoing Hypothermic Machine Perfusion. Transplantation 2019, 103, 314–322. [Google Scholar] [CrossRef]
- Hosgood, S.A.; Brown, R.J.; Nicholson, M.L. Advances in Kidney Preservation Techniques and Their Application in Clinical Practice. Transplantation 2021, 105, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Jochmans, I.; Brat, A.; Davies, L.; Hofker, H.S.; van de Leemkolk, F.E.M.; Leuvenink, H.G.D.; Mchir, S.R.K.; Pirenne, J.; Ploeg, R. Oxygenated versus standard cold perfusion preservation in kidney transplantation (COMPARE): A randomised, double-blind, paired, phase 3 trial. Lancet 2020, 396, 1653–1662. [Google Scholar] [CrossRef]
- Jing, L.; Yao, L.; Zhao, M.; Peng, L.P.; Liu, M. Organ preservation: From the past to the future. Acta Pharmacol. Sin. 2018, 39, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Douglas, W.R. Of pigs and men and research: A review of applications and analogies of the pig, sus scrofa, in human medical research. Space Life Sci. 1972, 3, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Hosgood, S.A. Renal transplantation after ex vivo normothermic perfusion: The first clinical study. Am. J. Transplant. 2013, 13, 1246–1252. [Google Scholar]
- von Horn, C.; Minor, T. Improved approach for normothermic machine perfusion of cold stored kidney grafts. Am. J. Transl. Res. 2018, 10, 1921–1929. [Google Scholar]
- Hosgood, S.A.; Thompson, E.; Moore, T.; Wilson, C.H.; Nicholson, M.L. Normothermic machine perfusion for the assessment and transplantation of declined human kidneys from donation after circulatory death donors. Br. J. Surg. 2018, 105, 388–394. [Google Scholar] [CrossRef]
- Arykbaeva, A.S.; de Vries, D.K.; Doppenberg, J.B.; Engelse, M.A.; Hankemeier, T.; Harms, A.C.; Wijermars, L.G.; Schaapherder, A.F.; Bakker, J.A.; Ploeg, R.J.; et al. Metabolic needs of the kidney graft undergoing normothermic machine perfusion. Kidney Int. 2021, 100, 301–310. [Google Scholar] [CrossRef]
- Hamelink, T.L.; Ogurlu, B.; De Beule, J.; Lantinga, V.A.; Pool, M.B.; Venema, L.H.; Leuvenink, H.G.; Jochmans, I.; Moers, C. Renal Normothermic Machine Perfusion. Transplantation 2021, 106, 268–279. [Google Scholar] [CrossRef]
- Mazilescu, L.I.; Urbanellis, P.; Kim, S.J.; Goto, T.; Noguchi, Y.; Konvalinka, A.; Reichman, T.W.; Sayed, B.A.; Mucsi, I.; Lee, J.Y.; et al. Normothermic Ex Vivo Kidney Perfusion for Human Kidney Transplantation: First North American Results. Transplantation 2022. epub ahead of print. [Google Scholar] [CrossRef]
- Stribos, E.G.D.; Hillebrands, J.L.; Olinga, P.; Mutsaers, H.A.M. Renal fibrosis in precision-cut kidney slices. Eur. J. Pharmacol. 2016, 790, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Stribos, E.G.; Luangmonkong, T.; Leliveld, A.M.; de Jong, I.J.; van Son, W.J.; Hillebrands, J.-L.; Seelen, M.A.; van Goor, H.; Olinga, P.; Mutsaers, H. Precision-cut human kidney slices as a model to elucidate the process of renal fibrosis. Transl. Res. 2016, 170, 8–16.e1. [Google Scholar] [CrossRef] [PubMed]
- Obatomi, D.K.; Thanh, N.T.K.; Brant, S.; Bach, P.H. The toxic mechanism and metabolic effects of atractyloside in precision-cut pig kidney and liver slices. Arch. Toxicol. 1998, 72, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Roskott, A.M.; Nieuwenhuijs, V.B.; Leuvenink, H.G.D.; Dijkstra, G.; Ottens, P.; de Jager, M.H.; Pereira, P.G.D.; Fidler, V.; Groothuis, G.M.M.; Ploeg, R.J.; et al. Reduced ischemia-reoxygenation injury in rat intestine after luminal preservation with a tailored solution. Transplantation 2010, 90, 622–629. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, I.A.M.; Olinga, P.; de Jager, M.H.; Merema, M.T.; de Kanter, R.; van de Kerkhof, E.G.; Groothuis, G.M.M. Preparation and incubation of precision-cut liver and intestinal slices for application in drug metabolism and toxicity studies. Nat. Protoc. 2010, 5, 1540–1551. [Google Scholar] [CrossRef] [PubMed]
- Tingle, S.J.; Figueiredo, R.S.; Moir, J.A.; Goodfellow, M.; Thompson, E.R.; Ibrahim, I.K.; Bates, L.; Talbot, D.; Wilson, C.H. Hypothermic machine perfusion is superior to static cold storage in deceased donor kidney transplantation: A meta-analysis. Clin. Transplant. 2020, 34, e13814. [Google Scholar] [CrossRef]
- Foucher, Y.; Fournier, M.-C.; Legendre, C.; Morelon, E.; Buron, F.; Girerd, S.; Ladrière, M.; Mourad, G.; Garrigue, V.; Glotz, D.; et al. Comparison of machine perfusion versus cold storage in kidney transplant recipients from expanded criteria donors: A cohort-based study. Nephrol. Dial. Transplant. 2020, 35, 1051–1059. [Google Scholar] [CrossRef]
- Bellini, M.I.; Charalampidis, S.; Herbert, P.E.; Bonatsos, V.; Crane, J.; Muthusamy, A.; Dor, F.J.M.F.; Papalois, V. Cold pulsatile machine perfusion versus static cold storage in kidney transplantation: A single centre experience. BioMed Res. Int. 2019, 2019, 7435248. [Google Scholar] [CrossRef]
- Brat, A.; de Vries, K.M.; van Heurn, E.W.E.; Huurman, V.A.L.; de Jongh, W.; Leuvenink, H.G.D.; van Zuilen, A.D.; Haase-Kromwijk, B.J.J.M.; de Jonge, J.; Berger, S.P.; et al. Hypothermic Machine Perfusion as a National Standard Preservation Method for Deceased Donor Kidneys. Transplantation 2021. epub ahead of print. [Google Scholar] [CrossRef]
- Garonzik-Wang, J.M.; Lonze, B.E.; Ruck, J.M.; Luo, X.; Massie, A.B.; Melancon, K.; Burdick, J.F.; Segev, R.L.; Sun, Z. Mitochondrial membrane potential and delayed graft function following kidney transplantation. Am. J. Transplant. 2019, 19, 585–590. [Google Scholar] [CrossRef]
- Moers, C.; Leuvenink, H.G.D.; Ploeg, R.J. Non-heart beating organ donation: Overview and future perspectives. Transpl. Int. 2007, 20, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Venema, L.H.; van Leeuwen, L.L.; Posma, R.A.; van Goor, H.; Ploeg, R.J.; Hannaert, P.; Hauet, T.; Minor, T.; Leuvenink, H.G. Impact of Red Blood Cells on Function and Metabolism of Porcine Deceased Donor Kidneys During Normothermic Machine Perfusion. Transplantation 2021. epub ahead of print. [Google Scholar] [CrossRef]
- Quack, M.N.G. van Kampen: Stochastic Processes in Physics and Chemistry. North Holland Publishing Company, Amsterdam 1981. 419 Seiten, Preis: $ 76.50. Ztschrift fr Elektrochemie, Berichte der Bunsengesellschaft fr Physikalische Chemie 1983, 87, 370. [Google Scholar] [CrossRef]
- Aldecoa, C.; Llau, J.V.; Nuvials, X.; Artigas, A. Role of albumin in the preservation of endothelial glycocalyx integrity and the microcirculation: A review. Ann. Intensive Care 2020, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, I.A.M.; Tajima, O.; Groten, J.P.; Wolterbeek, A.P.M. Intercellular communication and cell proliferation in precision-cut rat liver slices: Effect of medium composition and DDT. Cancer Lett. 2000, 154, 53–62. [Google Scholar] [CrossRef]
- Coskun, A.; Baykal, A.T.; Kazan, D.; Akgoz, M.; Senal, M.O.; Berber, I.; Titiz, I.; Bilsel, G.; Kilercik, H.; Karaosmanoğlu, K.; et al. Proteomic Analysis of Kidney Preservation Solutions Prior to Renal Transplantation. PLoS ONE 2016, 11, e0168755. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.J. Relationship between energy requirements for Na+ reabsorption and other renal functions. Kidney Int. 1986, 29, 32–40. [Google Scholar] [CrossRef]
- Longchamp, A.; Klauser, A.; Songeon, J.; Agius, T.; Nastasi, A.; Ruttiman, R.; Moll, S.; Meier, R.; Buhler, L.; Corpataux, J.-M.; et al. Ex Vivo Analysis of Kidney Graft Viability Using 31P Magnetic Resonance Imaging Spectroscopy. Transplantation 2020, 104, 1825–1831. [Google Scholar] [CrossRef]
- Bellini, M.I.; Yiu, J.; Nozdrin, M.; Papalois, V. The Effect of Preservation Temperature on Liver, Kidney, and Pancreas Tissue ATP in Animal and Preclinical Human Models. J. Clin. Med. 2019, 8, 1421. [Google Scholar] [CrossRef]
- Stribos, E.G.D.; Seelen, M.A.; van Goor, H.; Olinga, P.; Mutsaers, H.A.M. Murine Precision-Cut Kidney Slices as an ex vivo Model to Evaluate the Role of Transforming Growth Factor-β1 Signaling in the Onset of Renal Fibrosis. Front. Physiol. 2017, 8, 1026. [Google Scholar] [CrossRef]
- Bigaeva, E.; Stribos, E.G.D.; Mutsaers, H.; Piersma, B.; Leliveld, A.M.; De Jong, I.J.; Bank, R.A.; Seelen, M.A.; Van Goor, H.; Wollin, L.; et al. Inhibition of tyrosine kinase receptor signaling attenuates fibrogenesis in an ex vivo model of human renal fibrosis. Am. J. Physiol.-Ren. Physiol. 2020, 318, F117–F134. [Google Scholar] [CrossRef] [PubMed]
- Bigaeva, E.; Cavanzo, N.P.; Stribos, E.G.D.; De Jong, A.J.; Biel, C.; Mutsaers, H.A.M.; Jensen, M.S.; Nørregaard, R.; Leliveld, A.M.; De Jong, I.J.; et al. Predictive value of precision-cut kidney slices as an ex vivo screening platform for therapeutics in human renal fibrosis. Pharmaceutics 2020, 12, 459. [Google Scholar] [CrossRef] [PubMed]
- Tingskov, S.J.; Jensen, M.S.; Pedersen, C.E.T.; de Araujo, I.B.B.A.; Mutsaers, H.A.M.; Nørregaard, R. Tamoxifen attenuates renal fibrosis in human kidney slices and rats subjected to unilateral ureteral obstruction. Biomed. Pharmacother. 2021, 133, 111003. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, R.H.; Kresse, J.-C.; Farmer, L.K.; Thézénas, M.L.; Kessler, B.M.; Lindeman, J.H.N.; Sharples, E.J.; Welsh, G.I.; Norregaard, R.; Ploeg, R.J.; et al. Cytoskeletal protein degradation in brain death donor kidneys associates with adverse posttransplant outcomes. Am. J. Transplant. 2021, 22, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, L.L.; Leuvenink, H.G.D.; Olinga, P.; Ruigrok, M.J.R. Shifting paradigms for suppressing fibrosis in kidney transplants: Supplementing perfusion solutions with antifibrotic drugs. Front. Med. 2021, 8, 806774. [Google Scholar] [CrossRef]
- Grosse-Siestrup, C.; Unger, V.; Fehrenberg, C.; Baeyer, H.V.; Fischer, A.; Schäper, F.; Groneberg, D.A. A model of isolated autologously hemoperfused porcine slaughterhouse kidneys. Nephron 2002, 92, 414–421. [Google Scholar] [CrossRef]
- Venema, L.; Brat, A.; Moers, C.; ’t Hart, N.; Ploeg, R.; Hannaert, P.; Minor, T.; Leuvenink, H.G.D.; on behalf of the COPE consortium. Effects of Oxygen During Long-term Hypothermic Machine Perfusion in a Porcine Model of Kidney Donation After Circulatory Death. Transplantation 2019, 103, 2057–2064. [Google Scholar] [CrossRef]
- Hendriks, K.D.W.; Brüggenwirth, I.M.A.; Maassen, H.; Gerding, A.; Bakker, B.; Porte, R.J.; Henning, R.H.; Leuvenink, H.G.F. Renal temperature reduction progressively favors mitochondrial ROS production over respiration in hypothermic kidney preservation. J. Transl. Med. 2019, 17, 265. [Google Scholar] [CrossRef]
- Pool, M.B.F.; Hartveld, L.; Leuvenink, H.G.D.; Moers, C. Normothermic machine perfusion of ischaemically damaged porcine kidneys with autologous, allogeneic porcine and human red blood cells. PLoS ONE 2020, 15, e0229566. [Google Scholar] [CrossRef]
- Karuthu, S.; Blumberg, E.A. Common infections in kidney transplant recipients. Clin. J. Am. Soc. Nephrol. 2012, 7, 2058–2070. [Google Scholar] [CrossRef]
- Blum, M.F.; Liu, Q.; Soliman, B.; Dreher, P.; Okamoto, T.; Poggio, E.D.; Goldfarb, D.A.; Baldwin, W.M.; Quintini, C. Comparison of normothermic and hypothermic perfusion in porcine kidneys donated after cardiac death. J. Surg. Res. 2017, 216, 35–45. [Google Scholar] [CrossRef] [PubMed]
- van Furth, L.A.; Venema, L.H.; Hendriks, K.D.W.; Vogelaar, P.C.; Krenning, G.; Leuvenink, H.G.D. The Effects of 6-Chromanol SUL-138 during Hypothermic Machine Perfusion on Porcine Deceased Donor Kidneys. Transplantology 2021, 2, 304–314. [Google Scholar] [CrossRef]
- Golriz, M.; Fonouni, H.; Nickkholgh, A.; Hafezi, M.; Garoussi, C.; Mehrabi, A. Pig kidney transplantation: An up-to-date guideline. Eur. Surg. Res. 2012, 49, 121–129. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Medium Composition |
|---|---|
| WME | Williams Medium E (1X) with GlutaMAX (WME) (Gibco) + 10 µG/mL ciprofloxacin (Fresenius Kabi, France) |
| WME + Glucose | WME + 10 µG/mL ciprofloxacin + extra added D-(+)-glucose solution (Sigma-Aldrich, St. Louis, MO, USA) total end concentration 25 mM |
| WME + Dextran40 | WME + 10 µG/mL ciprofloxacin + 25 µG/mL Amphotericin B + 3.5% Dextran40 (Sigma-Aldrich, St. Louis, USA) |
| WME + Fungi (0.25) | WME + 10 µG/mL ciprofloxacin + 25 µG/mL Amphotericin B (Fungizone) (Merck) |
| WME + Fungi (1) | WME + 10 µG/mL ciprofloxacin + 100 µG/mL Amphotericin B |
| RPMI | Roswell Park Memorial Institute 1640 (RPMI) (Gibco) + 10 µG/mL ciprofloxacin + 11 mM D-(+)-glucose solution |
| Blood | Diluted blood NMP perfusate (See Appendix A) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Furth, L.A.; Leuvenink, H.G.D.; Seras, L.; de Graaf, I.A.M.; Olinga, P.; van Leeuwen, L.L. Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury. Transplantology 2022, 3, 139-151. https://doi.org/10.3390/transplantology3020015
van Furth LA, Leuvenink HGD, Seras L, de Graaf IAM, Olinga P, van Leeuwen LL. Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury. Transplantology. 2022; 3(2):139-151. https://doi.org/10.3390/transplantology3020015
Chicago/Turabian Stylevan Furth, L. Annick, Henri G. D. Leuvenink, Lorina Seras, Inge A. M. de Graaf, Peter Olinga, and L. Leonie van Leeuwen. 2022. "Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury" Transplantology 3, no. 2: 139-151. https://doi.org/10.3390/transplantology3020015
APA Stylevan Furth, L. A., Leuvenink, H. G. D., Seras, L., de Graaf, I. A. M., Olinga, P., & van Leeuwen, L. L. (2022). Exploring Porcine Precision-Cut Kidney Slices as a Model for Transplant-Related Ischemia-Reperfusion Injury. Transplantology, 3(2), 139-151. https://doi.org/10.3390/transplantology3020015