Biodegradation and Microbial Contamination of Limestone Surfaces: An Experimental Study from Batalha Monastery, Portugal

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Location and Sampling
2.2. Air Monitoring
2.3. Molecular Analysis of Bacterial Communities
2.4. Morphology Observation, Mineralogical, and Chemical Characterization of the Bio-Deteriorated Stone
2.4.1. Optical Microscopy (OM)
2.4.2. X-Ray Micro-Diffractometry (µ-XRD)
2.4.3. Low-Vacuum Scanning Electron Microscopy Coupled with Energy-Dispersive Spectrometry (LV-SEM + EDS)
3. Results
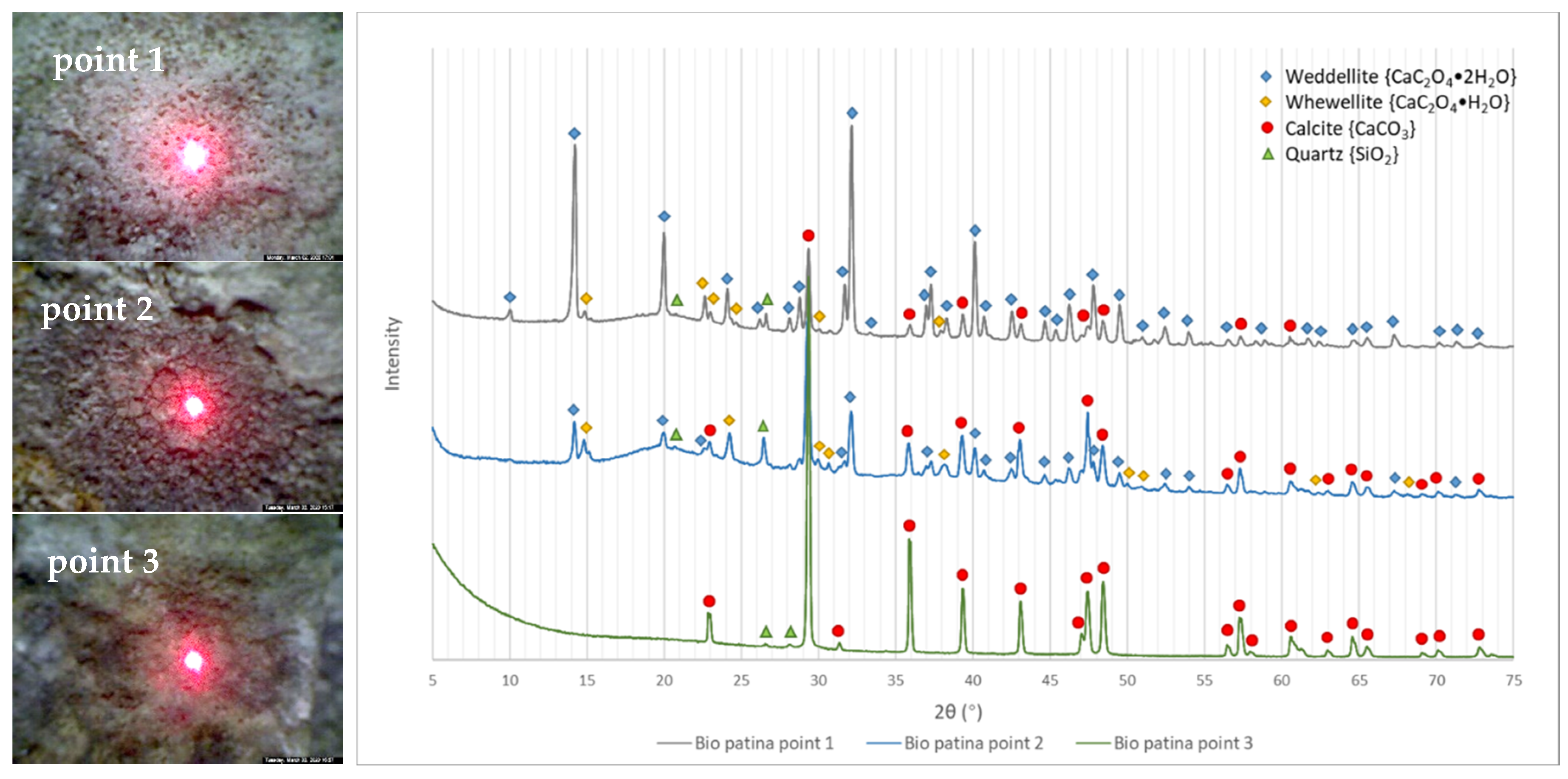
3.1. Optical Microscope Observation
3.2. XRD Results
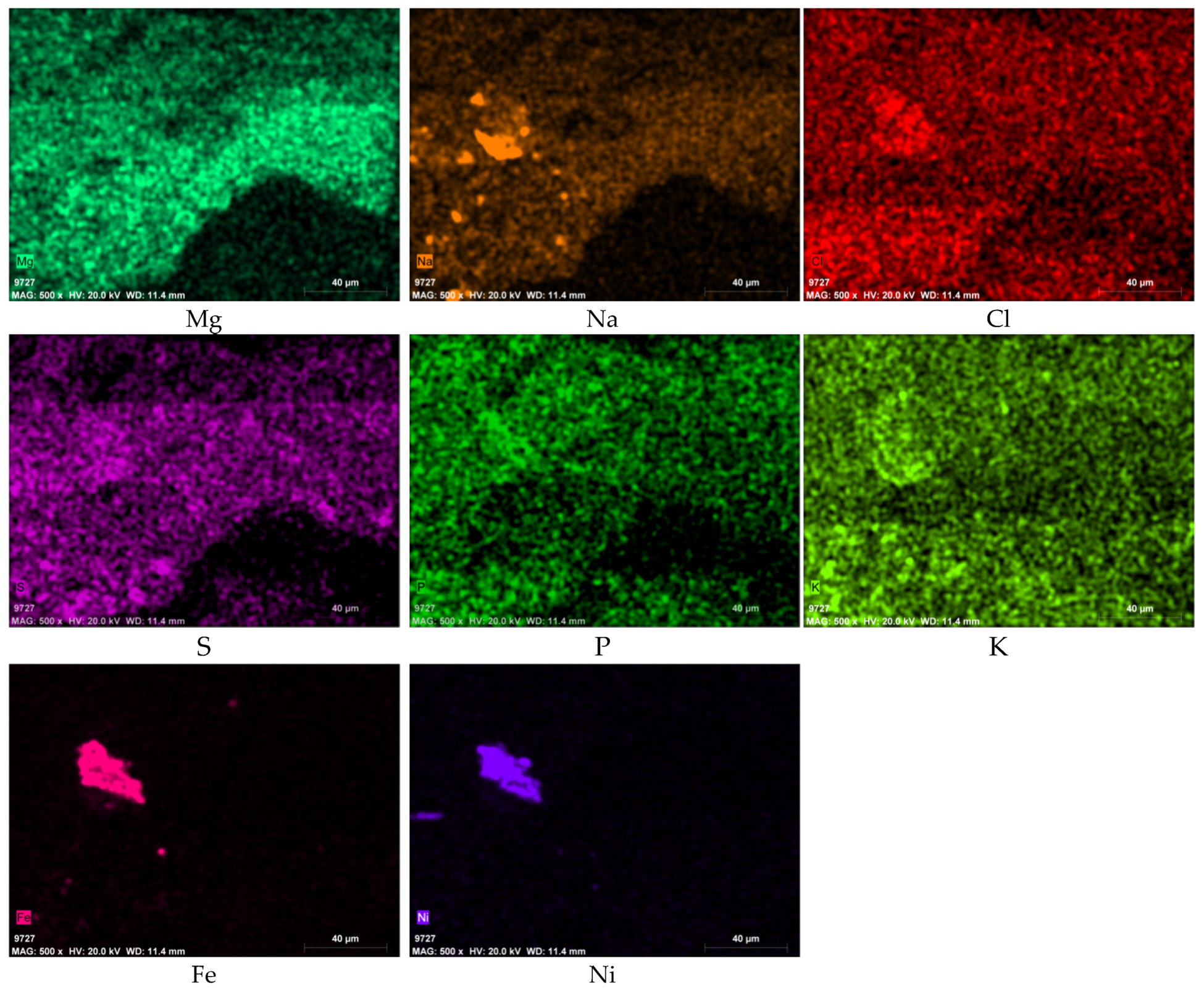
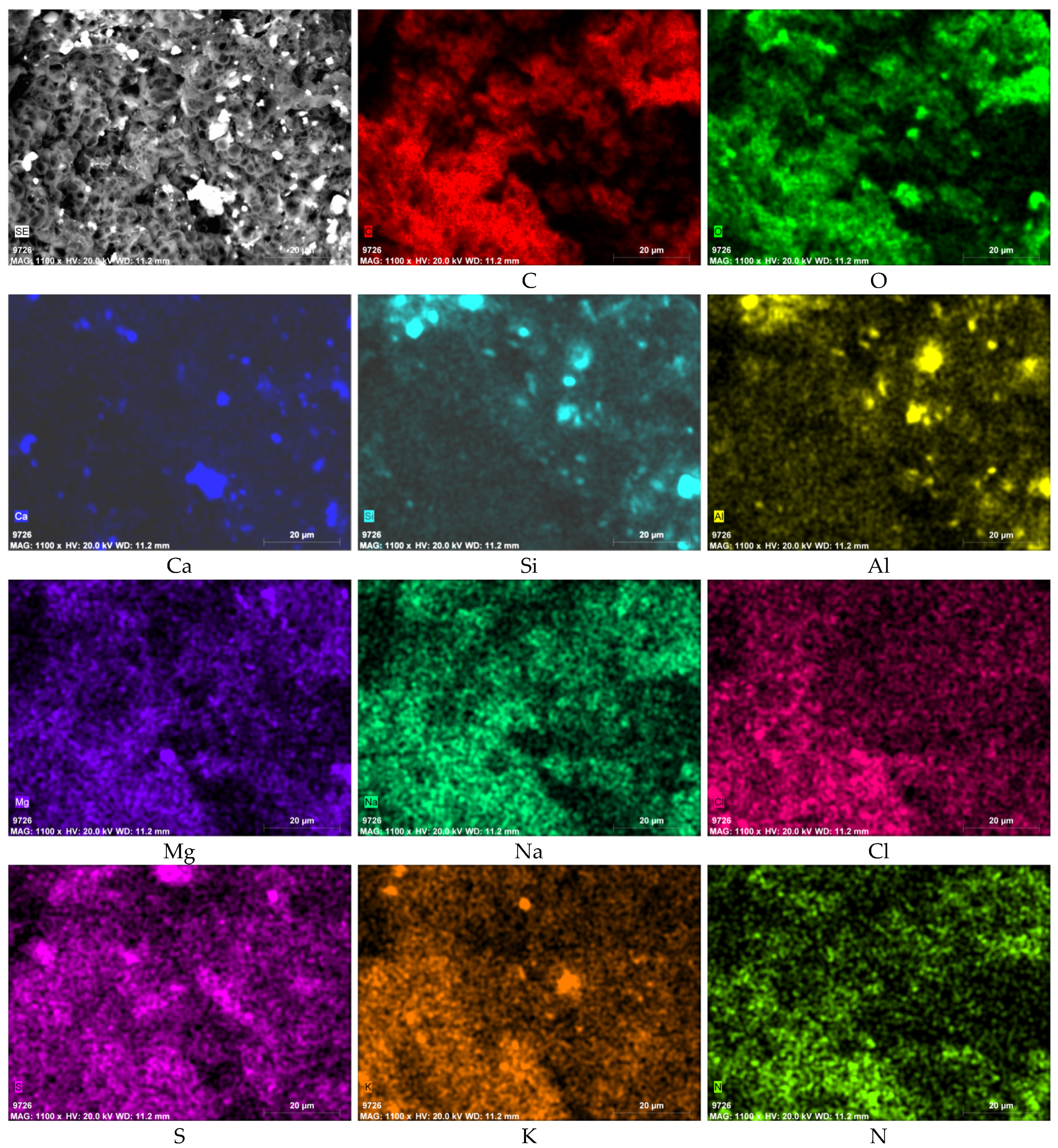
3.3. SEM-EDS Results
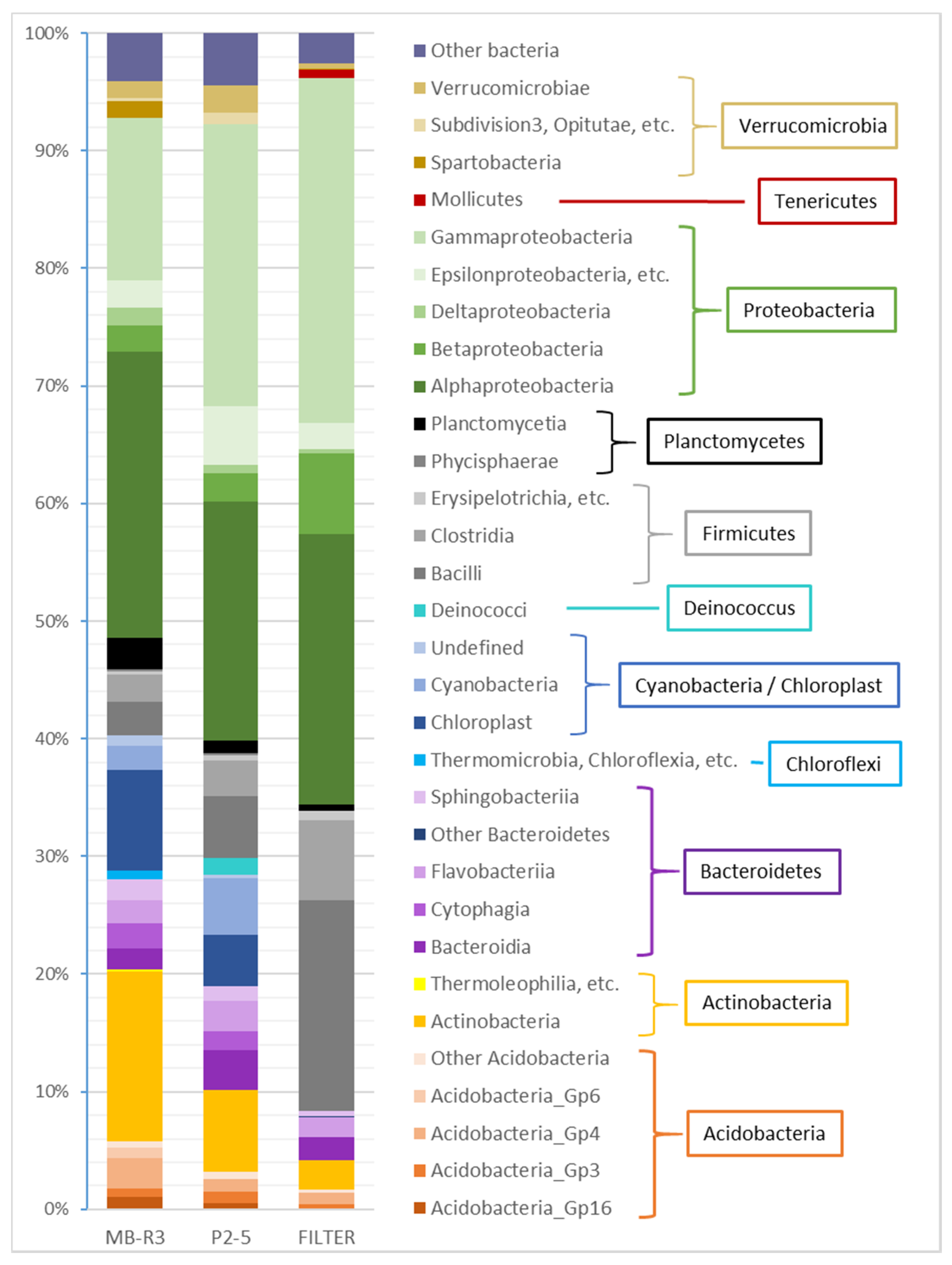
3.4. High-Throughput Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Ahmadjian, V. Resynthesis of lichens. In The Lichens; Elsevier Inc.: Cambridge, MA, USA; Academic Press: Cambridge, MA, USA, 1973; pp. 565–579. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Grube, M.; Cernava, T.; Soh, J.; Fuchs, S.; Aschenbrenner, I.; Lassek, C.; Wegner, U.; Becher, D.; Riedel, K.; Sensen, C.W.; et al. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. 2015, 9, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Trovão, J.; Portugal, A.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Pinheiro, A.C.; Catarino, L.; Gil, F.; Tiago, I. Fungal diversity and distribution across distinct biodeterioration phenomena in limestone walls of the old cathedral of Coimbra, UNESCO World Heritage Site. Int. Biodeterior. Biodegrad. 2019, 142, 91–102. [Google Scholar] [CrossRef]
- Pinheiro, A.C.; Mesquita, N.; Trovão, J.; Soares, F.; Tiago, I.; Coelho, C.; Carvalho, H.P.; Gil, F.; Catarino, L.; Piñar, G.; et al. Limestone biodeterioration: A review on the Portuguese cultural heritage scenario. J. Cult. Herit. 2019, 36, 275–285. [Google Scholar] [CrossRef]
- Macedo, M.F.; Miller, A.Z.; Dionísio, A.; Saiz-Jimenez, C. Biodiversity of cyanobacteria and green algae on monuments in the Mediterranean Basin: An overview. Microbiology 2009, 155, 3476–3490. [Google Scholar] [CrossRef] [PubMed]
- Prada, J.L.; Pugés, M.; Rocabayera, R. Estudio de las patologias del Retablo del Altar Mayor de la Sé da Guarda. In Património Estudos; Instituto Português do Património Arquitectónico: Lisbon, Portugal, 2002; p. 70. [Google Scholar]
- Ascaso, C.; Wierzchos, J.; Delgado Rodrigues, J.; Aires-Barros, L.; Henriques, F.; Charola, A.E. Endolithic microorganisms in the biodeterioration of the tower of Belem. Int. Z. FürBauinstandsetz. 1998, 4, 627–640. [Google Scholar]
- Ascaso, C.; Wierzchos, J.; Souza-Egipsy, V.; De los Rıos, A.; Rodrigues, J.D. In situ evaluation of the biodeteriorating action of microorganisms and the effects of biocides on carbonate rock of the Jeronimos Monastery (Lisbon). Int. Biodeterior. Biodegrad. 2002, 49, 1–12. [Google Scholar] [CrossRef]
- Mateus, D.M.; Silva, R.B.; Costa, F.M.; Coroado, J.P. Diversidade microbiológica do edifício da Sacristia Incompleta do Convento de Cristo, em Tomar, e avaliação do seu controlo por biocidas. Conserv. Património 2013, 17, 11–17. [Google Scholar] [CrossRef]
- Schiavon, N.; Chiavari, G.; Schiavon, G.; Fabbri, D. Nature and decay effects of urban soiling on granitic building stones. Sci. Total Environ. 1995, 167, 87–101. [Google Scholar] [CrossRef]
- Schiavon, N. Biodeterioration of calcareous and granitic building stones in urban environments. Geol. Soc. Lond. Spec. Publ. 2002, 205, 195–205. [Google Scholar] [CrossRef]
- Ortega-Morales, O.; Montero-Muñoz, J.L.; Neto, J.A.B.; Beech, I.B.; Sunner, J.; Gaylarde, C. Deterioration and microbial colonization of cultural heritage stone buildings in polluted and unpolluted tropical and subtropical climates: A meta-analysis. Int. Biodeterior. Biodegrad. 2019, 143, 104734. [Google Scholar] [CrossRef]
- Ding, Y.; Mirão, J.; Redol, P.; Dias, L.; Moita, P.; Angelini, E.; Grassini, S.; Schiavon, N. A combined petrographic and geochemical metrological approach to assess the provenance of the building limestone used in the Batalha Monastery (Portugal). In Metrology for Archaeology and Cultural Heritage; Battisti, V., Gallo, V., Eds.; IMEKO: Florence, Italy, 2019; p. 338. [Google Scholar]
- Dias, L.; Rosado, T.; Coelho, A.; Barrulas, P.; Lopes, L.; Moita, P.; Candeias, A.; Mirão, J.; Caldeira, A.T. Natural limestone discolouration triggered by microbial activity—A contribution. AIMS Microbiol. 2018, 4, 594. [Google Scholar] [CrossRef] [PubMed]
- Rosado, T.; Dias, L.; Lança, M.; Nogueira, C.; Santos, R.; Martins, M.R.; Candeias, A.; Mirão, J.; Caldeira, A.T. Assessment of microbiota present on a Portuguese historical stone convent using high-throughput sequencing approaches. MicrobiologyOpen 2020, 9, 1067–1084. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.; Rosado, T.; Candeias, A.; Mirão, J.; Caldeira, A.T. A change in composition, a change in colour: The case of limestone sculptures from the Portuguese National Museum of Ancient Art. J. Cult. Herit. 2020, 42, 255–262. [Google Scholar] [CrossRef]
- Mifsud, S.; Lanfranco, E.; Fiorentino, J.; Mifsud, S.D. An Updated Flora of Selmunett (St. Paul’s Island) including Mosses and Lichens. XJENZA 2016, 4, 142–159. [Google Scholar]
- Lisci, M.; Monte, M.; Pacini, E. Lichens and higher plants on stone: A review. Int. Biodeterior. Biodegrad. 2003, 51, 1–17. [Google Scholar] [CrossRef]
- Sayer, J.A.; Kierans, M.; Gadd, G.M. Solubilisation of some naturally occurring metal-bearing minerals, limescale and lead phosphate by Aspergillus niger. FEMS Microbiol. Lett. 1997, 154, 29–35. [Google Scholar] [CrossRef]
- Gharieb, M.M.; Sayer, J.A.; Gadd, G.M. Solubilization of natural gypsum (CaSO4.2H2O) and the formation of calcium oxalate by Aspergillus niger and Serpula himantioides. Mycol. Res. 1998, 102, 825–830. [Google Scholar] [CrossRef]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments—an updated overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef]
- Shi, J.; Wang, N.; Gao, H.; Baker, A.; Yao, X.; Zhang, D. Phosphorus solubility in aerosol particles related to particle sources and atmospheric acidification in Asian continental outflow. Atmos. Chem. Phys. 2019, 19, 847–860. [Google Scholar] [CrossRef]
- Leiva, D.; Clavero-León, C.; Carú, M.; Orlando, J. Intrinsic factors of Peltigera lichens influence the structure of the associated soil bacterial microbiota. FEMS Microbiol. Ecol. 2016, 92, fiw178. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.T.; Cropsey, G.W.; Caporaso, J.G.; Knight, R.; Fierer, N. Bacterial communities associated with the lichen symbiosis. Appl. Environ. Microbiol. 2011, 77, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, T.A.; Dedysh, S.N.; Zavarzin, G.A. The leading role of actinobacteria in aerobic cellulose degradation in Sphagnum peat bogs. Dokl. Biol. Sci. 2006, 410, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.A.; Danko, D.C.; Sandoval, T.A.; Pishchany, G.; Moncada, B.; Kolter, R.; Mason, C.E.; Zambrano, M.M. The microbiomes of seven lichen genera reveal host specificity, a reduced core community and potential as source of antimicrobials. Front. Microbiol. 2020, 11, 398. [Google Scholar] [CrossRef]
- Bjelland, T.; Grube, M.; Hoem, S.; Jorgensen, S.L.; Daae, F.L.; Thorseth, I.H.; Øvreås, L. Microbial metacommunities in the lichen–rock habitat. Environ. Microbiol. Rep. 2011, 3, 434–442. [Google Scholar] [CrossRef]
- Lee, Y.M.; Kim, E.H.; Lee, H.K.; Hong, S.G. Biodiversity and physiological characteristics of Antarctic and Arctic lichens-associated bacteria. World J. Microbiol. Biotechnol. 2014, 30, 2711–2721. [Google Scholar] [CrossRef]
- Özvan, A.; Dinçer, İ.; Akın, M.; Oyan, V.; Tapan, M. Experimental studies on ignimbrite and the effect of lichens and capillarity on the deterioration of Seljuk Gravestones. Eng. Geol. 2015, 185, 81–95. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Salvador, C.S.C.; Caldeira, A.T.; Angelini, E.; Schiavon, N. Biodegradation and Microbial Contamination of Limestone Surfaces: An Experimental Study from Batalha Monastery, Portugal. Corros. Mater. Degrad. 2021, 2, 31-45. https://doi.org/10.3390/cmd2010002
Ding Y, Salvador CSC, Caldeira AT, Angelini E, Schiavon N. Biodegradation and Microbial Contamination of Limestone Surfaces: An Experimental Study from Batalha Monastery, Portugal. Corrosion and Materials Degradation. 2021; 2(1):31-45. https://doi.org/10.3390/cmd2010002
Chicago/Turabian StyleDing, Yufan, Catia Sofia Clemente Salvador, Ana Teresa Caldeira, Emma Angelini, and Nick Schiavon. 2021. "Biodegradation and Microbial Contamination of Limestone Surfaces: An Experimental Study from Batalha Monastery, Portugal" Corrosion and Materials Degradation 2, no. 1: 31-45. https://doi.org/10.3390/cmd2010002
APA StyleDing, Y., Salvador, C. S. C., Caldeira, A. T., Angelini, E., & Schiavon, N. (2021). Biodegradation and Microbial Contamination of Limestone Surfaces: An Experimental Study from Batalha Monastery, Portugal. Corrosion and Materials Degradation, 2(1), 31-45. https://doi.org/10.3390/cmd2010002