Climate and Local Hydrography Underlie Recent Regime Shifts in Plankton Communities off Galicia (NW Spain)

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Series
2.2. Data Preparation
2.3. Statistical Analysis
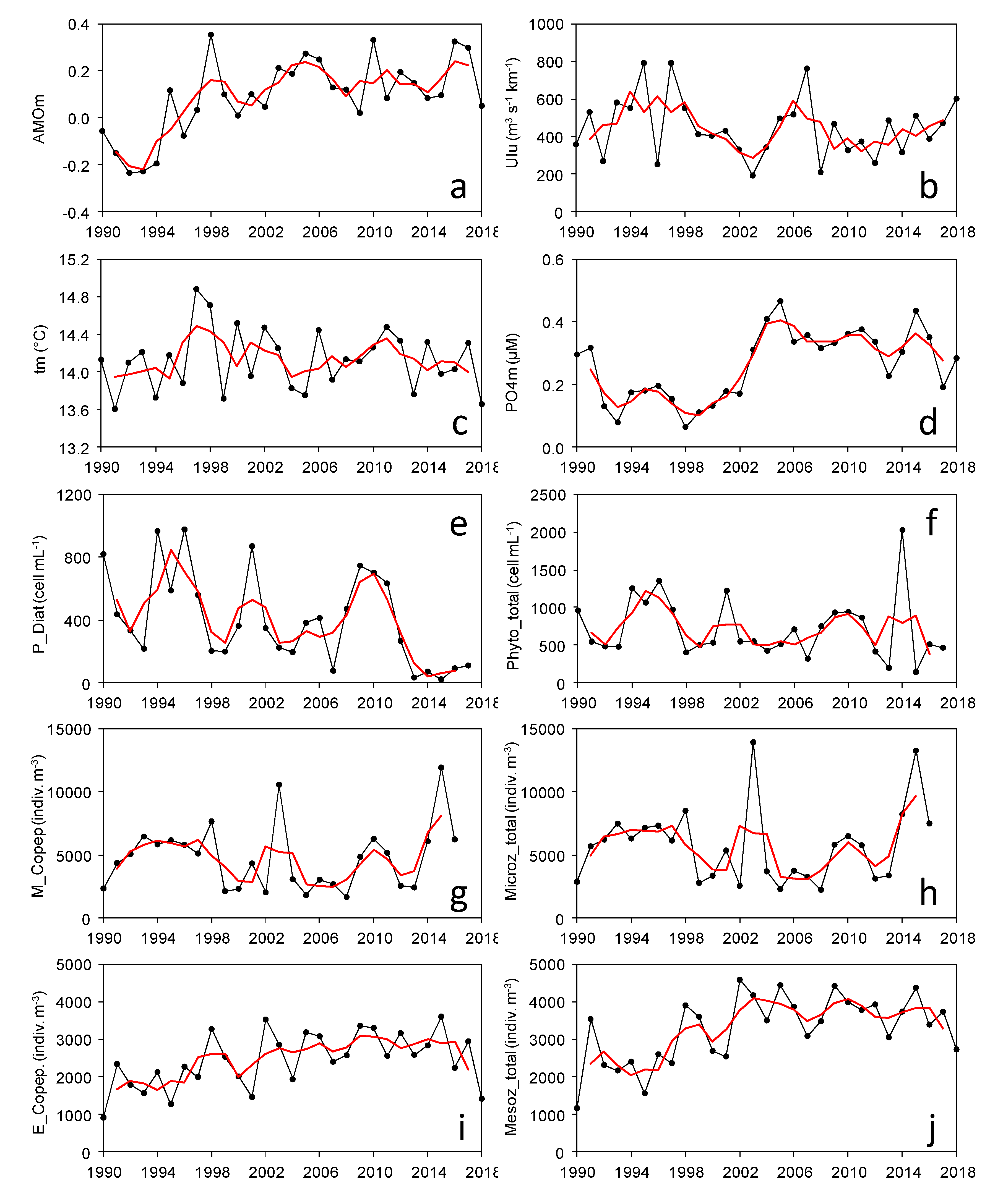
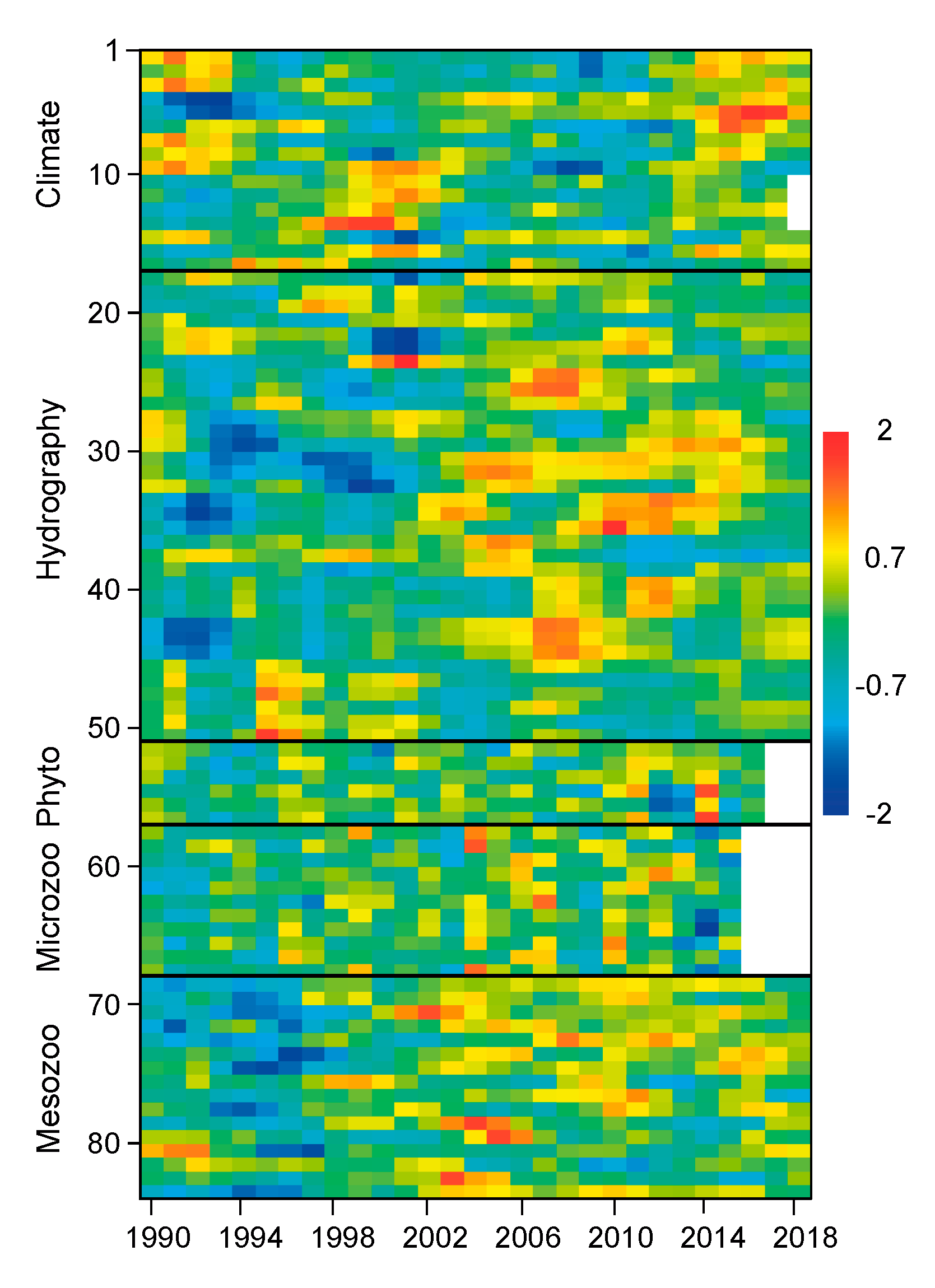
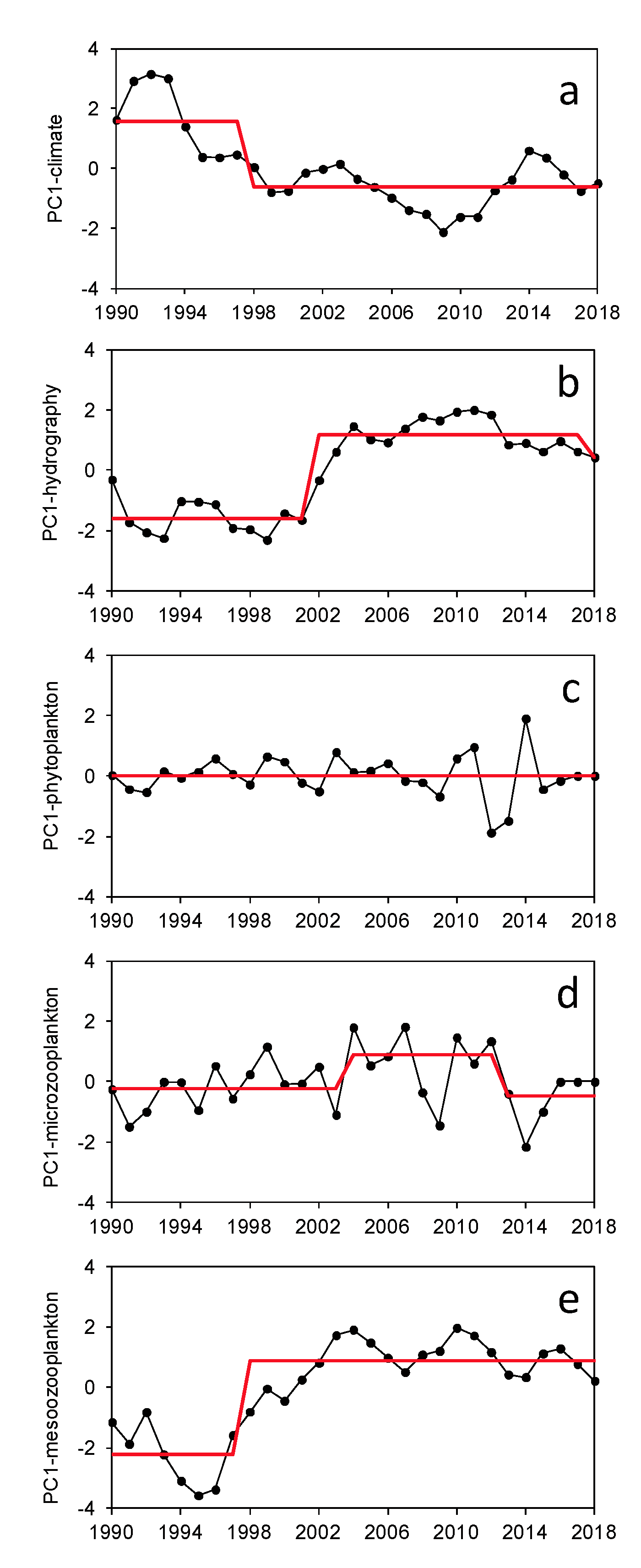
3. Results
3.1. Multiannual Variability of Series
3.2. Regime Shifts
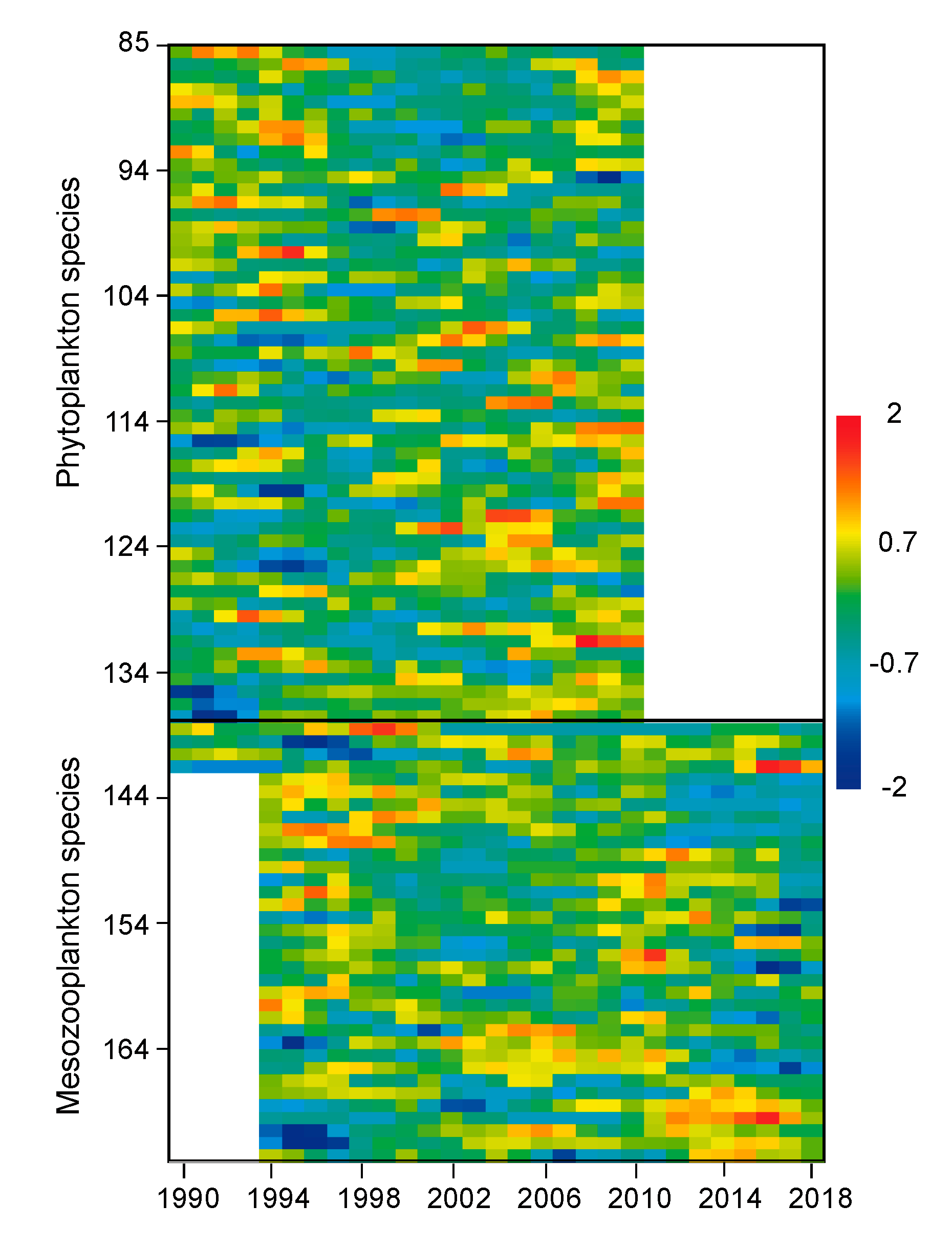
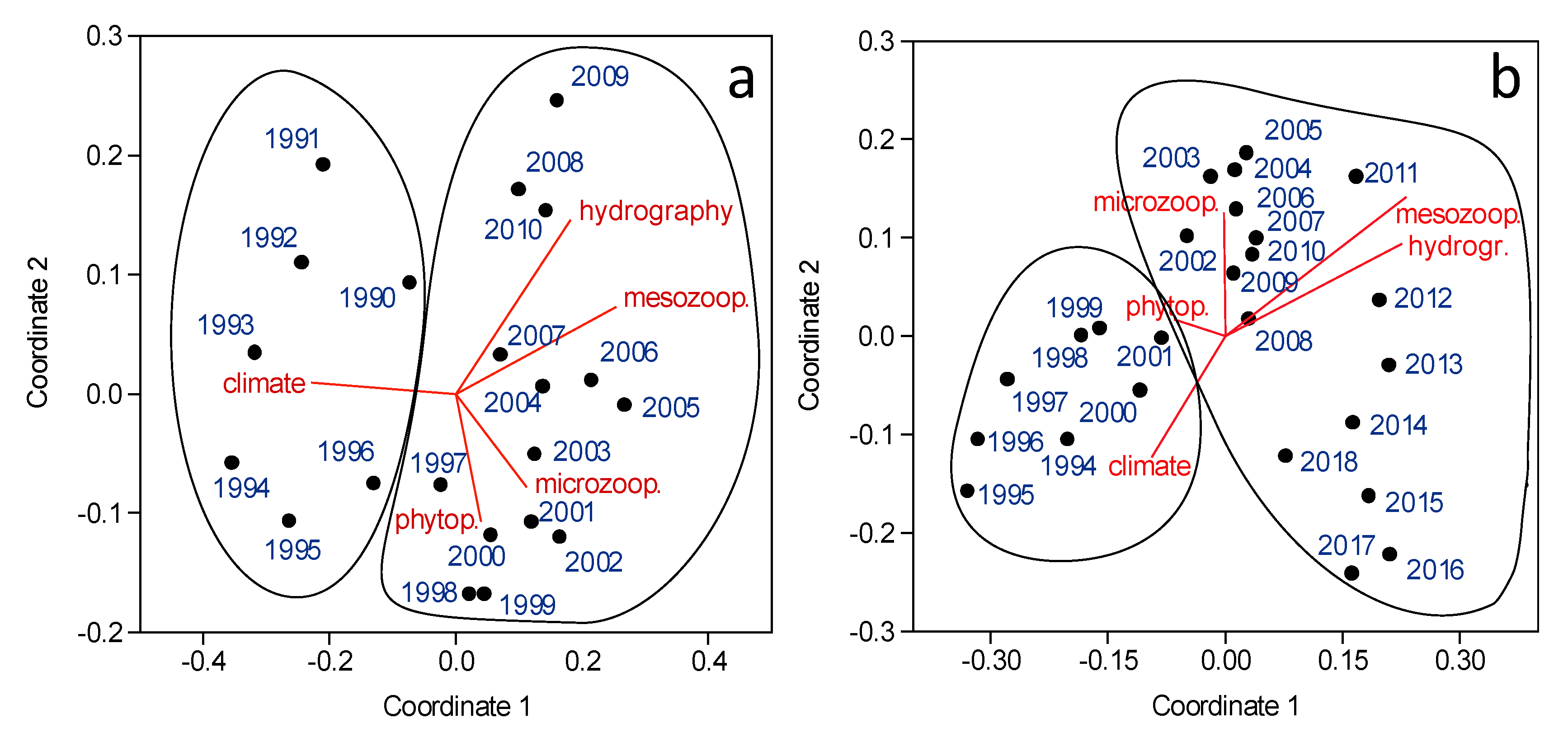
3.3. Regimes in Plankton Species Assemblages
4. Discussion
4.1. Regime Shifts in NW Iberia
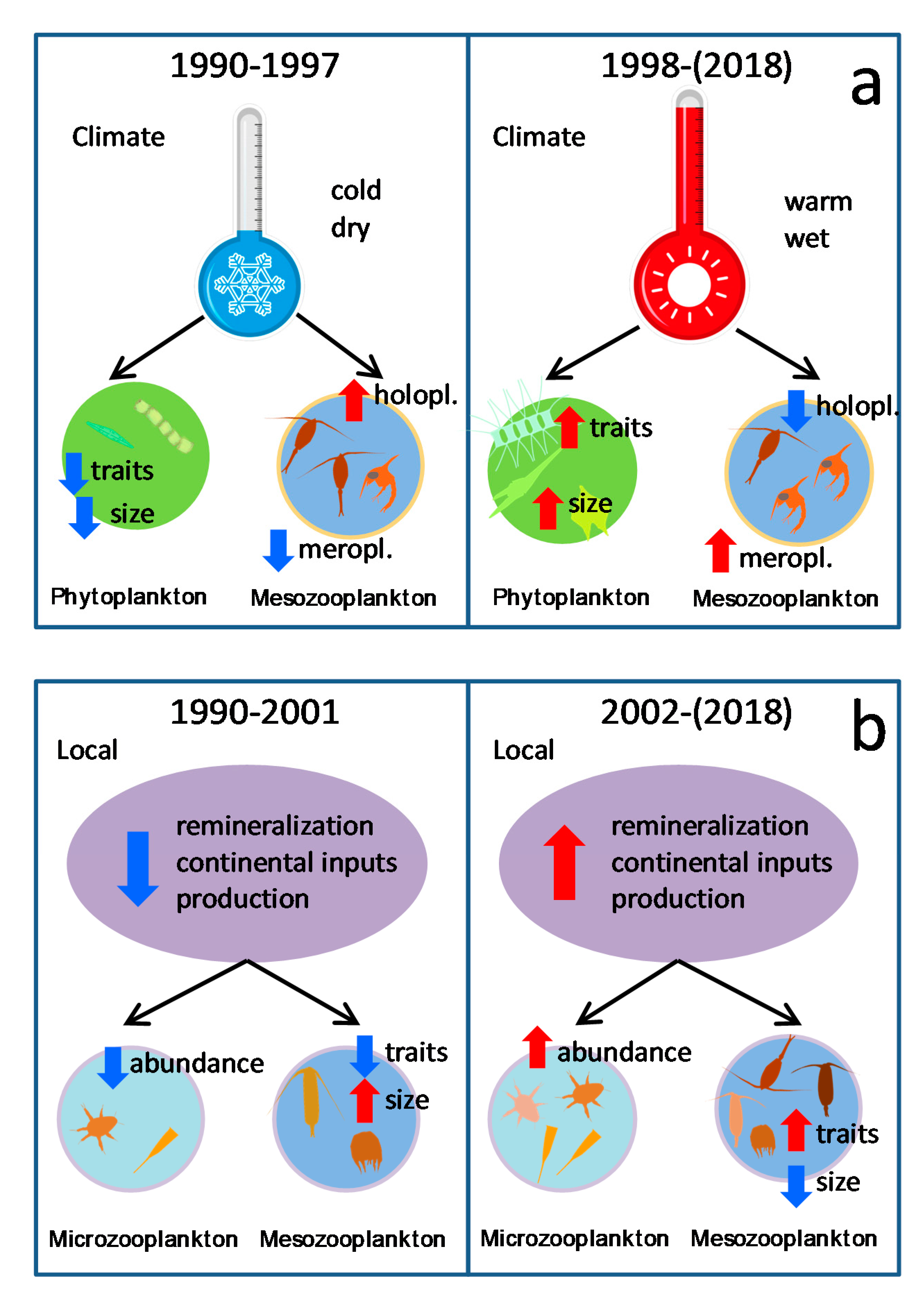
4.2. Shifts Related to Climate
4.3. Shifts Related to Local-Oceanography
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Möllmann, C.; Folke, C.; Edwards, M.; Conversi, A. Marine regime shifts around the globe: Theory, drivers and impacts. Phil. Trans. R. Soc. B 2015, 370. [Google Scholar] [CrossRef]
- Conversi, A.; Dakos, V.; Gardmark, A.; Ling, S.; Folke, C.; Mumby, P.J.; Greene, C.; Edwards, M.; Blenckner, T.; Casini, M.; et al. A holistic view of marine regime shifts. Phil. Trans. R. Soc. B 2015, 370. [Google Scholar] [CrossRef]
- Reid, P.C.; Edwards, M.; Beaugrand, G.; Skogen, M.; Stevens, D. Periodic changes in the zooplankton of the North Sea during the 20th century linked to oceanic inflow. Fish. Oceanogr. 2001, 12, 260–269. [Google Scholar] [CrossRef]
- Beaugrand, G. Long-term changes in copepod abundance and diversity in the North-East Atlantic in relation to fluctuations in the hydroclimatic environment. Fish. Oceanogr. 2003, 12, 270–283. [Google Scholar] [CrossRef]
- Hallett, T.B.; Coulson, T.; Pilkington, J.G.; Clutton-Brock, T.H.; Pemberton, J.M.; Grenfell, B.T. Why large-scale climate indices seem to predict ecological processes better than local weather. Nature 2004, 430, 71–75. [Google Scholar] [CrossRef]
- Molinero, J.C.; Reygondeau, G.; Bonnet, D. Climate variance influence on the non-stationary plankton dynamics. Mar. Environ. Res. 2013, 89, 91–96. [Google Scholar] [CrossRef]
- Alheit, J.; Gröger, J.; Licandro, P.; McQuinn, I.H.; Pohlmann, T.; Tsikliras, A.C. What happened in the mid-1990s? The coupled ocean-atmosphere processes behind climate-induced ecosystem changes in the northeast atlantic and the mediterranean. Deep-Sea Res. II 2019, 159, 130–142. [Google Scholar] [CrossRef]
- Möllmann, C.; Conversi, A.; Edwards, M. Comparative analysis of european wide marine ecosystem shifts: A large-scale approach for developing the basis for ecosystem-based management. Biol. Lett. 2011, 7, 484–486. [Google Scholar] [CrossRef][Green Version]
- Kraberg, A.C.; Wiltshire, K.H. Regime shifts in the marine environment: How do they affect ecosystem services. In The Mediterranean Sea: Its History and Present Challenges; Goffredo, S., Dubinsky, Z., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 2014; pp. 499–504. [Google Scholar]
- Beaugrand, G. The North Sea regime shift: Evidence, causes, mechanisms and consequences. Prog. Oceanogr. 2004, 60, 245–262. [Google Scholar] [CrossRef]
- McQuatters-Gollop, A.; Vermaat, J.E. Covariance among North Sea ecosystem state indicators during the past 50 years—Contrasts between coastal and open waters. J. Sea Res. 2011, 65, 284–292. [Google Scholar] [CrossRef]
- Peperzak, L.; Witte, H. Abiotic drivers of interannual phytoplankton variability and a 1999–2000 regime shift in the North Sea examined by multivariate statistics. J. Phycol. 2019, 55, 1274–1289. [Google Scholar] [CrossRef] [PubMed]
- Cabrero, Á.; González-Nuevo, G.; Gago, J.; Cabanas, J.M. Study of sardine (Sardina pilchardus) regime shifts in the Iberian Atlantic shelf waters. Fish. Oceanogr. 2019, 28, 305–316. [Google Scholar] [CrossRef]
- Hemery, G.; D’Amico, F.; Castege, I.; Dupont, B.; D’Elbee, J.; Lalanne, Y.; Mouches, C. Detecting the impact of oceano-climatic changes on marine ecosystems using a multivariate index: The case of the Bay of Biscay (North Atlantic-European ocean). Glob. Chang. Biol. 2008, 14, 27–38. [Google Scholar] [CrossRef]
- Luczak, C.; Beaugrand, G.; Jaffré, M.; Lenoir, S. Climate change impact on Balearic shearwater through a trophic cascade. Biol. Lett. 2011, 7, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Pershing, A.J.; Mills, K.E.; Record, N.R.; Stamieszkin, K.; Wurtzell, K.V.; Byron, C.J.; Fitzpatrick, D.; Golet, W.J.; Koob, E. Evaluating trophic cascades as drivers of regime shifts in different ocean ecosystems. Phil. Trans. R. Soc. B 2015, 370. [Google Scholar] [CrossRef]
- Kröncke, I.; Neumann, H.; Dippner, J.W.; Holbrook, S.; Lamy, T.; Miller, R.; Padedda, B.M.; Pulina, S.; Reed, D.C.; Reinikainen, M.; et al. Comparison of biological and ecological long-term trends related to northern hemisphere climate in different marine ecosystems. Nat. Conserv. 2019, 34, 311–341. [Google Scholar] [CrossRef]
- Hernández-Fariñas, T.; Soudant, D.; Barillé, L.; Belin, C.; Lefebvre, A.; Bacher, C. Temporal changes in the phytoplankton community along the French coast of the Eastern English Channel and the Southern Bight of the North Sea. ICES J. Mar. Sci. 2014, 71, 821–833. [Google Scholar] [CrossRef]
- Reygondeau, G.; Molinero, J.C.; Coombs, S.; MacKenzie, B.R.; Bonnet, D. Progressive changes in the western English Channel foster a reorganization in the plankton food web. Prog. Oceanogr. 2015, 137, 524–532. [Google Scholar] [CrossRef]
- Bode, A.; Alvarez-Ossorio, M.T.; Miranda, A.; Ruiz Villarreal, M. Shifts between gelatinous and crustacean plankton in a coastal upwelling region. ICES J. Mar. Sci. 2013, 70, 934–942. [Google Scholar] [CrossRef]
- Buttay, L.; Miranda, A.; Casas, G.; González-Quirós, R.; Nogueira, E. Long-term and seasonal zooplankton dynamics in the northwest Iberian shelf and its relationship with meteo-climatic and hydrographic variability. J. Plankton Res. 2016, 38, 106–121. [Google Scholar] [CrossRef]
- Valdés, L.; López-Urrutia, A.; Cabal, J.; Alvarez-Ossorio, M.; Bode, A.; Miranda, A.; Cabanas, M.; Huskin, I.; Anadón, R.; Alvarez-Marqués, F.; et al. A decade of sampling in the Bay of Biscay: What are the zooplankton time series telling us? Prog. Oceanogr. 2007, 74, 98–114. [Google Scholar] [CrossRef]
- Bode, A.; Alvarez-Ossorio, M.T.; Cabanas, J.M.; Miranda, A.; Varela, M. Recent trends in plankton and upwelling intensity off Galicia (NW Spain). Prog. Oceanogr. 2009, 83, 342–350. [Google Scholar] [CrossRef]
- Bode, A.; Anadón, R.; Morán, X.A.G.; Nogueira, E.; Teira, E.; Varela, M. Decadal variability in chlorophyll and primary production off NW Spain. Clim. Res. 2011, 48, 293–305. [Google Scholar] [CrossRef]
- Pérez, F.F.; Padin, X.A.; Pazos, Y.; Gilcoto, M.; Cabanas, M.; Pardo, P.C.; Doval, M.D.; Farina-Bustos, L. Plankton response to weakening of the Iberian coastal upwelling. Glob. Chang. Biol. 2010, 16, 1258–1267. [Google Scholar] [CrossRef]
- González-Gil, R.; González Taboada, F.; Höffer, J.; Anadón, R. Winter mixing and coastal upwelling drive long-term changes in zooplankton in the Bay of Biscay (1993-2010). J. Plankton Res. 2015, 37, 337–351. [Google Scholar] [CrossRef]
- Bode, A.; Álvarez, M.; Ruíz-Villarreal, M.; Varela, M.M. Changes in phytoplankton production and upwelling intensity off A Coruña (NW Spain) for the last 28 years. Ocean Dyn. 2019. [Google Scholar] [CrossRef]
- Alvarez-Salgado, X.A.; Beloso, S.; Joint, I.; Nogueira, E.; Chou, L.; Pérez, F.F.; Groom, S.; Cabanas, J.M.; Rees, A.P.; Elskens, M. New production of the NW Iberian shelf during the upwelling season over the period 1982–1999. Deep Sea Res. 2002, 49, 1725–1739. [Google Scholar] [CrossRef]
- Beca-Carretero, P.P.; Otero, J.; Land, P.E.; Groom, S.; Alvarez-Salgado, X.A. Seasonal and inter-annual variability of net primary production in the NW Iberian margin (1998–2016) in relation to wind stress and sea surface temperature. Prog. Oceanogr. 2019, 178. [Google Scholar] [CrossRef]
- Casabella, N.; Lorenzo, M.N.; Taboada, J.J. Trends of the Galician upwelling in the context of climate change. J. Sea Res. 2014, 93, 23–27. [Google Scholar] [CrossRef]
- Available online: http://www.seriestemporales-ieo.net (accessed on 24 September 2020).
- Available online: https://psl.noaa.gov/gcos_wgsp/Timeseries/ (accessed on 30 March 2020).
- Available online: https://crudata.uea.ac.uk/cru/data/nao/ (accessed on 25 March 2020).
- Edwards, M.; Beaugrand, G.; Helaouet, P.; Alheit, J.; Coombs, S. Marine ecosystem response to the Atlantic multidecadal oscillation. PLoS ONE 2013, 8, e57212. [Google Scholar] [CrossRef]
- Available online: https://opendata.aemet.es/ (accessed on 15 March 2020).
- González-Nuevo, G.; Gago, J.; Cabanas, J.M. Upwelling index: A powerful tool for marine research in the nw iberian upwelling system. J. Oper. Oceanogr. 2014, 7, 45–55. [Google Scholar] [CrossRef]
- Available online: http://www.indicedeafloramiento.ieo.es/ (accessed on 15 March 2020).
- Available online: https://doi.org/10.1594/PANGAEA.885413 (accessed on 24 September 2020).
- Available online: https://doi.pangaea.de/10.1594/PANGAEA.919082 (accessed on 24 September 2020).
- Bode, A.; Estévez, M.G.; Varela, M.; Vilar, J.A. Annual trend patterns of phytoplankton species abundance belie homogeneous taxonomical group responses to climate in the NE Atlantic upwelling. Mar. Environ. Res. 2015, 110, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://doi.org/10.1594/PANGAEA.908815 (accessed on 24 September 2020).
- Available online: https://doi.pangaea.de/10.1594/PANGAEA.919081 (accessed on 24 September 2020).
- Available online: https://doi.pangaea.de/10.1594/PANGAEA.919080 (accessed on 24 September 2020).
- Nogueira, E.; Perez, F.F.; Rios, A.F. Seasonal patterns and long-term trends in an estuarine upwelling ecosystem (Ria de Vigo, NW Spain). Estuar. Coast. Shelf Sci. 1997, 44, 285–300. [Google Scholar] [CrossRef]
- Rodionov, S.N. A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006; 192p. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Folland, C.K.; Knight, J.; Linderholm, H.; Fereday, D.; Ineson, S.; Hurrell, J.W. The summer North Atlantic Oscillation: Past, present and future. J. Clim. 2009, 22, 1082–1103. [Google Scholar] [CrossRef]
- López-Parages, J.; Rodríguez-Fonseca, B.; Dommenget, D.; Frauen, C. ENSO influence on the North Atlantic European climate: A non-linear and non-stationary approach. Clim. Dyn. 2016, 47, 2071–2084. [Google Scholar] [CrossRef]
- O’Reilly, C.H.; Woollings, T.; Zanna, L. The dynamical influence of the atlantic multidecadal oscillation on continental climate. J. Clim. 2017, 30, 7213–7230. [Google Scholar] [CrossRef]
- Knudsen, M.F.; Seidenkrantz, M.-S.; Jacobsen, B.H.; Kuijpers, A. Tracking the atlantic multidecadal oscillation through the last 8000 years. Nat. Commun. 2011, 2. [Google Scholar] [CrossRef]
- Marshall, J.; Kushnir, Y.; Battisti, D.; Chang, P.; Czaja, A.; Dickson, R.; Hurrell, J.; McCartney, M.; Saravanan, R.; Visbeck, M. North Atlantic climate variability: Phenomena, impacts and mechanisms. Int. J. Climatol. 2001, 21, 1863–1898. [Google Scholar] [CrossRef]
- Prieto, E.; Gonzalez-Pola, C.; Lavin, A.; Holliday, N.P. Interannual variability of the Northwestern Iberia deep ocean: Response to large-scale North Atlantic forcing. J. Geophys. Res. Oceans 2015, 120, 832–847. [Google Scholar] [CrossRef]
- Rios, A.F.; Pérez, F.F.; Fraga, F. Water masses in the upper and middle North Atlantic Ocean East of Azores. Deep Sea Res. 1992, 39, 645–658. [Google Scholar] [CrossRef]
- Pérez, F.F.; Mouriño, C.; Fraga, F.; Rios, A.F. Displacement of water masses and remineralization rates off the Iberian peninsula by nutrient anomalies. J. Mar. Res. 1993, 51, 869–892. [Google Scholar] [CrossRef][Green Version]
- Otero, P.; Ruiz-Villarreal, M.; Peliz, A. River plume fronts off NW Iberia from satellite observations and model data. ICES J. Mar. Sci. 2009, 66, 1853–1864. [Google Scholar] [CrossRef]
- deYoung, B.; Jarre, A. Regime shifts: Methods of analysis. In Encyclopedia of Ocean Sciences; Cochran, J.K., Bokuniewicz, H., Yager, P., Eds.; Academic Press: London, UK, 2019; pp. 489–492. [Google Scholar]
- Rodionov, S.N. Use of prewhitening in climate regime shift detection. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Liu, Q.; Wan, S.; Gu, B. A review of the detection methods for climate regime shifts. Discrete Dyn. Nat. Soc. 2016, 2016. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A. Trait-based community ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 615–639. [Google Scholar] [CrossRef]
- Otero, J.; Bode, A.; Álvarez-Salgado, X.A.; Varela, M. Role of functional traits variability in the response of individual phytoplankton species to changing environmental conditions in a coastal upwelling zone. Mar. Ecol. Prog. Ser. 2018, 596, 33–47. [Google Scholar] [CrossRef]
- Freund, J.A.; Grüner, N.; Brüse, S.; Wiltshire, K.H. Changes in the phytoplankton community at Helgoland, North Sea: Lessons from single spot time series analyses. Mar. Biol. 2012, 159, 2561–2571. [Google Scholar] [CrossRef]
- Alvarez-Fernandez, S.; Lindeboom, H.; Meesters, E. Temporal changes in plankton of the North Sea: Community shifts and environmental drivers. Mar. Ecol. Prog. Ser. 2012, 462, 21–38. [Google Scholar] [CrossRef]
- Boersma, M.; Wiltshire, K.H.; Kong, S.M.; Greve, W.; Renz, J. Long-term change in the copepod community in the Southern German Bight. J. Sea Res. 2015, 101, 41–50. [Google Scholar] [CrossRef]
- Pomerleau, C.; Sastri, A.R.; Beisner, B.E. Evaluation of functional trait diversity for marine zooplankton communities in the Northeast subarctic Pacific ocean. J. Plankton Res. 2015, 37, 712–726. [Google Scholar] [CrossRef]
- Dippner, J.W.; Kornilovs, G.; Junker, K. A multivariate Baltic Sea environmental index. Ambio 2012, 41, 699–708. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chust, G.; Allen, J.I.; Bopp, L.; Schrum, C.; Holt, J.; Tsiaras, K.; Zavatarelli, M.; Chifflet, M.; Cannaby, H.; Dadou, I.; et al. Biomass changes and trophic amplification of plankton in a warmer ocean. Glob. Chang. Biol. 2014, 20, 2124–2139. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1990–1997 | 2000–2010 | ||||||
|---|---|---|---|---|---|---|---|
| Series No. | Series | Taxonomic Group | Mean Abund. | Mean Abund. | D2 | D2/sd | Contr. % |
| 116 | Navicula transitans | Diatoms | 0.33 | 0.82 | 2.64 | 1.61 | 5.30 |
| 126 | Pseudo-nitzschia delicatissima | Diatoms | 3.00 | 12.93 | 2.14 | 1.16 | 4.31 |
| 108 | Chaetoceros spp. | Diatoms | 6.15 | 30.46 | 2.09 | 1.16 | 4.21 |
| 136 | Torodinium robustum | Dinoflagellates | 0.20 | 0.50 | 1.88 | 0.93 | 3.78 |
| 138 | Heterocapsa niei | Dinoflagellates | 4.05 | 8.26 | 1.71 | 0.90 | 3.43 |
| 97 | Lauderia annulata | Diatoms | 4.11 | 1.08 | 1.60 | 1.20 | 3.21 |
| 101 | Skeletonema costatum | Diatoms | 42.56 | 3.21 | 1.57 | 1.04 | 3.16 |
| 120 | Nitzschia spp. | Diatoms | 0.77 | 1.08 | 1.52 | 0.93 | 3.06 |
| 110 | Dictyocha fibula | Dictyochophyceae | 0.05 | 0.16 | 1.29 | 1.02 | 2.58 |
| 85 | Bacteriastrum delicatulum | Diatoms | 1.11 | 0.04 | 1.22 | 1.05 | 2.45 |
| 106 | Chaetoceros decipiens | Diatoms | 1.15 | 0.29 | 1.18 | 1.04 | 2.38 |
| 123 | Prorocentrum balticum | Dinoflagellates | 1.03 | 4.47 | 1.16 | 0.85 | 2.32 |
| 132 | Thalassiosira levanderi | Diatoms | 0.49 | 20.64 | 1.15 | 0.68 | 2.31 |
| 137 | Gyrodinium spirale | Dinoflagellates | 0.23 | 0.39 | 1.12 | 0.94 | 2.25 |
| 122 | Proboscia alata | Diatoms | 0.87 | 2.05 | 1.11 | 0.73 | 2.22 |
| 92 | Chaetoceros socialis | Diatoms | 252.08 | 131.83 | 1.07 | 0.80 | 2.16 |
| 91 | Chaetoceros gracilis | Diatoms | 3.56 | 2.30 | 1.05 | 0.84 | 2.11 |
| 1994–2001 | 2002–2018 | ||||||
|---|---|---|---|---|---|---|---|
| Series No. | Series | Taxonomic Group | Mean Abund. | Mean Abund. | D2 | D2/sd | Contr. % |
| 173 | Oithona similis | Copepoda | 34.12 | 133.02 | 2.80 | 1.04 | 7.63 |
| 172 | Oithona nana | Copepoda | 8.48 | 27.82 | 2.27 | 0.92 | 6.18 |
| 139 | Penilia avirostris | Cladocera | 0.44 | 0.02 | 1.87 | 1.28 | 5.10 |
| 140 | Podon intermedius | Cladocera | 66.51 | 126.59 | 1.72 | 0.86 | 4.68 |
| 141 | Evadne nordmani | Cladocera | 25.89 | 83.63 | 1.43 | 1.01 | 3.89 |
| 144 | Ctenocalanus vanus | Copepoda | 28.37 | 14.54 | 1.39 | 1.21 | 3.78 |
| 148 | Temora stylifera | Copepoda | 18.45 | 4.75 | 1.37 | 1.08 | 3.74 |
| 164 | Oncaea media | Copepoda | 238.55 | 438.45 | 1.33 | 0.75 | 3.62 |
| 168 | Pseudocalanus elongatus | Copepoda | 134.46 | 152.58 | 1.29 | 1.09 | 3.50 |
| 170 | Centropages typicus | Copepoda | 2.87 | 9.23 | 1.26 | 1.20 | 3.42 |
| 171 | Corycaeus spp. | Copepoda | 0.12 | 1.17 | 1.24 | 0.84 | 3.39 |
| 147 | Temora longicornis | Copepoda | 74.14 | 47.80 | 1.21 | 0.95 | 3.29 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bode, A.; Álvarez, M.; García García, L.M.; Louro, M.Á.; Nieto-Cid, M.; Ruíz-Villarreal, M.; Varela, M.M. Climate and Local Hydrography Underlie Recent Regime Shifts in Plankton Communities off Galicia (NW Spain). Oceans 2020, 1, 181-197. https://doi.org/10.3390/oceans1040014
Bode A, Álvarez M, García García LM, Louro MÁ, Nieto-Cid M, Ruíz-Villarreal M, Varela MM. Climate and Local Hydrography Underlie Recent Regime Shifts in Plankton Communities off Galicia (NW Spain). Oceans. 2020; 1(4):181-197. https://doi.org/10.3390/oceans1040014
Chicago/Turabian StyleBode, Antonio, Marta Álvarez, Luz María García García, Maria Ángeles Louro, Mar Nieto-Cid, Manuel Ruíz-Villarreal, and Marta M. Varela. 2020. "Climate and Local Hydrography Underlie Recent Regime Shifts in Plankton Communities off Galicia (NW Spain)" Oceans 1, no. 4: 181-197. https://doi.org/10.3390/oceans1040014
APA StyleBode, A., Álvarez, M., García García, L. M., Louro, M. Á., Nieto-Cid, M., Ruíz-Villarreal, M., & Varela, M. M. (2020). Climate and Local Hydrography Underlie Recent Regime Shifts in Plankton Communities off Galicia (NW Spain). Oceans, 1(4), 181-197. https://doi.org/10.3390/oceans1040014