Effects of Sleep and Age on Prospective Memory Consolidation: A Walk in a Virtual Museum


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. General Procedure
2.3. Prospective Memory Task
2.3.1. Virtual Environment
2.3.2. Encoding, Storage, and Recall of Intentions
2.4. Actigraphy Data Analysis
2.5. Statistical Analyses
3. Results
3.1. Effect of Time of Testing on Subjective Sleepiness and PM Performance
3.2. Effects of Sleep and Wakefulness on the Consolidation and Recall of EB and TB Intentions
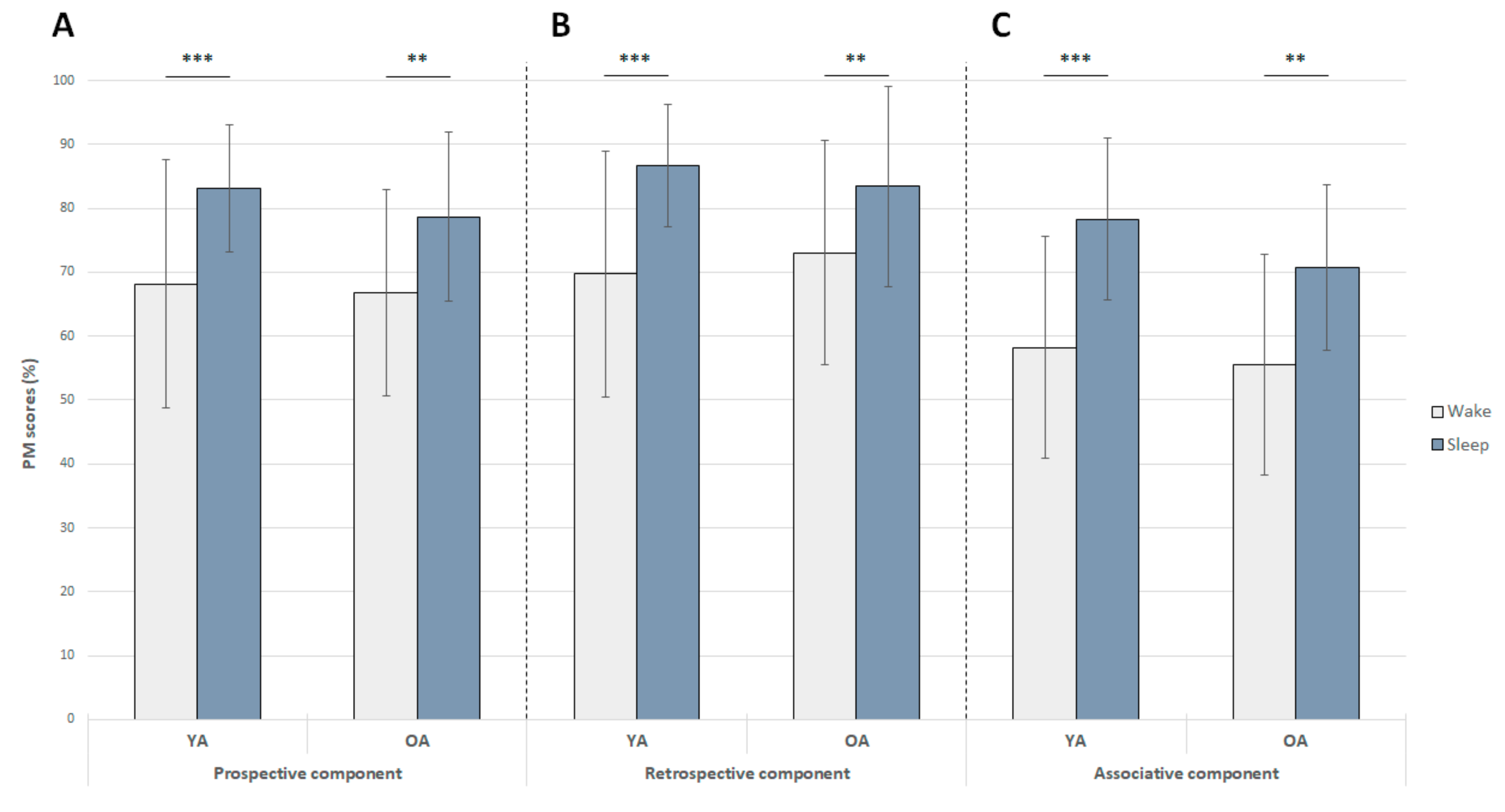
3.3. Effects of Sleep and Wakefulness on the Consolidation and Recall of Prospective, Retrospective, and Associative Components of PM
3.4. Performance on the Ongoing Task
3.5. Associations between Objective Measures of Sleep and PM
4. Discussion
4.1. Effects of Sleep on PM
4.2. Effects of Age on PM
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BDI | Beck Depression Inventory |
| EB | Event-based |
| KSS | Karolinska Sleepiness Scale |
| MMSE | Mini-Mental State Examination |
| PM | Prospective memory |
| PSQI | Pittsburgh Sleep Quality Index |
| REM | Rapid Eye Movement |
| SD SE | Standard deviation Sleep Efficiency |
| SEM SOL | Standard error of the mean Sleep Onset Latency |
| STAI | State-Trait Anxiety Inventory |
| SWS | Slow-wave sleep |
| TB TST WASO | Time-based Total Sleep Time Wake After Sleep Onset |
References
- Rauchs, G.; Desgranges, B.; Foret, J.; Eustache, F. The relationships between memory systems and sleep stages. J. Sleep Res. 2005, 14, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Diekelmann, S.; Born, J. The memory function of sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Rauchs, G.; Feyers, D.; Landeau, B.; Bastin, C.; Luxen, A.; Maquet, P.; Collette, F. Sleep contributes to the strengthening of some memories over others, depending on hippocampal activity at learning. J. Neurosci. 2011, 31, 2563–2568. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, I.; Diekelmann, S.; Molzow, I.; Ayoub, A.; Mölle, M.; Born, J. Sleep Selectively Enhances Memory Expected to Be of Future Relevance. J. Neurosci. 2011, 31, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, E.V.; Thielen, J.-W.; Takashima, A.; Barth, M.; Fernández, G. Sleep Supports Selective Retention of Associative Memories Based on Relevance for Future Utilization. PLoS ONE 2012, 7, e43426. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Born, J. Anticipated reward enhances offline learning during sleep. J. Exp. Psychol. Learn. Mem. Cogn. 2009, 35, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, M.A.; Einstein, G.O. Prospective Memory: An Overview and Synthesis of an Emerging Field; Sage Publications, Inc.: Thousand Oaks, CA, USA, 2007; ISBN 978-1-4129-2469-6. [Google Scholar]
- Scullin, M.K.; McDaniel, M.A. Remembering to execute a goal: Sleep on it. Psychol. Sci. 2010, 21, 1028–1035. [Google Scholar] [CrossRef]
- Diekelmann, S.; Wilhelm, I.; Wagner, U.; Born, J. Sleep to implement an intention. Sleep 2013, 36, 149–153. [Google Scholar] [CrossRef]
- Diekelmann, S.; Wilhelm, I.; Wagner, U.; Born, J. Sleep Improves Prospective Remembering by Facilitating Spontaneous-Associative Retrieval Processes. PLoS ONE 2013, 8, e77621. [Google Scholar] [CrossRef]
- Grundgeiger, T.; Bayen, U.J.; Horn, S.S. Effects of sleep deprivation on prospective memory. Memory 2014, 22, 679–686. [Google Scholar] [CrossRef]
- Esposito, M.J.; Occhionero, M.; Cicogna, P. Sleep Deprivation and Time-Based Prospective Memory. Sleep 2015, 38, 1823–1826. [Google Scholar] [CrossRef] [PubMed]
- Barner, C.; Seibold, M.; Born, J.; Diekelmann, S. Consolidation of Prospective Memory: Effects of Sleep on Completed and Reinstated Intentions. Front. Psychol. 2017, 7, 2025. [Google Scholar] [CrossRef] [PubMed]
- Barner, C.; Altgassen, M.; Born, J.; Diekelmann, S. Effects of sleep on the realization of complex plans. J. Sleep Res. 2019, 28, e12655. [Google Scholar] [CrossRef] [PubMed]
- Leong, R.L.; Cheng, G.H.-L.; Chee, M.W.; Lo, J.C. The effects of sleep on prospective memory: A systematic review and meta-analysis. Sleep Med. Rev. 2019, 47, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Einstein, G.O.; McDaniel, M.A. Normal aging and prospective memory. J. Exp. Psychol. Learn. Mem. Cogn. 1990, 16, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Guynn, M.J.; McDaniel, M.A.; Einstein, G.O. Prospective memory: When reminders fail. Mem. Cogn. 1998, 26, 287–298. [Google Scholar] [CrossRef]
- McDaniel, M.A.; Robinson-Riegler, B.; Einstein, G.O. Prospective remembering: Perceptually driven or conceptually driven processes? Mem. Cogn. 1998, 26, 121–134. [Google Scholar] [CrossRef]
- McDaniel, M.A.; Guynn, M.J.; Einstein, G.O.; Breneiser, J. Cue-Focused and Reflexive-Associative Processes in Prospective Memory Retrieval. J. Exp. Psychol. Learn. Mem. Cogn. 2004, 30, 605–614. [Google Scholar] [CrossRef]
- Marsh, R.L.; Hicks, J.L.; Cook, G.I.; Hansen, J.S.; Pallos, A.L. Interference to ongoing activities covaries with the characteristics of an event-based intention. J. Exp. Psychol. Learn. Mem. Cogn. 2003, 29, 861–870. [Google Scholar] [CrossRef]
- Gonneaud, J.; Kalpouzos, G.; Bon, L.; Viader, F.; Eustache, F.; Desgranges, B. Distinct and shared cognitive functions mediate event- and time-based prospective memory impairment in normal ageing. Memory 2011, 19, 360–377. [Google Scholar]
- McDaniel, M.A.; Einstein, G.O. The neuropsychology of prospective memory in normal aging: A componential approach. Neuropsychologia 2011, 49, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Lecouvey, G.; Gonneaud, J.; Piolino, P.; Madeleine, S.; Orriols, E.; Fleury, P.; Eustache, F.; Desgranges, B. Is binding decline the main source of the ageing effect on prospective memory? A ride in a virtual town. Socioaffect. Neurosci. Psychol. 2017, 7, 1304610. [Google Scholar] [CrossRef] [PubMed]
- Uttl, B. Transparent Meta-Analysis of Prospective Memory and Aging. PLoS ONE 2008, 3, e1568. [Google Scholar] [CrossRef] [PubMed]
- Aberle, I.; Rendell, P.G.; Rose, N.S.; McDaniel, M.A.; Kliegel, M. The Age Prospective Memory Paradox: Young Adults May Not Give Their Best Outside of the Lab. Dev. Psychol. 2010, 46, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Schnitzspahn, K.M.; Ihle, A.; Henry, J.D.; Rendell, P.G.; Kliegel, M. The age-prospective memory-paradox: An exploration of possible mechanisms. Int. Psychogeriatr. 2011, 23, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Niedźwieńska, A.; Barzykowski, K. The age prospective memory paradox within the same sample in time-based and event-based tasks. Aging Neuropsychol. Cogn. 2012, 19, 58–83. [Google Scholar] [CrossRef] [PubMed]
- Kvavilashvili, L.; Cockburn, J.; Kornbrot, D.E. Prospective memory and ageing paradox with event-based tasks: A study of young, young-old, and old-old participants. Q. J. Exp. Psychol. 2013, 66, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Phillips, L.H.; Henry, J.D.; Martin, M. Adult aging and prospective memory: The importance of ecological validity. In Prospective Memory: Cognitive, Neuroscience, Developmental, and Applied Perspectives; Taylor & Francis Group/Lawrence Erlbaum Associates: New York, NY, USA, 2008; pp. 161–185. [Google Scholar]
- Knight, R.G.; Titov, N. Use of Virtual Reality Tasks to Assess Prospective Memory: Applicability and Evidence. Brain Impair. 2009, 10, 3–13. [Google Scholar] [CrossRef]
- Backhaus, J.; Born, J.; Hoeckesfeld, R.; Fokuhl, S.; Hohagen, F.; Junghanns, K. Midlife decline in declarative memory consolidation is correlated with a decline in slow wave sleep. Learn. Mem. 2007, 14, 336–341. [Google Scholar] [CrossRef]
- Cherdieu, M.; Reynaud, E.; Uhlrich, J.; Versace, R.; Mazza, S. Does age worsen sleep-dependent memory consolidation? J. Sleep Res. 2014, 23, 53–60. [Google Scholar] [CrossRef]
- Mander, B.A.; Rao, V.; Lu, B.; Saletin, J.M.; Lindquist, J.R.; Ancoli-Israel, S.; Jagust, W.; Walker, M.P. Prefrontal atrophy, disrupted NREM slow waves and impaired hippocampal-dependent memory in aging. Nat. Neurosci. 2013, 16, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Rao, V.; Lu, B.; Saletin, J.M.; Ancoli-Israel, S.; Jagust, W.J.; Walker, M.P. Impaired prefrontal sleep spindle regulation of hippocampal-dependent learning in older adults. Cereb. Cortex 2014, 24, 3301–3309. [Google Scholar] [CrossRef] [PubMed]
- Varga, A.W.; Ducca, E.L.; Kishi, A.; Fischer, E.; Parekh, A.; Koushyk, V.; Yau, P.L.; Gumb, T.; Leibert, D.P.; Wohlleber, M.E.; et al. Effects of Aging on Slow Wave Sleep Dynamics and Human Spatial Navigational Memory Consolidation. Neurobiol. Aging 2016, 42, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and human aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.K.; Baran, B.; Pace-Schott, E.F.; Ivry, R.B.; Spencer, R.M.C. Sleep modulates word-pair learning but not motor sequence learning in healthy older adults. Neurobiol. Aging 2012, 33, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Aly, M.; Moscovitch, M. The effects of sleep on episodic memory in older and younger adults. Memery 2010, 18, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Cavuoto, M.G.; Ong, B.; Pike, K.E.; Nicholas, C.L.; Bei, B.; Kinsella, G.J. Objective but not subjective sleep predicts memory in community-dwelling older adults. J. Sleep Res. 2016, 25, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Tassi, P.; Muzet, A. Sleep inertia. Sleep Med. Rev. 2000, 4, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Fine, L.; Weinborn, M.; Ng, A.; Loft, S.; Li, Y.R.; Hodgson, E.; Parker, D.; Rainey Smith, S.; Sohrabi, H.R.; Brown, B.; et al. Sleep disruption explains age-related prospective memory deficits: Implications for cognitive aging and intervention. Neuropsychol. Dev. Cogn. B Aging Neuropsychol. Cogn. 2019, 26, 621–636. [Google Scholar] [CrossRef]
- Scullin, M.K.; Gao, C.; Fillmore, P.; Roberts, R.L.; Pruett, N.; Bliwise, D.L. REM Sleep Mediates Age-Related Decline in Prospective Memory Consolidation. Sleep 2019, 42. [Google Scholar] [CrossRef]
- Umeda, S.; Kurosaki, Y.; Terasawa, Y.; Kato, M.; Miyahara, Y. Deficits in prospective memory following damage to the prefrontal cortex. Neuropsychol. 2011, 49, 2178–2184. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Carlesimo, G.A.; Caltagirone, C. Prospective memory functioning: A new area of investigation in the clinical neuropsychology and rehabilitation of Parkinson’s disease and mild cognitive impairment. Review of evidence. Neurol. Sci. 2012, 33, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Einstein, G.O.; McDaniel, M.A.; Richardson, S.L.; Guynn, M.J.; Cunfer, A.R. Aging and prospective memory: Examining the influences of self-initiated retrieval processes. J. Exp. Psychol. Learn. Mem. Cogn. 1995, 21, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Hertzog, C.; Kidder, D.P.; Morrell, R.W.; Mayhorn, C.B. Effect of age on event-based and time-based prospective memory. Psychol. Aging 1997, 12, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Gonneaud, J.; Lecouvey, G.; Groussard, M.; Gaubert, M.; Landeau, B.; Mézenge, F.; de La Sayette, V.; Eustache, F.; Desgranges, B.; Rauchs, G. Functional dedifferentiation and reduced task-related deactivations underlie the age-related decline of prospective memory. Brain Imaging Behav. 2017, 11, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Hainselin, M.; Quinette, P.; Desgranges, B.; Martinaud, O.; Hannequin, D.; De La Sayette, V.; Viader, F.; Eustache, F. Can We Remember Future Actions yet Forget the Last Two Minutes? Study in Transient Global Amnesia. J. Cogn. Neurosci. 2011, 23, 4138–4149. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Durrant, S.J.; Cairney, S.A.; McDermott, C.; Lewis, P.A. Schema-conformant memories are preferentially consolidated during REM sleep. Neurobiol. Learn. Mem. 2015, 122, 41–50. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Spielberger, C.; Sydenham, S. State-Trait Anxiety Inventory and State-Trait Anger Expression Inventory; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1994; pp. 292–321. [Google Scholar]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An Inventory for Measuring Depression. Arch. Gen. Psychiatry 1961, 4, 561. [Google Scholar] [CrossRef]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. Mini-mental state: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Åkerstedt, T.; Gillberg, M. Subjective and Objective Sleepiness in the Active Individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.W.; Johns, M.W.; Lancaster, R.; Raptopoulos, P.; Angelopoulos, N.; Priest, R.G. The St. Mary’s Hospital sleep questionnaire: A study of reliability. Sleep 1981, 4, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Peigneux, P.; Muto, V.; Schenkel, M.; Knoblauch, V.; Münch, M.; De Quervain, D.J.-F.; Wirz-Justice, A.; Cajochen, C. Encoding Difficulty Promotes Postlearning Changes in Sleep Spindle Activity during Napping. J. Neurosci. 2006, 26, 8976–8982. [Google Scholar] [CrossRef]
- Drosopoulos, S.; Schulze, C.; Fischer, S.; Born, J. Sleep’s function in the spontaneous recovery and consolidation of memories. J. Exp. Psychol. Gen. 2007, 136, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Plihal, W.; Born, J. Effects of Early and Late Nocturnal Sleep on Declarative and Procedural Memory. J. Cogn. Neurosci. 1997, 9, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Tucker, M.; Hirota, Y.; Wamsley, E.; Lau, H.; Chaklader, A.; Fishbein, W. A daytime nap containing solely non-REM sleep enhances declarative but not procedural memory. Neurobiol. Learn. Mem. 2006, 86, 241–247. [Google Scholar] [CrossRef]
- Benson, K.; Feinberg, I. Sleep and Memory: Retention 8 and 24 Hours After Initial Learning. Psychophysiology 1975, 12, 192–195. [Google Scholar] [CrossRef]
- Benson, K.; Feinberg, I. The Beneficial Effect of Sleep in an Extended Jenkins and Dallenbach Paradigm. Psychophysiology 1977, 14, 375–384. [Google Scholar] [CrossRef]
- Diekelmann, S.; Wilhelm, I.; Born, J. The whats and whens of sleep-dependent memory consolidation. Sleep Med. Rev. 2009, 13, 309–321. [Google Scholar] [CrossRef]
- Payne, J.D.; Tucker, M.A.; Ellenbogen, J.M.; Wamsley, E.J.; Walker, M.P.; Schacter, D.L.; Stickgold, R. Memory for Semantically Related and Unrelated Declarative Information: The Benefit of Sleep, the Cost of Wake. PLoS ONE 2012, 7, e33079. [Google Scholar] [CrossRef] [PubMed]
- Tse, D.; Langston, R.F.; Kakeyama, M.; Bethus, I.; Spooner, P.A.; Wood, E.R.; Witter, M.P.; Morris, R.G.M. Schemas and memory consolidation. Science 2007, 316, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Van Kesteren, M.T.; Ruiter, D.J.; Fernández, G.; Henson, R.N. How schema and novelty augment memory formation. Trends Neurosci. 2012, 35, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Gilboa, A.; Marlatte, H. Neurobiology of Schemas and Schema-Mediated Memory. Trends Cogn. Sci. 2017, 21, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Landmann, N.; Kuhn, M.; Piosczyk, H.; Feige, B.; Baglioni, C.; Spiegelhalder, K.; Frase, L.; Riemann, D.; Sterr, A.; Nissen, C. The reorganisation of memory during sleep. Sleep Med. Rev. 2014, 18, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.A.; Durrant, S.J. Overlapping memory replay during sleep builds cognitive schemata. Trends Cogn. Sci. 2011, 15, 343–351. [Google Scholar] [CrossRef]
- Hennies, N.; Lambon Ralph, M.A.; Kempkes, M.; Cousins, J.N.; Lewis, P.A. Sleep spindle density predicts the effect of prior knowledge of memory consolidation. J. Neurosci. 2016, 36, 3799–3810. [Google Scholar] [CrossRef]
- Tamminen, J.; Payne, J.D.; Stickgold, R.; Wamsley, E.J.; Gaskell, M.G. Sleep spindle activity is associated with the integration of new memories and existing knowledge. J. Neurosci. 2010, 30, 14356–14360. [Google Scholar] [CrossRef]
- Fogel, S.; Martin, N.; LaFortune, M.; Barakat, M.; Debas, K.; LaVenture, S.; Latreille, V.; Gagnon, J.-F.; Doyon, J.; Carrier, J.; et al. NREM Sleep Oscillations and Brain Plasticity in Aging. Front. Neurol. 2012, 3, 176. [Google Scholar] [CrossRef]
- Nyberg, L. Functional brain imaging of episodic memory decline in ageing. J. Intern. Med. 2017, 281, 65–74. [Google Scholar] [CrossRef]
- Umeda, S.; Nagumo, Y.; Kato, M. Dissociative Contributions of Medial Temporal and Frontal Regions to Prospective Remembering. Rev. Neurosci. 2006, 17, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Kalpouzos, G.; Chételat, G.; Baron, J.-C.; Landeau, B.; Mevel, K.; Godeau, C.; Barre, L.; Constans, J.-M.; Viader, F.; Eustache, F.; et al. Voxel-based mapping of brain gray matter volume and glucose metabolism profiles in normal aging. Neurobiol. Aging 2009, 30, 112–124. [Google Scholar] [CrossRef] [PubMed]
- La Joie, R.; Fouquet, M.; Mezenge, F.; Landeau, B.; Villain, N.; Mevel, K.; Pélerin, A.; Eustache, F.; Desgranges, B.; Chételat, G. Differential effect of age on hippocampal subfields assessed using a new high-resolution 3T MR sequence. NeuroImage 2010, 53, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.J.; Johnson, M.K.; Raye, C.L.; Mather, M.; D’Esposito, M. Aging and reflective processes of working memory: Binding and test load deficits. Psychol. Aging 2000, 15, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.J.; Johnson, M.K.; Raye, C.L.; D’Esposito, M. fMRI evidence of age-related hippocampal dysfunction in feature binding in working memory. Cogn. Brain Res. 2000, 10, 197–206. [Google Scholar] [CrossRef]
- Plancher, G.; Gyselinck, V.; Nicolas, S.; Piolino, P. Age effect on components of episodic memory and feature binding: A virtual reality study. Neuropsychology 2010, 24, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Stern, Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef]
- Zimmerman, M.E.; Bigal, M.E.; Katz, M.J.; Brickman, A.M.; Lipton, R.B. Sleep Onset/Maintenance Difficulties and Cognitive Function in Nondemented Older Adults: The Role of Cognitive Reserve. J. Int. Neuropsychol. Soc. 2012, 18, 461–470. [Google Scholar] [CrossRef]
- Alchanatis, M.; Zias, N.; Deligiorgis, N.; Amfilochiou, A.; Dionellis, G.; Orphanidou, D. Sleep apnea-related cognitive deficits and intelligence: an implication of cognitive reserve theory. J. Sleep Res. 2005, 14, 69–75. [Google Scholar] [CrossRef]
- Petit, D.; Gagnon, J.-F.; Fantini, M.L.; Ferini-Strambi, L.; Montplaisir, J. Sleep and quantitative EEG in neurodegenerative disorders. J. Psychosom. Res. 2004, 56, 487–496. [Google Scholar] [CrossRef]
- Rendell, P.G.; Craik, F.I.M. Virtual week and actual week: Age-related differences in prospective memory. Appl. Cogn. Psychol. 2000, 14, S43–S62. [Google Scholar] [CrossRef]
- Henry, J.D.; MacLeod, M.S.; Phillips, L.H.; Crawford, J.R. A Meta-Analytic Review of Prospective Memory and Aging. Psychol. Aging 2004, 19, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Brandimonte, M.A.; Ferrante, D.; Bianco, C.; Villani, M.G. Memory for pro-social intentions: When competing motives collide. Cognition 2010, 114, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Penningroth, S.L.; Scott, W.D.; Freuen, M. Social motivation in prospective memory: Higher importance ratings and reported performance rates for social tasks. Can. J. Exp. Psychol. Rev. Can. Psychol. Exp. 2011, 65, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Himmer, L.; Schönauer, M.; Heib, D.P.J.; Schabus, M.; Gais, S. Rehearsal initiates systems memory consolidation, sleep makes it last. Sci. Adv. 2019, 5, eaav1695. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Adults | Older Adults | p Value | |
|---|---|---|---|
| M ± SD | M ± SD | ||
| Age (years) | 21.8 ± 2.8 | 62.7 ± 4.7 | <0.001 *** |
| Sex ratio (M/F) | 13 / 15 | 14 / 15 | 0.69 |
| Education (years) | 13.6 ± 1.7 | 14.2 ± 2.7 | 0.35 |
| STAI-A | 28.9 ± 5.0 | 31.9 ± 9.0 | 0.13 |
| STAI-B | 34.5 ± 5.2 | 34.9 ± 8.1 | 0.86 |
| BDI | 1.7 ± 1.4 | 2.0 ± 2.0 | 0.51 |
| MMSE | - | 29.5 ± 0.6 | - |
| PSQI | 2.8 ± 1.4 | 2.7 ± 1.8 | 0.78 |
| Sleep parameters | |||
| TST (min) | 377.6 ± 92.9 | 370.3 ± 63.1 | 0.74 |
| SOL (min) | 2.3 ± 6.7 | 3.3 ± 5.5 | 0.57 |
| WASO (min) | 66.4 ± 36.8 | 69.2 ± 26.2 | 0.75 |
| SE (%) | 84.3 ± 7.9 | 82.7 ± 9.5 | 0.53 |
| Sleep Fragmentation | 27.6 ± 10.8 | 32.8 ± 19.3 | 0.25 |
| POINTS | PM INTENTIONS | ||||
|---|---|---|---|---|---|
| EB-link intentions “At the media library, rent a DVD on the Normandy landings.” | EB-nolink intentions “At the restaurant, buy the movie program.” | TB intentions “At 05:58 p.m., take a picture of the tank.” | |||
| COMPONENTS | Prospective | 2 | Recall occured when entering the media library for the first time | Recall occured when entering the restaurant for the first time | Recall occured at the exact time |
| 1 | Recall occured when entering the media library for the second time | Recall occured when entering the restaurant for the second time | Recall occured within 1 minute of the target time | ||
| 0.5 | Recall occured when entering the media library for the third or more time | Recall occured when entering the restaurant for the third or more time | Recall occured within 2 minutes of the target time | ||
| 0 | The participant did not remember that something had to be done at the media library | The participant did not remember that something had to be done at the restaurant | Recall occured more than 3 minutes before or after the target time/The participant did not remember that something had to be done at 05:58 p.m. | ||
| Retrospective | 2 | Action perfectly recalled | Action perfectly recalled | Action perfectly recalled | |
| 1 | “I have to rent a DVD on World War II” | “I have to buy the program of the exhibition” | “I have to take a picture of the plane” | ||
| 0.5 | “I have to rent a DVD…” | “I have to buy a program…” | “I have to take a picture…” | ||
| 0 | The action was not recalled | The action was not recalled | The action was not recalled | ||
| Associative | 2 | Prospective and retrospective components recalled together | Prospective and retrospective components recalled together | Prospective and retrospective components recalled together | |
| 0 | Prospective and retrospective components recalled separately | Prospective and retrospective components recalled separately | Prospective and retrospective components recalled separately | ||
| Young Adults | Older Adults | |
|---|---|---|
| EB-link intentions | 7.7 ± 2.5 | 9.0 ± 2.7 |
| EB-nolink intentions | 9.8 ± 2.7 | 13.7 ± 2.8 *** |
| TB intentions | 11.8 ± 2.6 | 14.6 ± 2.5 ** |
| Morning | Evening | |||
|---|---|---|---|---|
| Young adults | Ongoing task (d’) | 2.4 ± 0.5 | 2.3 ± 0.9 | |
| PM intentions | Global PM scores (%) | 79.4 ± 9.2 | 78.7 ± 8.5 | |
| EB-link (%) | 84.5 ± 10.7 | 84.7 ± 11.3 | ||
| EB-nolink (%) | 69.5 ± 13.6 | 73.8 ± 12.1 | ||
| TB (%) | 84.1 ± 13.4 | 77.7 ± 14.7 | ||
| PM components | Prospective (%) | 73.4 ± 11.7 | 72.9 ± 7.4 | |
| Retrospective (%) | 89.1 ± 6.0 | 93.6 ± 6.4 | ||
| Associative (%) | 75.6 ± 12.5 | 69.6 ± 14.5 | ||
| Older adults | Ongoing task (d’) | 2.0 ± 0.7 | 2.3 ± 0.5 | |
| PM intentions | Global PM scores (%) | 76.8 ± 11.4 | 79.4 ± 8.6 | |
| EB-link (%) | 86.2 ± 9.6 | 84.1 ± 9.1 | ||
| EB-nolink (%) | 71.6 ± 17.7 | 73.4 ± 12.5 | ||
| TB (%) | 72.6 ± 14.4 | 80.8 ± 17.6 | ||
| PM components | Prospective (%) | 67.3 ± 13.8 | 73.6 ± 10.8 | |
| Retrospective (%) | 91.1 ± 7.2 | 93.6 ± 6.9 | ||
| Associative (%) | 72.0 ± 15.6 | 71.2 ± 11.7 |
| Wake | Sleep | |||
|---|---|---|---|---|
| Young adults | Ongoing task (d’) | 2.7 ± 0.7 # | 2.5 ± 0.8 | |
| PM intentions | Global PM scores (%) | 65.4 ± 15.9 | 82.7 ± 8.4 *** | |
| EB-link (%) | 67.9 ± 19.5 | 86.8 ± 9.9 *** | ||
| EB-nolink (%) | 56.8 ± 19.2 | 75.1 ± 9.5 *** | ||
| TB (%) | 71.4 ± 17.4 | 86.2 ± 12.7 ** | ||
| PM components | Prospective (%) | 68.2 ± 15.7 | 83.2 ± 8.5 *** | |
| Retrospective (%) | 69.7 ± 15.0 | 86.7 ± 6.8 *** | ||
| Associative (%) | 58.2 ± 18.6 | 78.3 ± 12.8 *** | ||
| Older adults | Ongoing task (d’) | 2.1 ± 0.8 | 2.2 ± 0.5 | |
| PM intentions | Global PM scores (%) | 65.1 ± 10.6 | 77.6 ± 10.6 *** | |
| EB-link (%) | 67.8 ± 16.2 | 82.4 ± 13.2 ** | ||
| EB-nolink (%) | 61.0 ± 17.5 | 69.1 ± 15.7 | ||
| TB (%) | 66.6 ± 17.2 | 81.3 ± 13.0 ** | ||
| PM components | Prospective (%) | 66.8 ± 8.6 | 78.6 ± 11.4 ** | |
| Retrospective (%) | 73.1 ± 13.2 | 83.4 ± 10.9 ** | ||
| Associative (%) | 55.6 ± 14.2 | 70.7 ± 15.3 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehel, S.; Legrand, N.; Lecouvey, G.; Laniepce, A.; Bertran, F.; Fleury, P.; Madeleine, S.; Eustache, F.; Desgranges, B.; Rauchs, G. Effects of Sleep and Age on Prospective Memory Consolidation: A Walk in a Virtual Museum. Clocks & Sleep 2019, 1, 332-351. https://doi.org/10.3390/clockssleep1030028
Rehel S, Legrand N, Lecouvey G, Laniepce A, Bertran F, Fleury P, Madeleine S, Eustache F, Desgranges B, Rauchs G. Effects of Sleep and Age on Prospective Memory Consolidation: A Walk in a Virtual Museum. Clocks & Sleep. 2019; 1(3):332-351. https://doi.org/10.3390/clockssleep1030028
Chicago/Turabian StyleRehel, Stéphane, Nicolas Legrand, Grégory Lecouvey, Alice Laniepce, Françoise Bertran, Philippe Fleury, Sophie Madeleine, Francis Eustache, Béatrice Desgranges, and Géraldine Rauchs. 2019. "Effects of Sleep and Age on Prospective Memory Consolidation: A Walk in a Virtual Museum" Clocks & Sleep 1, no. 3: 332-351. https://doi.org/10.3390/clockssleep1030028
APA StyleRehel, S., Legrand, N., Lecouvey, G., Laniepce, A., Bertran, F., Fleury, P., Madeleine, S., Eustache, F., Desgranges, B., & Rauchs, G. (2019). Effects of Sleep and Age on Prospective Memory Consolidation: A Walk in a Virtual Museum. Clocks & Sleep, 1(3), 332-351. https://doi.org/10.3390/clockssleep1030028