Sleep Network Deterioration as a Function of Dim-Light-At-Night Exposure Duration in a Mouse Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Surgeries
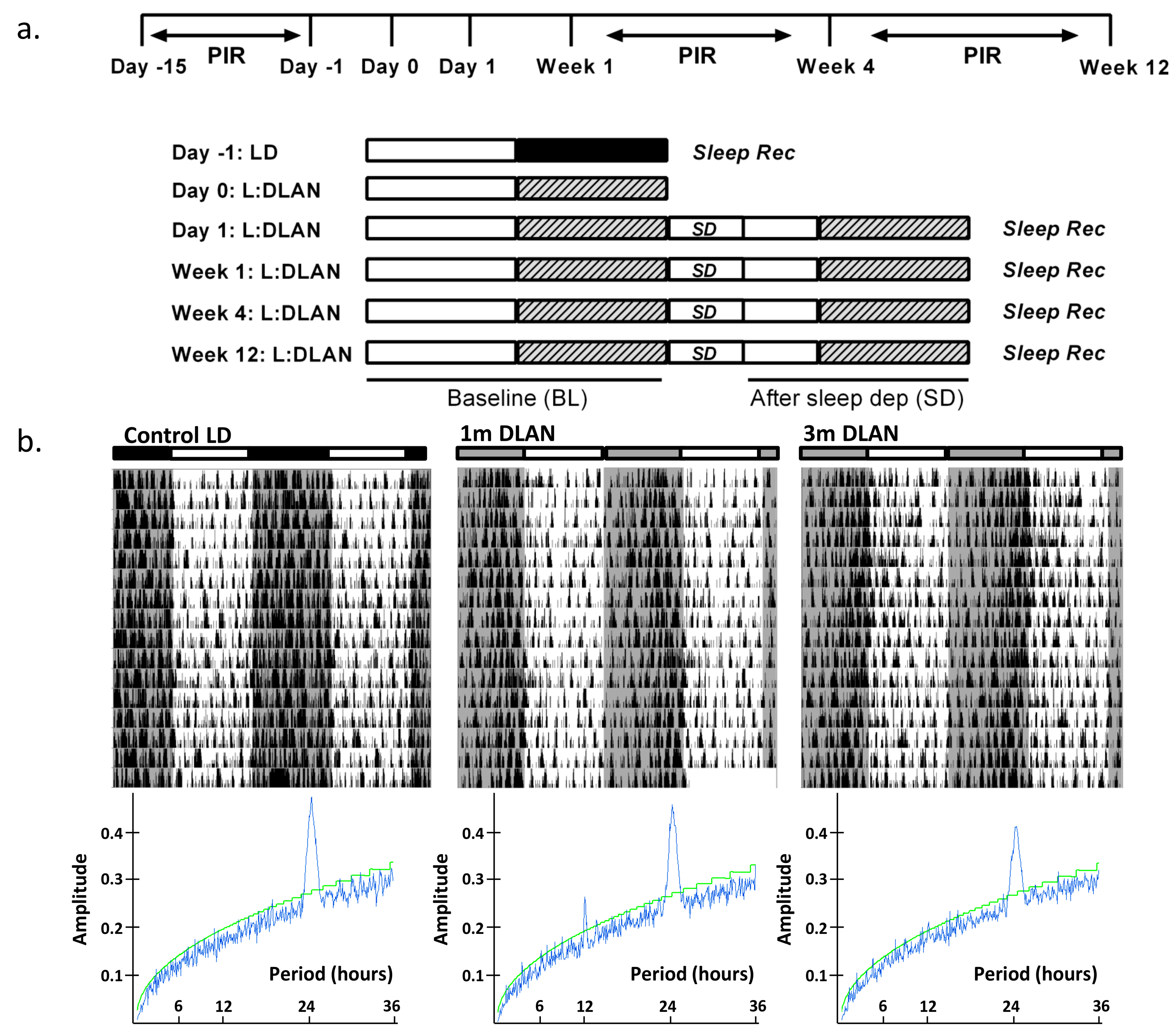
2.3. Light Schedules and Behavioral Recordings
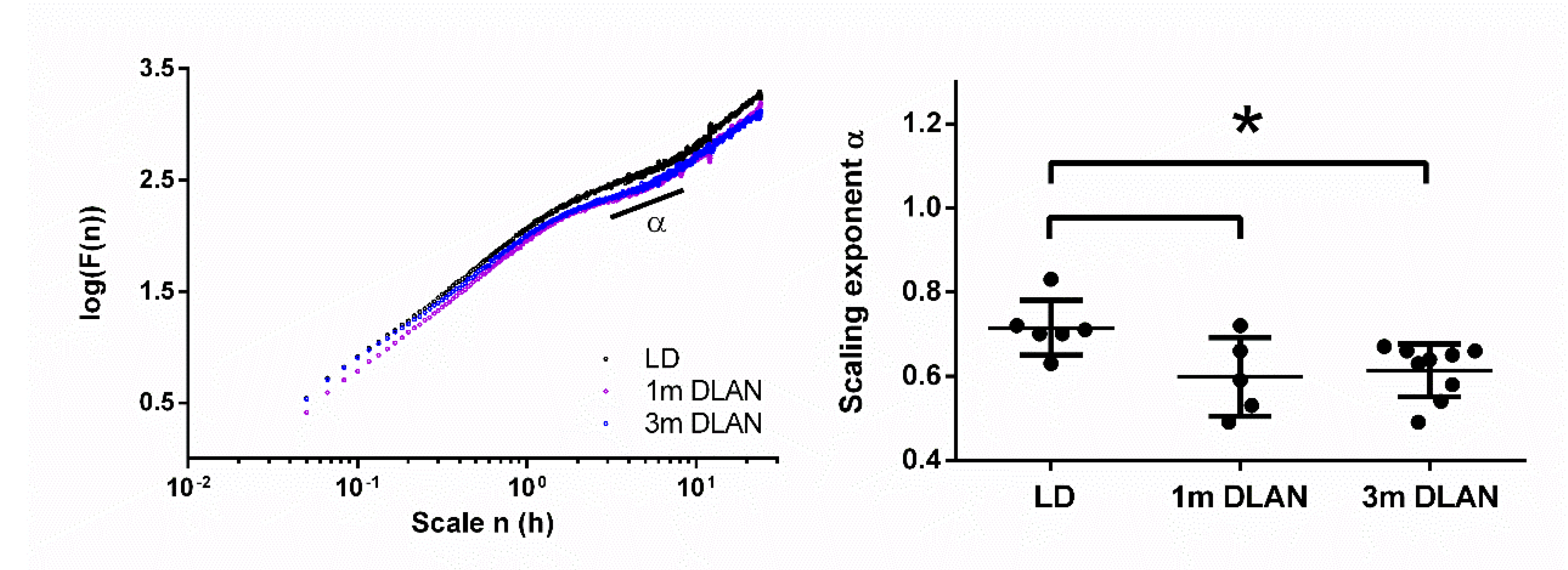
2.4. Assessment of Scale Invariance Using Detrended Fluctuation Analysis (DFA)
2.5. EEG Recordings
2.6. Data Analysis and Statistics
3. Results
3.1. Rest-Ctivity Behavioral Data and DFA Analysis
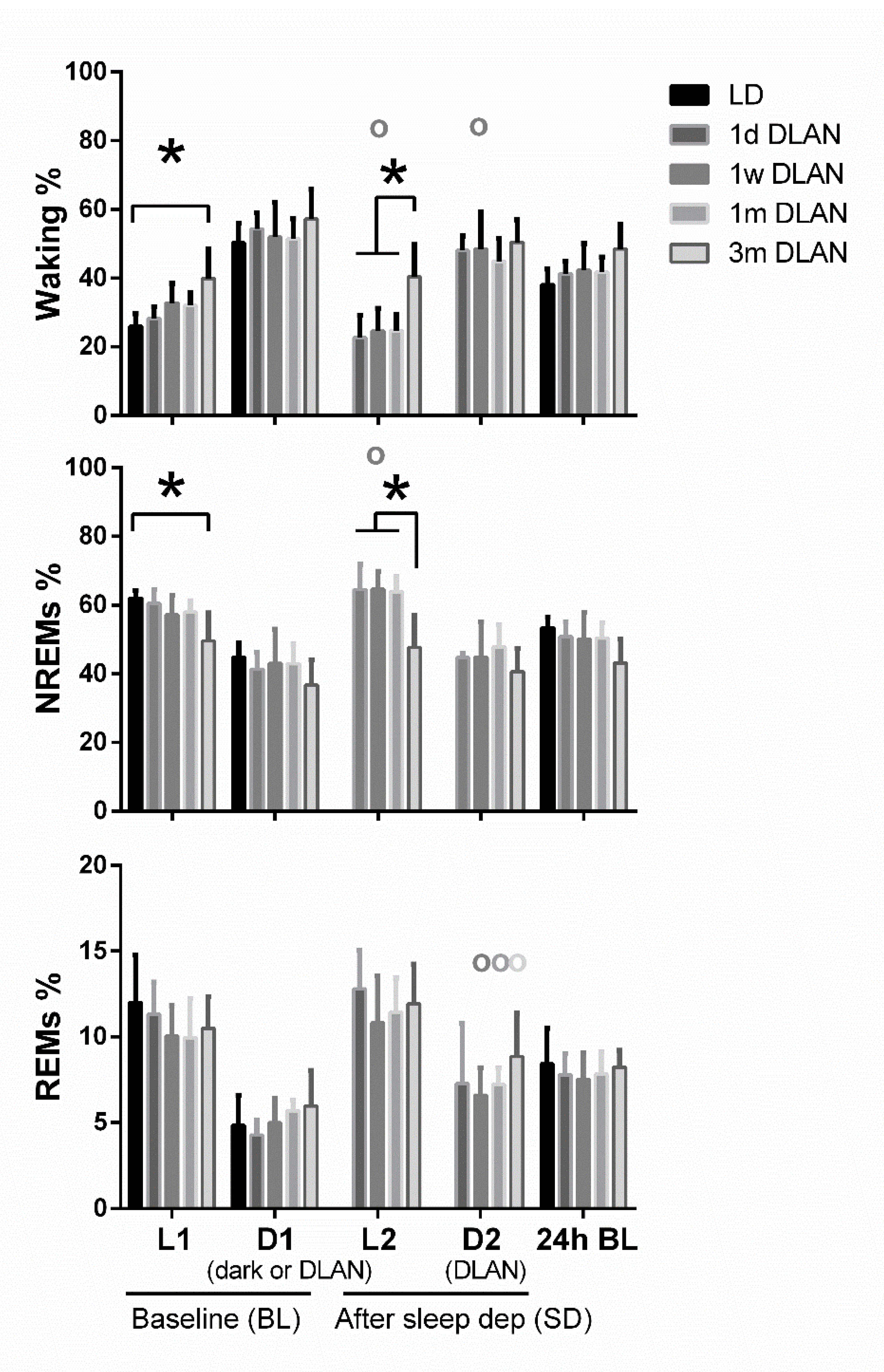
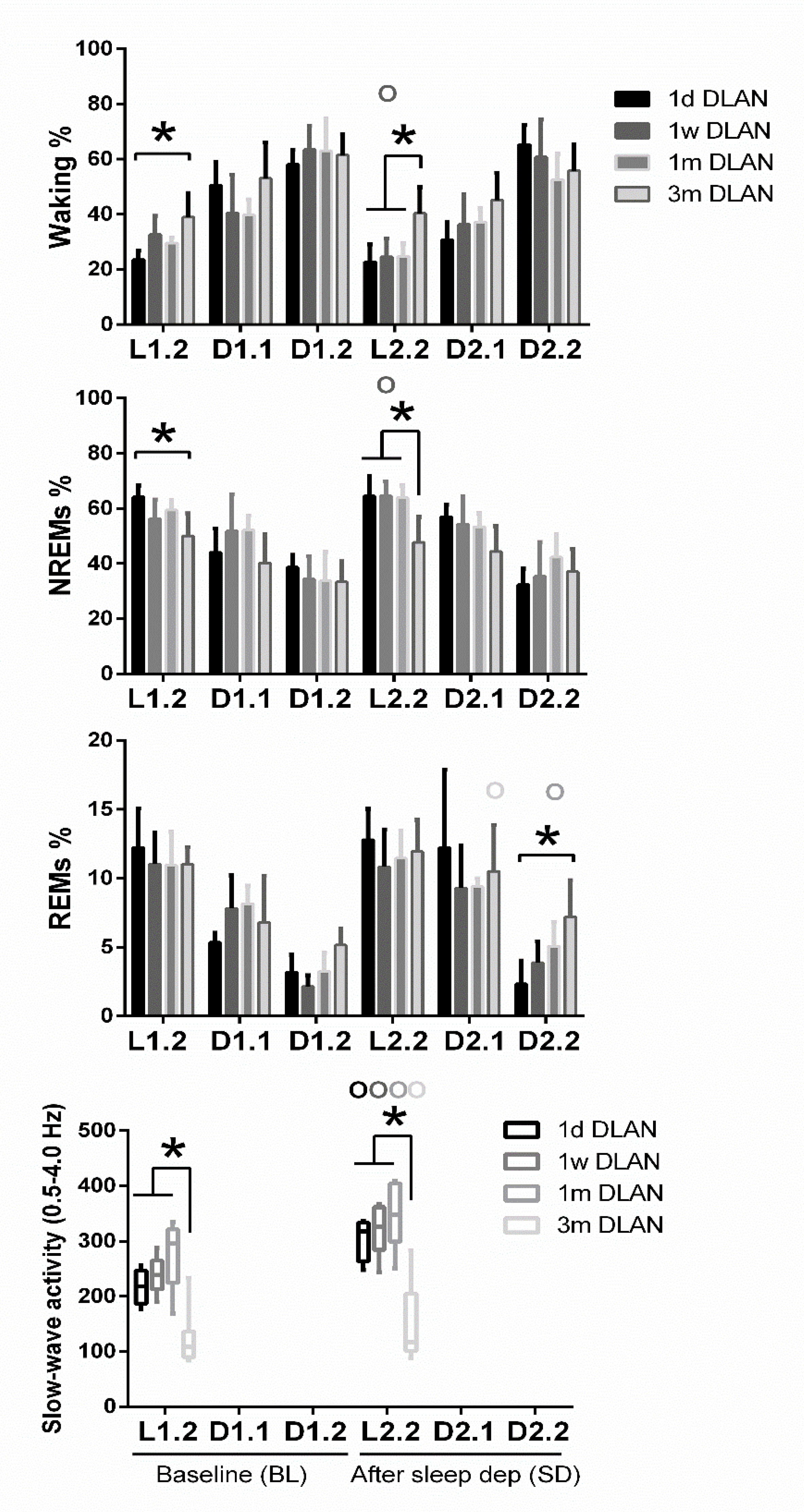
3.2. Sleep Architecture
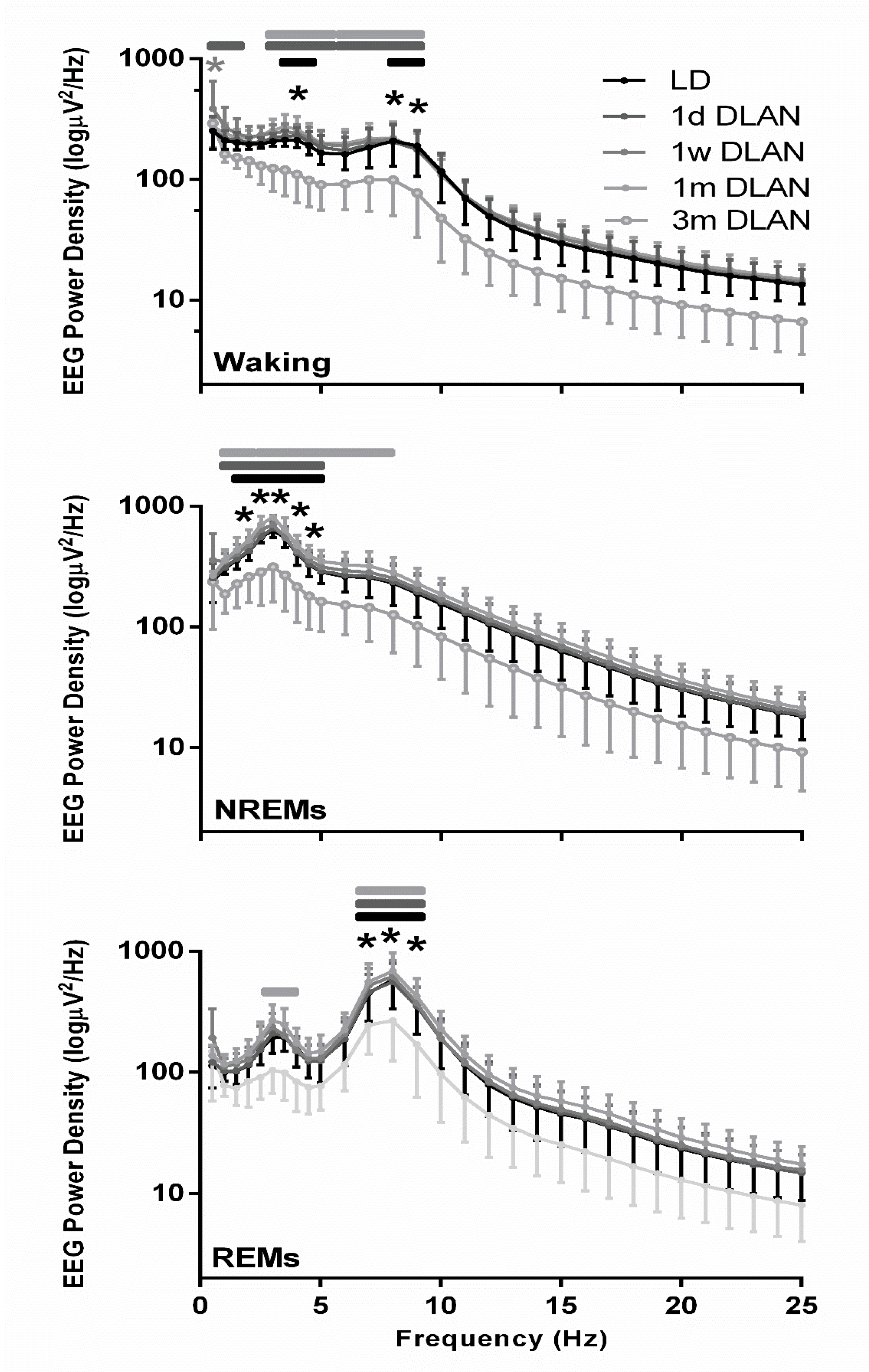
3.3. EEG Power Density
4. Discussion
4.1. Sleep Architecture
4.2. Rest-Activity Behavior and Fractal Patterns
4.3. Eeg Power Density in Waking, Nrem, and Rem Sleep and Slow-Wave Activity (Swa) in Nrem Sleep
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, F.G. Report and Recommendations of IAU Commission 50, Reports on Astronomy; IAU Transactions XVIIA; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1979; pp. 218–222. [Google Scholar]
- Cinzano, P.; Falchi, F.; Elvidge, C. The first World Atlas of the artificial night sky brightness. Mon. Not. R. Astron. Soc. 2001, 328, 689–707. [Google Scholar] [CrossRef]
- Navara, K.J.; Nelson, R.J. The dark side of light at night: Physiological, epidemiological, and ecological consequences. J. Pineal Res. 2007, 43, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Kronauer, R.E.; Czeisler, C.A. Phase-shifting human circadian rhythms: Influence of sleep timing, social contact and light exposure. J. Physiol. 1996, 495, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.A.; Wright, K.P. Influence of Light on Circadian Rhythmicity in Humans I. In Regulation of Sleep and Circadian Rhythms; Turek, F.W., Zee, P.C., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1999; pp. 149–180. [Google Scholar]
- Zeitzer, J.M.; Dijk, D.-J.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; DeBoer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Achermann, P.; Borbely, A.A. Sleep Homeostasis and Models of Sleep Regulation. In Principles and Practice of Sleep Medicine; Kryger, M.H., Roth, T., Dement, W.C., Eds.; Elsevier: Philadelphia, PA, USA, 2017; pp. 377–387. [Google Scholar]
- Cho, J.R.; Joo, E.Y.; Koo, D.L.; Hong, S.B. Let there be no light: The effect of bedside light on sleep quality and background electroencephalographic rhythms. Sleep Med. 2013, 14, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.-H.; Lee, H.-J.; Yoon, H.-K.; Kang, S.-G.; Bok, K.-N.; Jung, K.-Y.; Kim, L.; Lee, E.-I. Exposure to dim artificial light at night increases REM sleep and awakenings in humans. Chronobiol. Int. 2015, 33, 117–123. [Google Scholar] [CrossRef]
- Fonken, L.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.; Nelson, R.J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J. Boil. Rhythm. 2013, 28, 262–271. [Google Scholar] [CrossRef]
- Stenvers, D.J.; Van Dorp, R.; Foppen, E.; Mendoza, J.; Opperhuizen, A.-L.; Fliers, E.; Bisschop, P.H.; Meijer, J.H.; Kalsbeek, A.; DeBoer, T. Dim light at night disturbs the daily sleep-wake cycle in the rat. Sci. Rep. 2016, 6, 35662. [Google Scholar] [CrossRef]
- Le Tallec, T.; Perret, M.; Théry, M. Light Pollution Modifies the Expression of Daily Rhythms and Behavior Patterns in a Nocturnal Primate. PLoS ONE 2013, 8, e79250. [Google Scholar] [CrossRef]
- Borniger, J.C.; Weil, Z.M.; Zhang, N.; Nelson, R.J. Dim Light at Night Does Not Disrupt Timing or Quality of Sleep in Mice. Chronobiol. Int. 2013, 30, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, M.; DeBoer, T. Effects of Chronic Dim-light-at-night Exposure on Sleep in Young and Aged Mice. Neuroscience 2019, 426, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Pittman-Polletta, B.R.; Scheer, F.A.J.L.; Butler, M.P.; Shea, S.A.; Hu, K. The role of the circadian system in fractal neurophysiological control. Boil. Rev. Camb. Philos. 2013, 88, 873–894. [Google Scholar]
- Goldberger, A.L.; Amaral, L.A.N.; Hausdorff, J.M.; Ivanov, P.C.; Peng, C.-K.; Stanley, H.E. Fractal dynamics in physiology: Alterations with disease and aging. Proc. Natl. Acad. Sci. USA 2002, 99 (Suppl. 1), 2466–2472. [Google Scholar] [CrossRef]
- Hardstone, R.; Poil, S.-S.; Schiavone, G.; Jansen, R.; Nikulin, V.V.; Mansvelder, H.D.; Linkenkaer-Hansen, K. Detrended Fluctuation Analysis: A Scale-Free View on Neuronal Oscillations. Front. Physiol. 2012, 3. [Google Scholar] [CrossRef]
- Peng, C.K.; Havlin, S.; Hausdorff, J.M.; Mietus, J.E.; Stanley, H.; Goldberger, A.L. Fractal mechanisms and heart rate dynamics. Long-range correlations and their breakdown with disease. J. Electrocardiol. 1995, 28, 59–65. [Google Scholar] [CrossRef]
- Gu, C.; Coomans, C.; Hu, K.; Scheer, F.A.J.L.; Stanley, H.E.; Meijer, J.H. Lack of exercise leads to significant and reversible loss of scale invariance in both aged and young mice. Proc. Natl. Acad. Sci. USA 2015, 112, 2320–2324. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Cirelli, C.; Tononi, G. Electrophysiological correlates of sleep homeostasis in freely behaving rats. Prog. Brain Res. 2011, 193, 17–38. [Google Scholar]
- Panagiotou, M.; Vyazovskiy, V.V.; Meijer, J.H.; DeBoer, T. Differences in electroencephalographic non-rapid-eye movement sleep slow-wave characteristics between young and old mice. Sci. Rep. 2017, 7, 43656. [Google Scholar] [CrossRef]
- De Boer, T.; Van Diepen, M.H.C.; Ferrari, M.D.; Maagdenberg, A.M.J.M.V.D.; Meijer, J.H. Reduced Sleep and Low Adenosinergic Sensitivity in Cacna1a R192Q Mutant Mice. Sleep 2013, 36, 127–136. [Google Scholar] [CrossRef]
- Panagiotou, M.; DeBoer, T.; Meijer, J.H. Chronic high-caloric diet modifies sleep homeostasis in mice. Eur. J. Neurosci. 2018, 47, 1339–1352. [Google Scholar]
- Panagiotou, M.; Papagiannopoulos, K.; Rohling, J.H.T.; Meijer, J.H.; DeBoer, T. How Old Is Your Brain? Slow-Wave Activity in Non-rapid-eye-movement Sleep as a Marker of Brain Rejuvenation after Long-Term Exercise in Mice. Front. Aging Neurosci. 2018, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Jenni, O.G.; DeBoer, T.; Achermann, P. Development of the 24-h rest-activity pattern in human infants. Infant Behav. Dev. 2006, 29, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-K.; Buldyrev, S.V.; Havlin, S.; Simons, M.; Stanley, H.; Goldberger, A.L. Mosaic organization of DNA nucleotides. Interdiscip. Top. 1994, 49, 1685–1689. [Google Scholar]
- Hu, K.; Ivanov, P.C.; Chen, Z.; Carpena, P.; Stanley, H.E. Effect of trends on detrended fluctuation analysis. Phys. Rev. E 2001, 64, 011114. [Google Scholar] [CrossRef]
- DeBoer, T.; Ruijgrok, G.; Meijer, J.H. Short light-dark cycles affect sleep in mice. Eur. J. Neurosci. 2007, 26, 3518–3523. [Google Scholar]
- Huber, R.; DeBoer, T.; Tobler, I. Effects of sleep deprivation on sleep and sleep EEG in three mouse strains: Empirical data and simulations. Brain Res. 2000, 857, 8–19. [Google Scholar] [CrossRef]
- Tobler, I.; Franken, P.; Alfoldi, P.; Borbély, A.A. Room light impairs sleep in the albino rat. Behav. Brain Res. 1994, 63, 205–211. [Google Scholar] [CrossRef]
- Benca, R.M.; Gilliland, M.A.; Obermeyer, W.H. Effects of Lighting Conditions on Sleep and Wakefulness in Albino Lewis and Pigmented Brown Norway Rats. Sleep 1998, 21, 451–460. [Google Scholar] [CrossRef][Green Version]
- Lancel, M.; Droste, S.K.; Sommer, S.; Reul, J.M.H.M. Influence of regular voluntary exercise on spontaneous and social stress-affected sleep in mice. Eur. J. Neurosci. 2003, 17, 2171–2179. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Ruijgrok, G.; DeBoer, T.; Tobler, I. Running Wheel Accessibility Affects the Regional Electroencephalogram during Sleep in Mice. Cereb. Cortex 2005, 16, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Meijer, J.H.; Shea, S.A.; Tjebbe vanderLeest, H.; Pittman-Polletta, B.; Houben, T.; van Oosterhout, F.; Deboer, T.; Scheer, F.A. Fractal patterns of neural activity exist within the suprachiasmatic nucleus and require extrinsic network interactions. PLoS ONE 2012, 7, e48927. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Van Someren, E.J.; Shea, S.A.; Scheer, F.A. Reduction of scale invariance of activity fluctuations with aging and Alzheimer’s disease: Involvement of the circadian pacemaker. Proc. Natl. Acad. Sci. USA 2009, 106, 2490–2494. [Google Scholar] [CrossRef] [PubMed]
- Vanderwolf, C. The electrocorticogram in relation to physiology and behavior: A new analysis. Electroencephalogr. Clin. Neurophysiol. 1992, 82, 165–175. [Google Scholar] [CrossRef]
- Huber, R.; Boer, D.; Tobler, T. Prion protein: A role in sleep regulation? J. Sleep Res. 1999, 8, 30–36. [Google Scholar] [CrossRef]
- Steriade, M.; Dossi, R.; Nunez, A. Network modulation of a slow intrinsic oscillation of cat thalamocortical neurons implicated in sleep delta waves: Cortically induced synchronization and brainstem cholinergic suppression. J. Neurosci. 1991, 11, 3200–3217. [Google Scholar] [CrossRef]
- Steriade, M.; Nunez, A.; Amzică, F. Intracellular analysis of relations between the slow (<1 Hz) neocortical oscillation and other sleep rhythms of the electroencephalogram. J. Neurosci. 1993, 13, 3266–3283. [Google Scholar]
- De Haan, W.; Mott, K.; van Straaten, E.C.; Scheltens, P.; Stam, C.J. Activity dependent degeneration explains hub vulnerability in Alzheimer’s disease. PLoS Comput. Biol. 2012, 8. [Google Scholar] [CrossRef]
- Stenberg, D. Neuroanatomy and neurochemistry of sleep. Cell. Mol. Life Sci. 2007, 64, 1187–1204. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panagiotou, M.; Rohling, J.H.T.; Deboer, T. Sleep Network Deterioration as a Function of Dim-Light-At-Night Exposure Duration in a Mouse Model. Clocks & Sleep 2020, 2, 308-324. https://doi.org/10.3390/clockssleep2030023
Panagiotou M, Rohling JHT, Deboer T. Sleep Network Deterioration as a Function of Dim-Light-At-Night Exposure Duration in a Mouse Model. Clocks & Sleep. 2020; 2(3):308-324. https://doi.org/10.3390/clockssleep2030023
Chicago/Turabian StylePanagiotou, Maria, Jos H.T. Rohling, and Tom Deboer. 2020. "Sleep Network Deterioration as a Function of Dim-Light-At-Night Exposure Duration in a Mouse Model" Clocks & Sleep 2, no. 3: 308-324. https://doi.org/10.3390/clockssleep2030023
APA StylePanagiotou, M., Rohling, J. H. T., & Deboer, T. (2020). Sleep Network Deterioration as a Function of Dim-Light-At-Night Exposure Duration in a Mouse Model. Clocks & Sleep, 2(3), 308-324. https://doi.org/10.3390/clockssleep2030023