
Variation in Dairy Milk Composition and Properties Has Little Impact on Cheese Ripening: Insights from a Traditional Swedish Long-Ripening Cheese

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Raw Milk Gross Composition, SCC, and Bacteria
2.3. Casein Micelle Size
2.4. Rennet-Induced Coagulation
2.5. Plasmin Activity
2.6. Total Proteolysis
2.7. Milk Fat Composition
2.8. Cheese Production
2.9. Cheese Final Sensory Evaluation and Ripening Time
2.10. Statistical Design
3. Results and Discussion
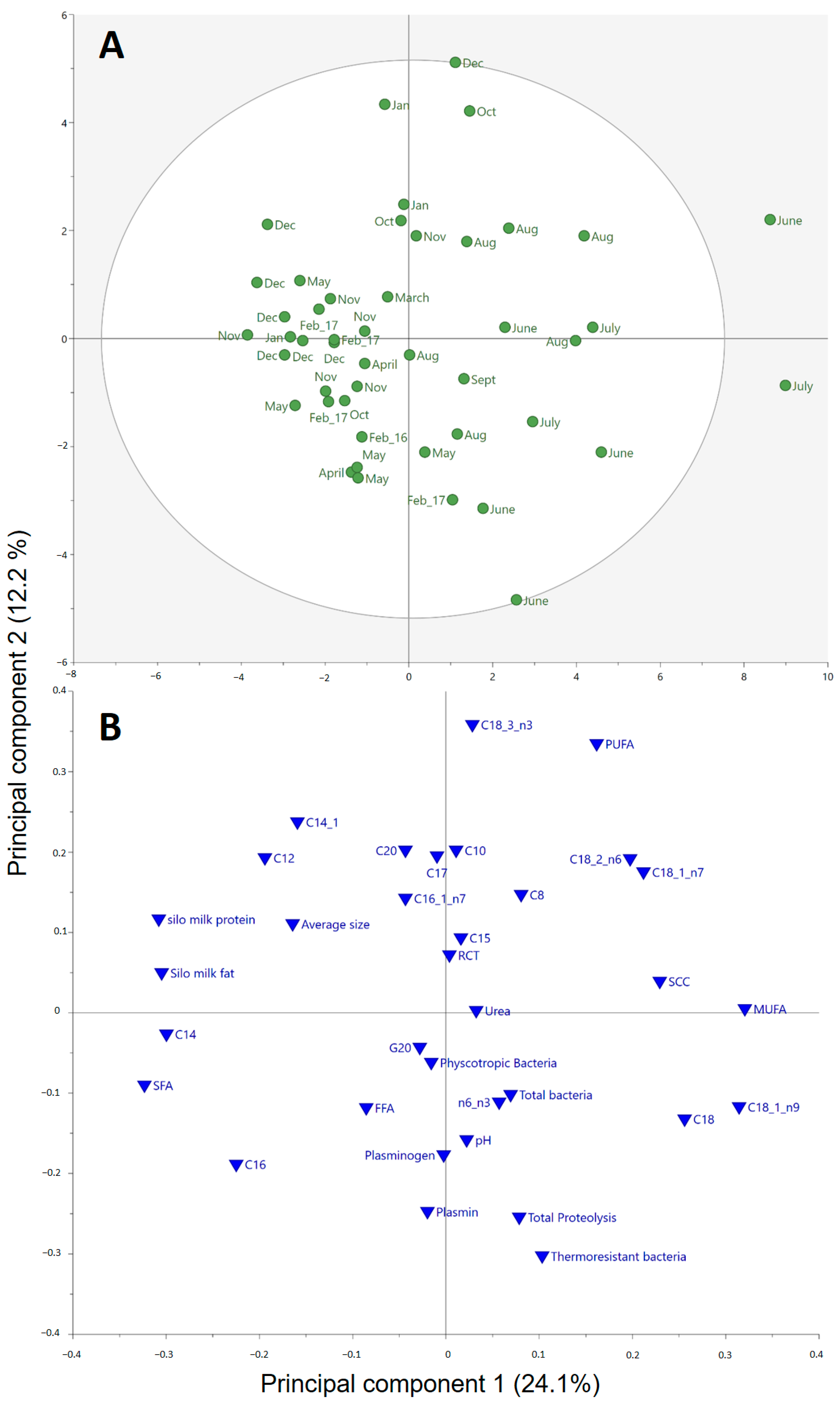
3.1. Variation in Raw Milk Composition and Properties
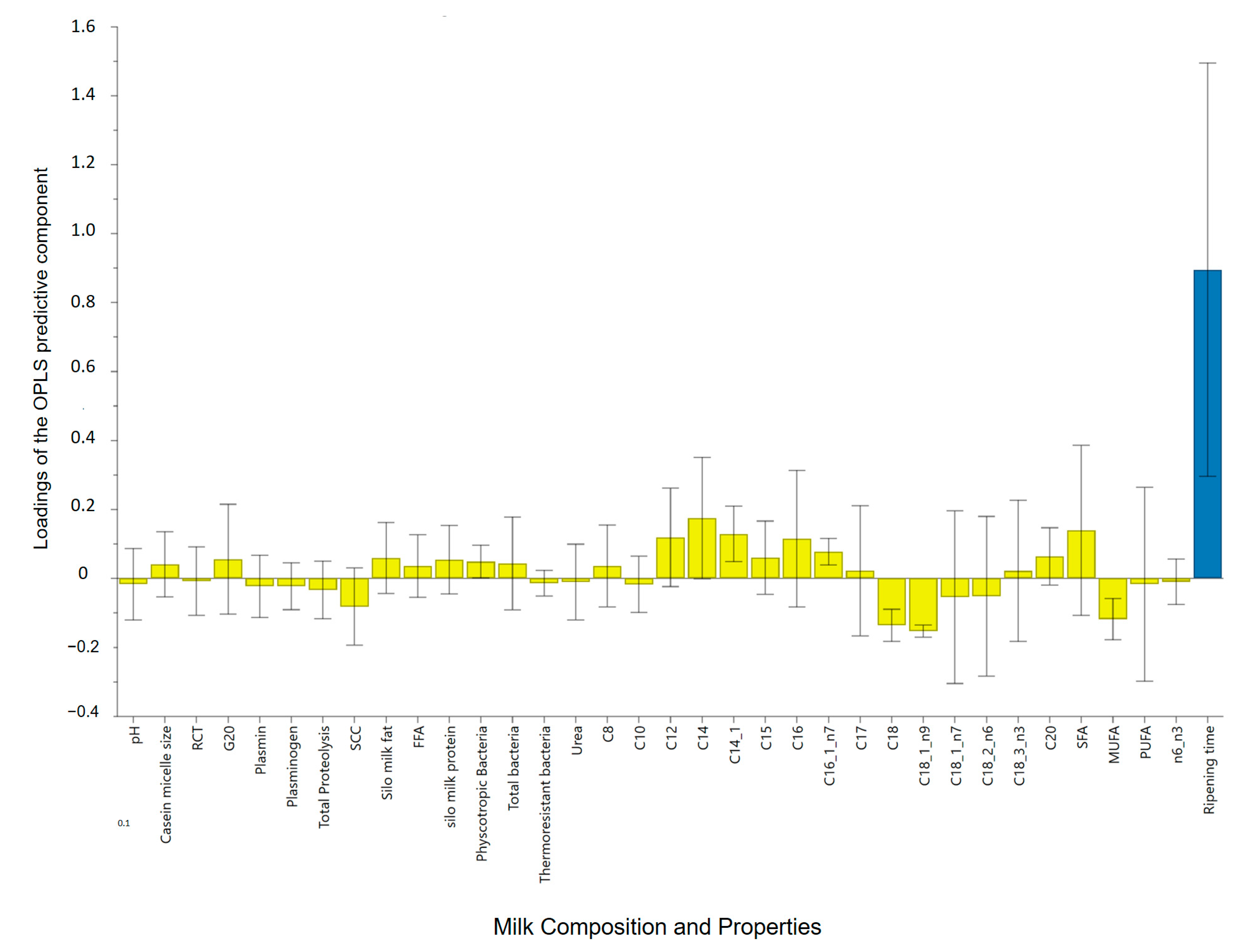
3.2. Cheese Ripening Time
3.3. Ripening Time and Milk Quality
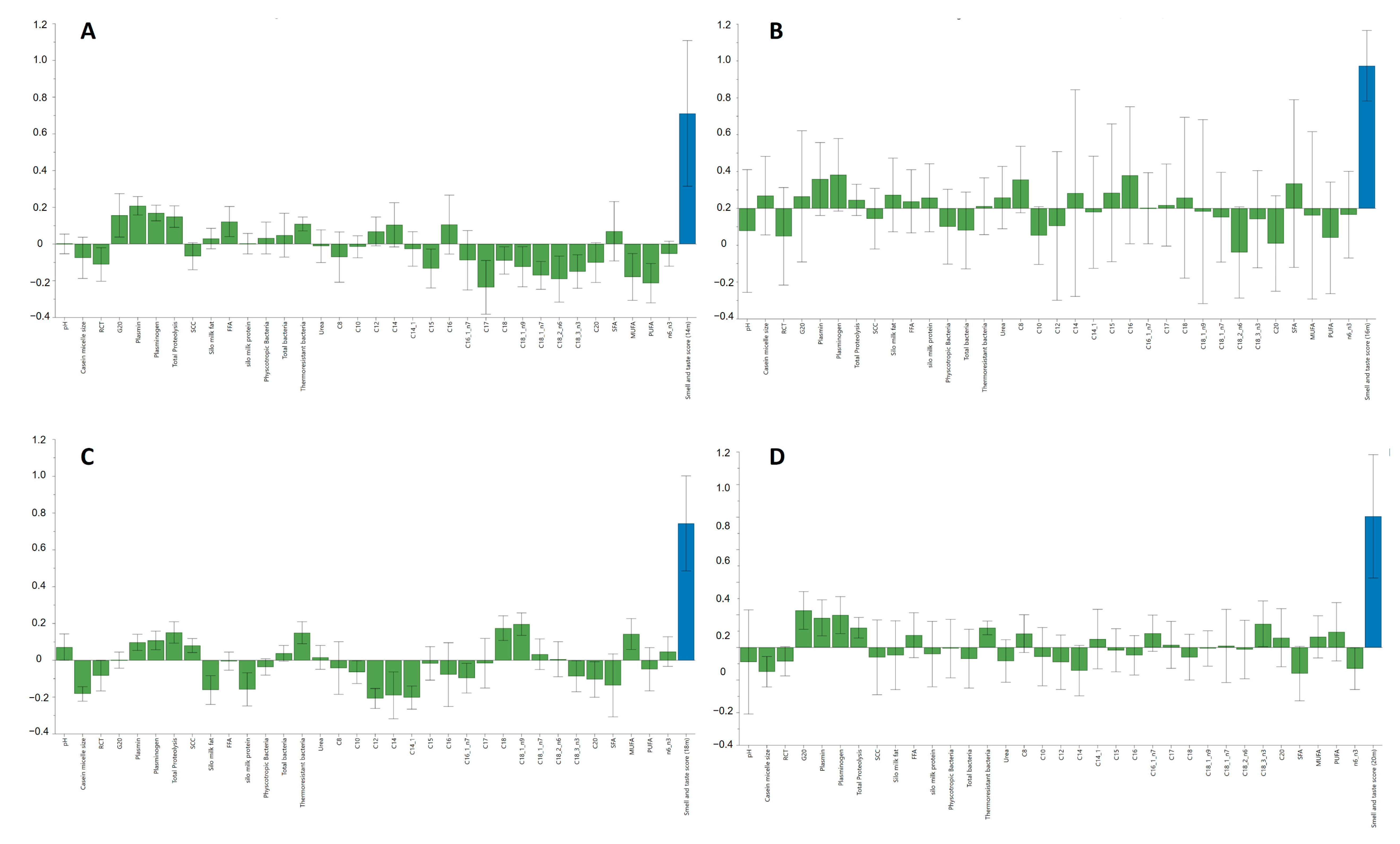
3.4. Effect of Raw Milk Quality Attributes on “Smell and Taste” Sensory Score Values during Ripening
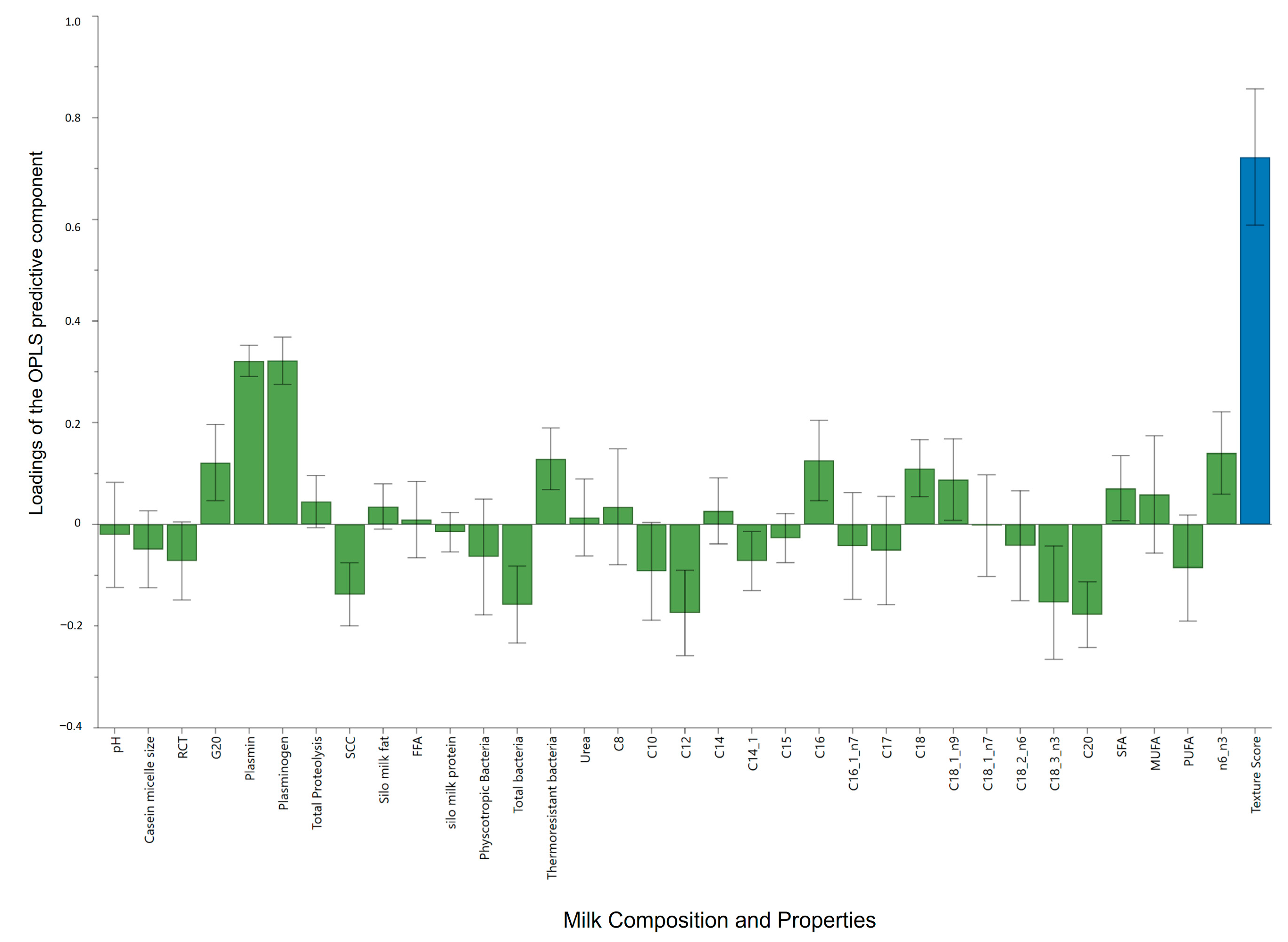
3.5. Effect of Milk Quality Attributes on Texture Scores of Cheeses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heck, J.M.L.; van Valenberg, H.J.F.; Dijkstra, J.; van Hooijdonk, A.C.M. Seasonal Variation in the Dutch Bovine Raw Milk Composition. J. Dairy Sci. 2009, 92, 4745–4755. [Google Scholar] [CrossRef]
- O’Brien, B.; Guinee, T.P. Milk|Seasonal Effects on Processing Properties of Cows’ Milk. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 598–606. ISBN 978-0-12-374407-4. [Google Scholar]
- Williams, R.P.W. The Relationship between the Composition of Milk and the Properties of Bulk Milk Products. Aust. J. Dairy Technol. 2002, 57, 30–44. [Google Scholar]
- Larsen, M.K.; Nielsen, J.H.; Butler, G.; Leifert, C.; Slots, T.; Kristiansen, G.H.; Gustafsson, A.H. Milk Quality as Affected by Feeding Regimens in a Country with Climatic Variation. J. Dairy Sci. 2010, 93, 2863–2873. [Google Scholar] [CrossRef]
- Holt, C.; Muir, D.D. Natural Variations in the Average Size of Bovine Casein Micelles II. Milk Samples from Creamery Bulk Silos in South West Scotland. J. Dairy Res. 1978, 45, 347–353. [Google Scholar] [CrossRef]
- Chen, B.; Lewis, M.J.; Grandison, A.S. Effect of Seasonal Variation on the Composition and Properties of Raw Milk Destined for Processing in the UK. Food Chem. 2014, 158, 216–223. [Google Scholar] [CrossRef]
- Walker, G.P.; Wijesundera, C.; Dunshea, F.R.; Doyle, P.T.; Walker, G.P.; Wijesundera, C.; Dunshea, F.R.; Doyle, P.T. Seasonal and Stage of Lactation Effects on Milk Fat Composition in Northern Victoria. Anim. Prod. Sci. 2013, 53, 560–572. [Google Scholar] [CrossRef]
- Collomb, M.; Bisig, W.; Bütikofer, U.; Sieber, R.; Bregy, M.; Etter, L. Seasonal Variation in the Fatty Acid Composition of Milk Supplied to Dairies in the Mountain Regions of Switzerland. Dairy Sci. Technol. 2008, 88, 631–647. [Google Scholar] [CrossRef]
- Frössling, J.; Ohlson, A.; Hallén-Sandgren, C. Incidence and Duration of Increased Somatic Cell Count in Swedish Dairy Cows and Associations with Milking System Type. J. Dairy Sci. 2017, 100, 7368–7378. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ye, A.; Singh, H. Seasonal Variations in Composition, Properties, and Heat-Induced Changes in Bovine Milk in a Seasonal Calving System. J. Dairy Sci. 2019, 102, 7747–7759. [Google Scholar] [CrossRef]
- Lin, Y.; O’Mahony, J.A.; Kelly, A.L.; Guinee, T.P. Seasonal Variation in the Composition and Processing Characteristics of Herd Milk with Varying Proportions of Milk from Spring-Calving and Autumn-Calving Cows. J. Dairy Res. 2017, 84, 444–452. [Google Scholar] [CrossRef]
- Priyashantha, H.; Lundh, Å.; Höjer, A.; Bernes, G.; Nilsson, D.; Hetta, M.; Saedén, K.H.; Gustafsson, A.H.; Johansson, M. Composition and Properties of Bovine Milk: A Case Study from Dairy Farms in Northern Sweden; Part II. Effect of Monthly Variation. J. Dairy Sci. 2021, 104. [Google Scholar] [CrossRef]
- Delahunty, C.M.; Drake, M.A. Sensory Character of Cheese and its Evaluation. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; General Aspects; Academic Press: Cambridge, MA, USA, 2004; Volume 1, pp. 455–487. [Google Scholar]
- Priyashantha, H.; Höjer, A.; Saedén, K.H.; Lundh, Å.; Johansson, M.; Bernes, G.; Geladi, P.; Hetta, M. Use of Near-Infrared Hyperspectral (NIR-HS) Imaging to Visualize and Model the Maturity of Long-Ripening Hard Cheeses. J. Food Eng. 2020, 264, 109687. [Google Scholar] [CrossRef]
- Priyashantha, H.; Lundh, Å.; Höjer, A.; Bernes, G.; Nilsson, D.; Hetta, M.; Saedén, K.H.; Gustafsson, A.H.; Johansson, M. Composition and Properties of Bovine Milk: A Case Study from Dairy Farms in Northern Sweden; Part I. Effect of Dairy Farming System. J. Dairy Sci. 2021, 104. [Google Scholar] [CrossRef]
- Clay, N.; Garnett, T.; Lorimer, J. Dairy Intensification: Drivers, Impacts and Alternatives. Ambio 2020, 49, 35–48. [Google Scholar] [CrossRef]
- Wehr, H.M.; Frank, J.F. Standard Methods for the Examination of Dairy Products; American Public Health Association: Washington, DC, USA, 2004; ISBN 978-0-87553-002-4. [Google Scholar]
- NordVal International-NMKL 86 Aerobic Microorganisms. Determination in Foods at 37 °C, 30 °C, 25 °C, 20 °C, 17/7 °C or 6.5 °C by the Colony Count Method. 2013. Available online: https://www.nmkl.org/index.php/en/publications/item/aerobe-mikroorgansimer-bestemmelse-i-levnedsmidler-ved-37-c-30-c-25-c-20-c-17-7-c-eller-65-c-efter-kolonitalsmetoden-nmkl-86-5-utg-2013 (accessed on 24 April 2021).
- ISO/TC 34/SC 9 ISO 8552:2004 IDF 132:2004 Specifies a Rapid Method for Estimating the Number of Psychrotrophic Microorganisms by Means of the Colony-Count Technique at 21 °C. Available online: https://www.iso.org/cms/render/live/en/sites/isoorg/contents/data/standard/03/18/31881.html (accessed on 24 April 2021).
- Priyashantha, H.; Lundh, Å.; Höjer, A.; Hetta, M.; Johansson, M.; Langton, M. Interactive Effects of Casein Micelle Size and Calcium and Citrate Content on Rennet-induced Coagulation in Bovine Milk. J. Texture Stud. 2019, 50, 508–519. [Google Scholar] [CrossRef]
- Johansson, M.; Högberg, M.; Andrén, A. Relation between AS1-Casein Content and Coagulation Properties of Milk from Swedish Dairy Goats. Open Food Sci. J. 2015, 9. [Google Scholar] [CrossRef]
- de Vries, R.; Brandt, M.; Lundh, Å.; Holtenius, K.; Hettinga, K.; Johansson, M. Short Communication: Influence of Shortening the Dry Period of Swedish Dairy Cows on Plasmin Activity in Milk. J. Dairy Sci. 2016, 99, 9300–9306. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Lundh, Å.; de Vries, R.; Sjaunja, K.S. Composition and Enzymatic Activity in Bulk Milk from Dairy Farms with Conventional or Robotic Milking Systems. J. Dairy Res. 2017, 84, 154–158. [Google Scholar] [CrossRef]
- Hara, A.; Radin, N.S. Lipid Extraction of Tissues with a Low-Toxicity Solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Appelqvist, L.-Å. Rapid Methods of Lipid Extraction and Fatty Acid Methyl Ester Preparation for Seed and Leaf Tissue with Special Remarks on Preventing the Accumulation of Lipid Contaminants; Almqvist & Wiksell: Stockholm, Sweden, 1968. [Google Scholar]
- Blomqvist, J.; Pickova, J.; Tilami, S.K.; Sampels, S.; Mikkelsen, N.; Brandenburg, J.; Sandgren, M.; Passoth, V. Oleaginous Yeast as a Component in Fish Feed. Sci. Rep. 2018, 8, 15945. [Google Scholar] [CrossRef]
- Rehn, U.; Petersen, M.A.; Saedén, K.H.; Ardö, Y. Ripening of Extra-Hard Cheese Made with Mesophilic DL-Starter. Int. Dairy J. 2010, 20, 844–851. [Google Scholar] [CrossRef]
- Wold, S.; Esbensen, K.; Geladi, P. Principal Component Analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Lindmark-Månsson, H.; Fondén, R.; Pettersson, H.-E. Composition of Swedish Dairy Milk. Int. Dairy J. 2003, 13, 409–425. [Google Scholar] [CrossRef]
- Lindmark-Månsson, H. Den Svenska Mejerimjölkens Sammansättning 2009; Svensk Mjölk Forskning; Swedish Dairy Association: Stockholm, Sweden, 2012. [Google Scholar]
- Verdier-Metz, I.; Coulon, J.-B.; Pradel, P. Relationship between Milk Fat and Protein Contents and Cheese Yield. Anim. Res. 2001, 50, 365–371. [Google Scholar] [CrossRef]
- Karlsson, M.A.; Langton, M.; Innings, F.; Wikström, M.; Lundh, Å.S. Short Communication: Variation in the Composition and Properties of Swedish Raw Milk for Ultra-High-Temperature Processing. J. Dairy Sci. 2017, 100, 2582–2590. [Google Scholar] [CrossRef] [PubMed]
- McSweeney, P.L.H.; Nursten, H.E.; Urbach, G. Flavours and off-Flavours in Milk and Dairy Products. In Advanced Dairy Chemistry Volume 3: Lactose, Water, Salts and Vitamins; Fox, P.F., Ed.; Springer: Boston, MA, USA, 1997; pp. 403–468. ISBN 978-1-4757-4409-5. [Google Scholar]
- Bynum, D.G.; Senyk, G.F.; Barbano, D.M. Determination of Free Fatty Acid Content of Cheddar Cheese by a Copper Soap Method. J. Dairy Sci. 1984, 67, 1521–1524. [Google Scholar] [CrossRef]
- Pastorino, A.J.; Hansen, C.L.; McMahon, D.J. Effect of PH on the Chemical Composition and Structure-Function Relationships of Cheddar Cheese. J. Dairy Sci. 2003, 86, 2751–2760. [Google Scholar] [CrossRef]
- France, T.C.; O’Mahony, J.A.; Kelly, A.L. The Plasmin System in Milk and Dairy Products. In Agents of Change: Enzymes in Milk and Dairy Products; Kelly, A.L., Larsen, L.B., Eds.; Food Engineering Series; Springer: Cham, Switzerland, 2021; pp. 11–55. ISBN 978-3-030-55482-8. [Google Scholar]
- Adda, J.; Gripon, J.C.; Vassal, L. The Chemistry of Flavour and Texture Generation in Cheese. Food Chem. 1982, 9, 115–129. [Google Scholar] [CrossRef]
- Fox, P.F. Proteolysis during Cheese Manufacture and Ripening. J. Dairy Sci. 1989, 72, 1379–1400. [Google Scholar] [CrossRef]
- Ardö, Y.; McSweeney, P.L.H.; Magboul, A.A.A.; Upadhyay, V.K.; Fox, P.F. Chapter 18-Biochemistry of Cheese Ripening: Proteolysis. In Cheese, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 445–482. ISBN 978-0-12-417012-4. [Google Scholar]
- Glantz, M.; Rosenlöw, M.; Lindmark-Månsson, H.; Johansen, L.B.; Hartmann, J.; Höjer, A.; Waak, E.; Löfgren, R.; Saedén, K.H.; Svensson, C.; et al. Impact of Protease and Lipase Activities on Quality of Swedish Raw Milk. Int. Dairy J. 2020, 104724. [Google Scholar] [CrossRef]
- Barbano, D.M.; Ma, Y.; Santos, M.V. Influence of Raw Milk Quality on Fluid Milk Shelf Life. J. Dairy Sci. 2006, 89, E15–E19. [Google Scholar] [CrossRef]
- Senyk, G.F.; Barbano, D.M.; Shipe, W.F. Proteolysis in Milk Associated with Increasing Somatic Cell Counts. J. Dairy Sci. 1985, 68, 2189–2194. [Google Scholar] [CrossRef]
- Ismail, B.; Nielsen, S.S. Invited Review: Plasmin Protease in Milk: Current Knowledge and Relevance to Dairy Industry. J. Dairy Sci. 2010, 93, 4999–5009. [Google Scholar] [CrossRef]
- Bobbo, T.; Cipolat-Gotet, C.; Bittante, G.; Cecchinato, A. The Nonlinear Effect of Somatic Cell Count on Milk Composition, Coagulation Properties, Curd Firmness Modeling, Cheese Yield, and Curd Nutrient Recovery. J. Dairy Sci. 2016, 99, 5104–5119. [Google Scholar] [CrossRef] [PubMed]
- Chassagne, M.; Barnouin, J.; Le Guenic, M. Expert Assessment Study of Milking and Hygiene Practices Characterizing Very Low Somatic Cell Score Herds in France. J. Dairy Sci. 2005, 88, 1909–1916. [Google Scholar] [CrossRef]
- Sant’Anna, A.C.; Paranhos da Costa, M.J.R. The Relationship between Dairy Cow Hygiene and Somatic Cell Count in Milk. J. Dairy Sci. 2011, 94, 3835–3844. [Google Scholar] [CrossRef]
- Silva, N.M.A.; Bastos, L.P.F.; Oliveira, D.L.S.; Oliveira, M.C.P.P.; Fonseca, L.M. Influence of Somatic Cell Count and Total Bacterial Counts of Raw Milk in Cheese Yield Using Small-Scale Methodology. Arq. Bras. Med. Veterinária E Zootec. 2012, 64, 1367–1372. [Google Scholar] [CrossRef]
- Auldist, M. Milk Quality and Udder Health|Effect on Processing Characteristics. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 902–907. ISBN 978-0-12-374407-4. [Google Scholar]
- Slaghuis, B.; Wolters, G.; Reinemann, D.J. Milking and Handling of Raw Milk|Milking Hygiene. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2002; pp. 632–637. ISBN 978-0-12-374407-4. [Google Scholar]
- Kelly, A.L.; Leitner, G.; Merin, U. Milk quality and udder health|test methods and standards. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 894–901. ISBN 978-0-12-374407-4. [Google Scholar]
- Stepaniak, L. Psychrotrophic Bacteria|Other Psychrotrophs. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 384–389. ISBN 978-0-12-374407-4. [Google Scholar]
- de Oliveira, G.B.; Favarin, L.; Luchese, R.H.; McIntosh, D. Psychrotrophic Bacteria in Milk: How Much Do We Really Know? Braz. J. Microbiol. 2015, 46, 313–321. [Google Scholar] [CrossRef]
- McPhee, J.D.; Griffiths, M.W. Psychrotrophic Bacteria|Pseudomonas spp. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 379–383. ISBN 978-0-12-374407-4. [Google Scholar]
- Reinemann, D.; Wolters, G.; Rasmussen, M. Review of Practices for Cleaning and Sanitation of Milking Machines. Int. Dairy Fed. Bull. 2003, 381, 3–18. [Google Scholar]
- García de Fernando, G. Lactic Acid Bacteria|Enterococcus in Milk and Dairy Products. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 153–159. ISBN 978-0-12-374407-4. [Google Scholar]
- Christiansen, P.; Nielsen, E.W.; Vogensen, F.K.; Brogren, C.-H.; Ardö, Y. Heat Resistance of Lactobacillus Paracasei Isolated from Semi-Hard Cheese Made of Pasteurised Milk. Int. Dairy J. 2006, 16, 1196–1204. [Google Scholar] [CrossRef]
- Glantz, M.; Devold, T.G.; Vegarud, G.E.; Månsson, H.L.; Stålhammar, H.; Paulsson, M. Importance of Casein Micelle Size and Milk Composition for Milk Gelation. J. Dairy Sci. 2010, 93, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- de Kruif, C.G. (Kees); Huppertz, T. Casein Micelles: Size Distribution in Milks from Individual Cows. J. Agric. Food Chem. 2012, 60, 4649–4655. [Google Scholar] [CrossRef]
- Sandra, S.; Ho, M.; Alexander, M.; Corredig, M. Effect of Soluble Calcium on the Renneting Properties of Casein Micelles as Measured by Rheology and Diffusing Wave Spectroscopy. J. Dairy Sci. 2012, 95, 75–82. [Google Scholar] [CrossRef]
- Frederiksen, P.D.; Hammershøj, M.; Bakman, M.; Andersen, P.N.; Andersen, J.B.; Qvist, K.B.; Larsen, L.B. Variations in Coagulation Properties of Cheese Milk from Three Danish Dairy Breeds as Determined by a New Free Oscillation Rheometry-Based Method. Dairy Sci. Technol. 2011, 91, 309–321. [Google Scholar] [CrossRef]
- Hallén, E.; Allmere, T.; Näslund, J.; Andrén, A.; Lundén, A. Effect of Genetic Polymorphism of Milk Proteins on Rheology of Chymosin-Induced Milk Gels. Int. Dairy J. 2007, 17, 791–799. [Google Scholar] [CrossRef]
- Lucas, A.; Rock, E.; Chamba, J.-F.; Verdier-Metz, I.; Brachet, P.; Coulon, J.-B. Respective Effects of Milk Composition and the Cheese-Making Process on Cheese Compositional Variability in Components of Nutritional Interest. Le Lait 2006, 86, 21–41. [Google Scholar] [CrossRef]
- Lu, J.; Langton, M.; Sampels, S.; Pickova, J. Lipolysis and Oxidation in Ultra-High Temperature Milk Depend on Sampling Month, Storage Duration, and Temperature. J. Food Sci. 2019, 84, 1045–1053. [Google Scholar] [CrossRef]
- Ferlay, A.; Agabriel, C.; Sibra, C.; Journal, C.; Martin, B.; Chilliard, Y. Tanker Milk Variability in Fatty Acids According to Farm Feeding and Husbandry Practices in a French Semi-Mountain Area. Dairy Sci. Technol. 2008, 88, 193–215. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Corl, B.A.; Dwyer, D.A.; Saebø, A.; Bauman, D.E. Identification of the Conjugated Linoleic Acid Isomer That Inhibits Milk Fat Synthesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R179–R184. [Google Scholar] [CrossRef]
- Glover, K.E.; Budge, S.; Rose, M.; Rupasinghe, H.P.V.; MacLaren, L.; Green-Johnson, J.; Fredeen, A.H. Effect of Feeding Fresh Forage and Marine Algae on the Fatty Acid Composition and Oxidation of Milk and Butter. J. Dairy Sci. 2012, 95, 2797–2809. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, Rumen Biohydrogenation and Nutritional Quality of Cow and Goat Milk Fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Wilkinson, M.G. Acceleration of Cheese Ripening. In Cheese: Chemistry, Physics and Microbiology: Volume 1 General Aspects; Fox, P.F., Ed.; Springer: Boston, MA, USA, 1993; pp. 523–555. ISBN 978-1-4615-2650-6. [Google Scholar]
- Frétin, M.; Martin, B.; Buchin, S.; Desserre, B.; Lavigne, R.; Tixier, E.; Cirié, C.; Bord, C.; Montel, M.-C.; Delbès, C.; et al. Milk Fat Composition Modifies the Texture and Appearance of Cantal-Type Cheeses but Not Their Flavor. J. Dairy Sci. 2019, 102, 1131–1143. [Google Scholar] [CrossRef]
- Hurtaud, C.; Peyraud, J.L.; Michel, G.; Berthelot, D.; Delaby, L. Winter Feeding Systems and Dairy Cow Breed Have an Impact on Milk Composition and Flavour of Two Protected Designation of Origin French Cheeses. Animal 2009, 3, 1327–1338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coppa, M.; Ferlay, A.; Monsallier, F.; Verdier-Metz, I.; Pradel, P.; Didienne, R.; Farruggia, A.; Montel, M.C.; Martin, B. Milk Fatty Acid Composition and Cheese Texture and Appearance from Cows Fed Hay or Different Grazing Systems on Upland Pastures. J. Dairy Sci. 2011, 94, 1132–1145. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | p | Sampling Month | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| February-16 | March | April | May | June | July | August | September | October | November | December | January | 17-February | |||
| n = 2 | n = 4 | n = 5 | n = 7 | n = 5 | n = 7 | n = 7 | n = 1 | n = 5 | n = 6 | n = 7 | n = 4 | n = 5 | |||
| Fat | Mean | ** | 4.39 abc | 4.43 abc | 4.36 abc | 4.43 abc | 4.17 bc | 4.28 abc | 4.19 c | 4.40 abc | NA | 4.42 a | 4.45 a | 4.39 ab | 4.36 ab |
| (g/100 g) | (SD) | (0.00) | (0.00) | (0.09) | (0.09) | (0.07) | (0.22) | (0.05) | (0.00) | NA | (0.09) | (0.04) | (0.04) | (0.10) | |
| Protein | Mean | ** | 3.50 abc | 3.47 abc | 3.52 abc | 3.45 bc | 3.43 c | 3.46 abc | 3.44 c | 3.53 abc | NA | 3.59 a | 3.59 a | 3.57 ab | 3.48 abc |
| (g/100 g) | (SD) | (0.04) | (0.00) | (0.01) | (0.02) | (0.01) | (0.00) | (0.02) | (0.00) | NA | (0.06) | (0.04) | (0.03) | (0.23) | |
| Urea | Mean | 0.55 | 4.35 | 4.20 | 4.05 | 4.00 | 4.60 | 4.00 | 4.24 | 4.10 | NA | 4.40 | 4.05 | 3.97 | 4.16 |
| (mmol/L) | (SD) | (0.35) | (0.00) | (0.35) | (0.28) | (0.85) | (0.14) | (0.21) | (0.00) | NA | (0.28) | (0.37) | (0.28) | (0.22) | |
| FFA | Mean | 0.99 | 0.82 | 0.81 | 0.74 | 0.88 | 0.83 | 0.72 | 0.76 | 0.81 | NA | 0.80 | 0.87 | 0.74 | 0.83 |
| (mmol/100 g fat) | (SD) | (0.16) | (0.00) | (0.00) | (0.15) | (0.47) | (0.00) | (0.14) | (0.00) | NA | (0.05) | (0.26) | (0.02) | (0.09) | |
| pH | Mean | ** | 6.73 abc | 6.70 abc | 6.72 ab | 6.74 ab | 6.80 ab | 6.73 ab | 6.76 ab | 6.48 c | 6.67 bc | 6.78 ab | 6.70 abc | 6.70 abc | 6.81 a |
| (SD) | (0.02) | (0.02) | (0.03) | (0.03) | (0.10) | (0.00) | (0.06) | (0.00) | (0.02) | (0.02) | (0.03) | (0.00) | (0.00) | ||
| PL | Mean | * | NA | 2.91 a | NA | 2.70 a | 2.81 a | NA | 1.56 c | NA | 2.67 ab | NA | 1.80 bc | NA | 2.39 abc |
| (units/mL) | (SD) | NA | (0.76) | NA | (0.73) | (0.32) | NA | (0.54) | NA | (0.73) | NA | (1.03) | NA | (0.94) | |
| PG | Mean | ** | NA | 70.83 a | NA | 66.59 a | 64.44 ab | NA | 43.51 c | NA | 58.37 abc | NA | 48.89 bc | NA | 48.72 bc |
| (units/mL) | (SD) | NA | (4.88) | NA | (9.91) | (7.63) | NA | (9.74) | NA | (8.63) | NA | (6.69) | NA | (9.28) | |
| TP | Mean | ** | NA | 32.06 ab | NA | 33.82 ab | 29.23 bc | 40.53 a | NA | NA | NA | NA | 20.73 c | NA | NA |
| (mM Leuc. Eq.) | (SD) | NA | (1.17) | NA | (2.74) | (4.33) | (8.88) | NA | NA | NA | NA | (4.78) | NA | NA | |
| SCC | Mean | ** | 174 abc | 180 abc | 133.5 c | 182.5 abc | 221 ab | 197 abc | 233.8 a | 231 abc | NA | 174.8 bc | 191.3 abc | 174.2 bc | 197.2 abc |
| (103/mL) | (SD) | (9.90) | (5) | (7.78) | (30.4) | (20.7) | (21) | (41.1) | (0) | NA | (6.5) | (25.9) | (18.8) | (18.4) | |
| TBC | Mean | ** | 27 b | 32 b | 15.2 b | 50.3 b | 58.2 ab | 60.4 ab | 109 a | 53 ab | 14.5 b | 64.0 ab | 65 b | 21.2 b | 36.6 b |
| (CFU 103/mL) | (SD) | (8.7) | (12.19) | (10.8) | (29.6) | (12.15) | (36.0) | (58.4) | (0) | (5.0) | (59) | (113) | (9.3) | (25.3) | |
| PBC | Mean | 0.87 | 1.6 | 12.7 | 1.4 | 4.6 | 7.5 | 12.1 | 11.02 | 1.60 | 7.1 | 6.5 | 11.2 | 5.3 | 4.5 |
| (CFU 103/mL) | (SD) | (0.7) | (13.7) | (1.3) | (2.9) | (5.9) | (15.9) | (12.4) | (0) | (9.8) | (7.8) | (17.5) | (4.1) | (3.4) | |
| TRBC | Mean | 0.06 | 7.5 | 2.0 | 6.7 | 6.7 | 9.9 | 6.5 | 3.5 | 1.0 | NA | 1.9 | 0.7 | 1.1 | 4.3 |
| (CFU 103/mL) | (SD) | (2.1) | (0) | (4.6) | (4.5) | (0) | (5.5) | (2.4) | (0) | NA | (1.1) | (0.3) | (0.5) | (2.9) | |
| CMS | Mean | ** | 170 ab | 153 abc | 138 abc | 129 bc | 130 bc | NA | 80 c | 117 abc | 127 bc | NA | 192 a | 200 a | 197 a |
| (nm) | (SD) | (1) | (16) | (24) | (39) | (10) | NA | (2) | (0) | (29) | NA | (38) | (28) | (9) | |
| G20 | Mean | * | 60 ab | 66 ab | 66 ab | 67 a | 60 ab | 52 ab | 58 ab | NA | 65 ab | NA | 57 ab | NA | 42 b |
| (Pa) | (SD) | (5) | (3) | (2) | (8) | (3) | (10) | (21) | NA | (14) | NA | (5) | NA | (14) | |
| RCT | Mean | 0.87 | 504 | 457 | 467 | 450 | 471 | 494 | 461 | NA | 564 | NA | 462 | NA | 581 |
| (s) | (SD) | (40) | (11) | (11) | (39) | (11) | (59) | (80) | NA | (174) | NA | (128) | NA | (50) | |
| Fatty Acids | p | Sampling Month | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| February-16 | March | April | May | June | July | August | September | October | November | December | January | 17-February | |||
| n = 2 | n = 2 | n = 2 | n = 5 | n = 4 | n = 4 | n = 6 | n = 2 | n = 3 | n = 6 | n = 8 | n = 4 | n = 3 | |||
| C8:0 (Caprylic acid) | Mean | 0.72 | 0.16 | 0.13 | 0.00 | 0.04 | 0.05 | 0.13 | 0.06 | 0.00 | 0.02 | 0.00 | 0.07 | 0.13 | 0.18 |
| (SD) | (0.22) | (0.18) | (0.00) | (0.09) | (0.05) | (0.26) | (0.15) | (0.00) | (0.02) | (0.00) | (0.11) | (0.15) | (0.16) | ||
| C10:0 (Capric acid) | Mean | ** | 0.36 ab | 0.62 ab | 0.07 ab | 0.34 ab | 0.50 ab | 0.24 b | 1.01 a | 0.69 ab | 1.00 | 0.90 ab | 0.62 ab | 0.39 ab | 0.20 ab |
| (SD) | (0.29) | (0.49) | (0.10) | (0.25) | (0.44) | (0.27) | (0.31) | (0.68) | (0.57) | (0.43) | (0.16) | (0.04) | (0.18) | ||
| C12:0 (Lauric acid) | Mean | ** | 3.12 abc | 3.01 abc | 2.59 abc | 2.98 abc | 2.57 bc | 2.41 c | 3.30 ab | 2.52 abc | 3.38 abc | 3.61 a | 3.45 a | 3.27 abc | 2.66 abc |
| (SD) | (0.14) | (0.23) | (0.27) | (0.25) | (0.55) | (0.39) | (0.43) | (0.49) | (0.35) | (0.21) | (0.29) | (0.32) | (0.81) | ||
| C14:0 (Myristoleic acid) | Mean | ** | 13.85 ab | 13.64 abc | 13.66 abc | 13.57 ab | 12.74 bc | 12.54 c | 13.16 abc | 12.69 abc | 13.27 abc | 13.79 a | 13.79 a | 13.69 a | 13.80 a |
| (SD) | (0.17) | (0.43) | (0.08) | (0.07) | (0.39) | (0.82) | (0.47) | (0.00) | (0.16) | (0.30) | (0.20) | (0.48) | (0.45) | ||
| C15:0 (Pentadecylic acid) | Mean | 0.20 | 1.17 | 1.43 | 1.21 | 1.17 | 1.14 | 1.26 | 1.25 | 1.14 | 1.07 | 1.11 | 1.31 | 1.34 | 1.19 |
| (SD) | (0.03) | (0.03) | (0.04) | (0.07) | (0.04) | (0.07) | (0.16) | (0.01) | (0.33) | (0.25) | (0.16) | (0.15 | (0.03) | ||
| C16:0 (Palmitoleic acid) | Mean | * | 38.59 abc | 37.73 abc | 39.09 abc | 39.01 a | 36.99 abc | 36.13 bc | 36.46 c | 38.08 abc | 37.13 abc | 37.47 abc | 38.05 abc | 37.48 abc | 38.20 abc |
| (SD) | (0.01) | (0.53) | (1.16) | (0.27) | (1.68) | (0.89) | (0.86) | (1.00) | (2.02) | (1.52) | (0.83) | (1.16) | (0.50) | ||
| C17:0 (Margaric acid) | Mean | * | 0.55 ab | 0.55 ab | 0.62 ab | 0.53 b | 0.58 ab | 0.68 a | 0.56 ab | 0.52 ab | 0.55 ab | 0.55 ab | 0.56 ab | 0.66 ab | 0.57 ab |
| (SD) | (0.00) | (0.04) | (0.09) | (0.02) | (0.04) | (0.13) | (0.03) | (0.02) | (0.04) | (0.04) | (0.03) | (0.14) | (0.03) | ||
| C18:0 (Stearic acid) | Mean | ** | 12.28 abc | 12.06 abc | 12.10 abc | 12.16 bc | 13.22 ab | 13.53 a | 12.58 abc | 12.97 abc | 12.18 abc | 12.39 abc | 11.96 c | 12.19 bc | 12.52 abc |
| (SD) | (0.60) | (0.14) | (0.09) | (0.22) | (0.63) | (0.37) | (0.70) | (0.06) | (0.53) | (0.64) | (0.45) | (0.21) | (0.85) | ||
| C20:0 (Arachidic acid) | Mean | 0.35 | 0.20 | 0.18 | 0.23 | 0.23 | 0.19 | 0.20 | 0.31 | 0.20 | 0.27 | 0.20 | 0.21 | 0.39 | 0.24 |
| (SD) | (0.01) | (0.01) | (0.04) | (0.05) | (0.09) | (0.05) | (0.18) | (0.02) | (0.07) | (0.05) | (0.05) | (0.22) | (0.21) | ||
| C14:1 (Myristoleic acid) | Mean | ** | 1.13 abcd | 1.29 abcd | 1.11 abcd | 1.23 abcd | 1.05 cd | 1.05 bd | 1.14 abcd | 1.05 abcd | 1.32 ab | 1.20 abcd | 1.27 a | 1.19 abcd | 1.14 abcd |
| (SD) | (0.04) | (0.06) | (0.03) | (0.15) | (0.05) | (0.09) | (0.15) | (0.01) | (0.09) | (0.05) | (0.13) | (0.02) | (0.04) | ||
| C16:1 (Palmitoleic acid) | Mean | 0.15 | 1.39 | 1.91 | 2.02 | 1.55 | 1.57 | 1.72 | 1.68 | 1.71 | 1.80 | 1.60 | 1.81 | 1.92 | 1.80 |
| (SD) | (0.60) | (0.16) | (0.34) | (0.23) | (0.13) | (0.14) | (0.22) | (0.03) | (0.11) | (0.24) | (0.24) | (0.38) | (0.14) | ||
| C18:1 (Oleic acid) | Mean | ** | 23.84 bcd | 23.34 cd | 23.70 cd | 23.59 cd | 25.54 ab | 26.33 a | 24.51 bc | 25.16 abc | 24.12 bcd | 23.75 cd | 23.30 d | 23.42 cd | 23.81 cd |
| (SD) | (0.08) | (0.48) | (0.25) | (0.56) | (0.59) | (0.86) | (0.86) | (0.06) | (0.31) | (0.62) | (0.45) | (0.28) | (0.73) | ||
| C18:2 (Linoleic acid) | Mean | 0.73 | 1.69 | 1.76 | 1.75 | 1.71 | 1.96 | 1.88 | 1.85 | 1.66 | 1.99 | 1.74 | 1.81 | 1.89 | 1.74 |
| (SD) | (0.00) | (0.19) | (0.15) | (0.15) | (0.33) | (0.20) | (0.21) | (0.01) | (0.39) | (0.07) | (0.33) | (0.17) | (0.07) | ||
| C18:3 (Linolenic acid) | Mean | 0.78 | 0.58 | 0.55 | 0.55 | 0.59 | 0.63 | 0.58 | 0.62 | 0.52 | 0.66 | 0.61 | 0.59 | 0.69 | 0.70 |
| (SD) | (0.03) | (0.02) | (0.00) | (0.13) | (0.08) | (0.05) | (0.11) | (0.00) | (0.10) | (0.08) | (0.09) | (0.18) | (0.23) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Priyashantha, H.; Johansson, M.; Langton, M.; Sampels, S.; Jayarathna, S.; Hetta, M.; Saedén, K.H.; Höjer, A.; Lundh, Å. Variation in Dairy Milk Composition and Properties Has Little Impact on Cheese Ripening: Insights from a Traditional Swedish Long-Ripening Cheese. Dairy 2021, 2, 336-355. https://doi.org/10.3390/dairy2030027
Priyashantha H, Johansson M, Langton M, Sampels S, Jayarathna S, Hetta M, Saedén KH, Höjer A, Lundh Å. Variation in Dairy Milk Composition and Properties Has Little Impact on Cheese Ripening: Insights from a Traditional Swedish Long-Ripening Cheese. Dairy. 2021; 2(3):336-355. https://doi.org/10.3390/dairy2030027
Chicago/Turabian StylePriyashantha, Hasitha, Monika Johansson, Maud Langton, Sabine Sampels, Shishanthi Jayarathna, Mårten Hetta, Karin Hallin Saedén, Annika Höjer, and Åse Lundh. 2021. "Variation in Dairy Milk Composition and Properties Has Little Impact on Cheese Ripening: Insights from a Traditional Swedish Long-Ripening Cheese" Dairy 2, no. 3: 336-355. https://doi.org/10.3390/dairy2030027
APA StylePriyashantha, H., Johansson, M., Langton, M., Sampels, S., Jayarathna, S., Hetta, M., Saedén, K. H., Höjer, A., & Lundh, Å. (2021). Variation in Dairy Milk Composition and Properties Has Little Impact on Cheese Ripening: Insights from a Traditional Swedish Long-Ripening Cheese. Dairy, 2(3), 336-355. https://doi.org/10.3390/dairy2030027