Adverse Neurological Effects of Short-Term Sleep Deprivation in Aging Mice Are Prevented by SS31 Peptide


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
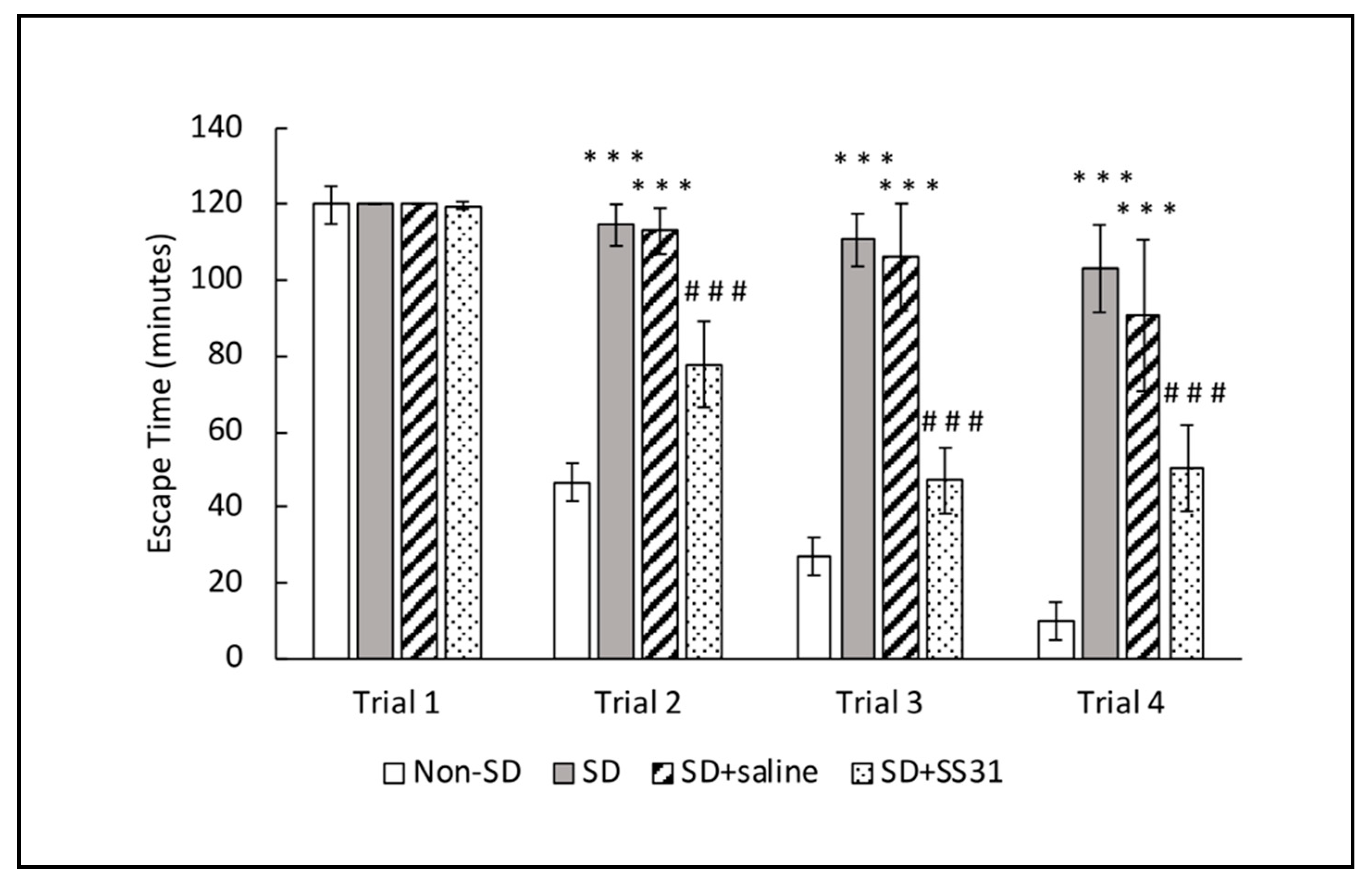
2.1. Sleep-Deprived Learning Impairment Was Prevented in Aging Mice Treated with SS31
2.2. ATP Increased and ROS Decreased in Mitochondria from Brain and Liver of Sleep-Deprived Mice Treated with SS31
2.3. SS31 Restores Hippocampal Regulatory Proteins for Synaptic Plasticity, and Decreases Inflammatory Cytokines in Sleep-Deprived Mice
3. Discussion
4. Methods
4.1. Animals and Treatment Schedule
4.2. Sleep Deprivation Procedure
4.3. Spatial Navigation Box Maze
4.4. Western Blot
4.5. Isolation of Mitochondria
4.6. ROS Assay
4.7. ATP Assay
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maquet, P. The role of sleep in learning and memory. Science 2001, 294, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Abel, T.; Havekes, R.; Saletin, J.M.; Walker, M.P. Sleep, plasticity and memory from molecules to whole-brain networks. Curr. Biol. 2013, 23, R774–R788. [Google Scholar] [CrossRef]
- Stuchlik, A. Dynamic learning and memory, synaptic plasticity and neurogenesis: An update. Front. Behav. Neurosci. 2014, 8, 106. [Google Scholar] [CrossRef]
- Lüscher, C.; Malenka, R.C. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Persp. Biol. 2012, 4, a005710. [Google Scholar] [CrossRef]
- McDermott, C.M.; Hardy, M.N.; Bazan, N.G.; Magee, J.C. Sleep deprivation-induced alterations in excitatory synaptic transmission in the CA1 region of the rat hippocampus. J. Physiol. 2005, 570, 553–565. [Google Scholar] [CrossRef]
- Wadhwa, M.; Kumari, P.; Chauhan, G.; Roy, K.; Alam, S.; Kishore, K.; Ray, K.; Panjwani, U. Sleep deprivation induces spatial memory impairment by altered hippocampus neuroinflammatory responses and glial cells activation in rats. J. Neuroimmunol. 2017, 312, 38–48. [Google Scholar] [CrossRef]
- Andreazza, A.C.; Andersen, M.L.; Alvarenga, T.A.; De Oliveira, M.R.; Armani, F.; Ruiz, F.S.; Giglio, L.; Moreira, J.C.; Kapczinski, F.; Tufik, S. Impairment of the mitochondrial electron transport chain due to sleep deprivation in mice. J. Psychiatr. Res. 2010, 44, 775–780. [Google Scholar] [CrossRef]
- Hasan, S.; Dauvilliers, Y.; Mongrain, V.; Franken, P.; Tafti, M. Age-related changes in sleep in inbred mice are genotype dependent. Neurobiol. Aging 2012, 33, 195.e13–195.e26. [Google Scholar] [CrossRef] [PubMed]
- Pace-Schott, E.F.; Spencer, R.M.C. Sleep-dependent memory consolidation in healthy aging and mild cognitive impairment. Brain Imag. Behav. Neurosci. 2015, 25, 307–330. [Google Scholar] [CrossRef]
- Chen, W.-W.; Zhang, X.; Huang, W.-J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef]
- Barrientos, R.M.; Kitt, M.M.; Watkins, L.R.; Maier, S.F. Neuroinflammation in the normal aging hippocampus. Neuroscience 2015, 309, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Kristofikova, Z.; Sirova, J.; Klaschka, J.; Ovsepian, S.V. Acute and chronic sleep deprivation-related changes in n-methyl-d-aspartate receptor-nitric oxide signalling in the rat cerebral cortex with reference to aging and brain lateralization. Int. J. Mol. Sci. 2019, 20, 3273. [Google Scholar] [CrossRef]
- Zhao, W.; Xu, Z.; Cao, J.; Fu, Q.; Wu, Y.; Zhang, X.; Long, Y.; Zhang, X.; Yang, Y.; Li, Y.-F.; et al. Elamipretide (SS-31) improves mitochondrial dysfunction, synaptic and memory impairment induced by lipopolysaccharide in mice. J. Neuroinflamm. 2019, 16, 230. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, M.; Li, H.; Sun, X.; Hao, S.; Ji, M.; Yang, J.-J.; Li, K. BDNF pathway is involved in the protective effects of SS-31 on isoflurane-induced cognitive deficits in aging mice. Behav. Brain Res. 2016, 305, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Hao, S.; Sun, X.-R.; Zhang, H.; Li, H.; Zhao, H.; Ji, M.-H.; Yang, J.-J.; Li, K. Elamipretide (SS-31) Ameliorates Isoflurane-Induced Long-Term Impairments of Mitochondrial Morphogenesis and Cognition in Developing Rats. Front. Cell. Neurosci. 2017, 11, 119. [Google Scholar] [CrossRef]
- Tarantini, S.; Valcarcel-Ares, N.M.; Yabluchanskiy, A.; Fülöp, G.A.; Hertelendy, P.; Gautam, T.; Farkas, E.; Perz, A.; Rabinovitch, P.S.; Sonntag, W.E.; et al. Treatment with the mitochondrial-targeted antioxidant peptide SS-31 rescues neurovascular coupling responses and cerebrovascular endothelial function and improves cognition in aged mice. Aging Cell 2018, 17, e12731. [Google Scholar] [CrossRef]
- Mukherjee, K.; Lee, A.; Zhu, L.; Darvas, M.; Ladiges, W. Sleep-deprived cognitive impairment in aging mice is alleviated by rapamycin. Aging Pathobiol. Ther. 2019, 1, 5–9. [Google Scholar] [CrossRef]
- Darvas, M.; Mukherjee, K.; Lee, A.; Ladiges, W. A novel one-day learning procedure for mice. Curr. Protoc. Mouse Biol. 2020, 10, e68. [Google Scholar] [CrossRef]
- Silva, A.J. Molecular and cellular cognitive studies of the role of synaptic plasticity in memory. J. Neurobiol. 2002, 54, 224–237. [Google Scholar] [CrossRef]
- Wang, H.-T.; Xu, J.; Lazarovici, P.; Quirion, R.; Zheng, W. cAMP Response Element-Binding Protein (CREB): A Possible Signaling Molecule Link in the Pathophysiology of Schizophrenia. Front. Mol. Neurosci. 2018, 11, 255. [Google Scholar] [CrossRef]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Gaetani, L.; Di Gregorio, M.; Tozzi, A.; Ghiglieri, V.; Calabresi, P.; Di Filippo, M. Hippocampal neuroplasticity and inflammation: Relevance for multiple sclerosis. Mult. Scler. Demyelinating Dis. 2017, 2, 2. [Google Scholar] [CrossRef]
- Nisticò, R.; Salter, E.W.; Nicolas, C.; Feligioni, M.; Mango, D.; Bortolotto, Z.A.; Gressens, P.; Collingridge, G.L.; Peineau, S. Synaptoimmunology - roles in health and disease. Mol. Brain 2017, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Ladiges, W.C.; Knoblaugh, S.E.; Morton, J.F.; Korth, M.J.; Sopher, B.L.; Baskin, C.R.; Macauley, A.; Goodman, A.G.; Leboeuf, R.C.; Katze, M.G. Pancreatic -Cell Failure and Diabetes in Mice With a Deletion Mutation of the Endoplasmic Reticulum Molecular Chaperone Gene P58IPK. Diabetes 2005, 54, 1074–1081. [Google Scholar] [CrossRef]
- Klein, J.A.; Ackerman, S.L. Oxidative stress, cell cycle, and neurodegeneration. J. Clin. Investig. 2003, 111, 785–793. [Google Scholar] [CrossRef]
- Everson, C.A.; Henchen, C.J.; Szabo, A.; Hogg, N. Cell injury and repair resulting from sleep loss and sleep recovery in laboratory rats. Sleep 2014, 37, 1929–1940. [Google Scholar] [CrossRef]
- Periasamy, S.; Hsu, D.-Z.; Fu, Y.-H.; Liu, M.-Y. Sleep deprivation-induced multi-organ injury: Role of oxidative stress and inflammation. EXCLI J. 2015, 14, 672–683. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Boil. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Shigiyama, F.; Kumashiro, N.; Tsuneoka, Y.; Igarashi, H.; Yoshikawa, F.; Kakehi, S.; Funato, H.; Hirose, T. Mechanisms of sleep deprivation-induced hepatic steatosis and insulin resistance in mice. Am. J. Physiol. Metab. 2018, 315, E848–E858. [Google Scholar] [CrossRef]
- Dai, D.-F.; Chiao, Y.A.; Marcinek, D.J.; Szeto, H.H.; Rabinovitch, P. Mitochondrial oxidative stress in aging and healthspan. Longev. Healthspan 2014, 3, 6. [Google Scholar] [CrossRef]
- Zhao, K.; Zhao, G.-M.; Wu, D.; Soong, Y.; Birk, A.V.; Schiller, P.W.; Szeto, H.H. Cell-permeable peptide antioxidants targeted to inner mitochondrial membrane inhibit mitochondrial swelling, oxidative cell death, and reperfusion injury. J. Biol. Chem. 2004, 279, 34682–34690. [Google Scholar] [CrossRef] [PubMed]
- Lash, L.H.; Jones, D.P. Mitochondrial Dysfunction, 1st ed.; Academic Press: New York, NY, USA, 1993. [Google Scholar]
- Yan, L.-J.; Yang, S.-H.; Shu, H.; Prokai, L.; Forster, M.J. Histochemical staining and quantification of dihydrolipoamide dehydrogenase diaphorase activity using blue native PAGE. Electrophoresis 2007, 28, 1036–1045. [Google Scholar] [CrossRef]
- Yan, L.-J.; Thangthaeng, N.; Forster, M.J. Changes in dihydrolipoamide dehydrogenase expression and activity during postnatal development and aging in the rat brain. Mech. Aging Dev. 2008, 129, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Gómez, C.; López-Cepero, J.M.; Boveris, A. Beneficial effects of moderate exercise on mice aging: Survival, behavior, oxidative stress, and mitochondrial electron transfer. Am. J. Physiol. Integr. Comp. Physiol. 2004, 286, R505–R511. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Dou, Y.; Ladiges, W.C. Adverse Neurological Effects of Short-Term Sleep Deprivation in Aging Mice Are Prevented by SS31 Peptide. Clocks & Sleep 2020, 2, 325-333. https://doi.org/10.3390/clockssleep2030024
Wu J, Dou Y, Ladiges WC. Adverse Neurological Effects of Short-Term Sleep Deprivation in Aging Mice Are Prevented by SS31 Peptide. Clocks & Sleep. 2020; 2(3):325-333. https://doi.org/10.3390/clockssleep2030024
Chicago/Turabian StyleWu, Jinzi, Yan Dou, and Warren C. Ladiges. 2020. "Adverse Neurological Effects of Short-Term Sleep Deprivation in Aging Mice Are Prevented by SS31 Peptide" Clocks & Sleep 2, no. 3: 325-333. https://doi.org/10.3390/clockssleep2030024
APA StyleWu, J., Dou, Y., & Ladiges, W. C. (2020). Adverse Neurological Effects of Short-Term Sleep Deprivation in Aging Mice Are Prevented by SS31 Peptide. Clocks & Sleep, 2(3), 325-333. https://doi.org/10.3390/clockssleep2030024