Chronotype-Dependent Changes in Sleep Habits Associated with Dim Light Melatonin Onset in the Antarctic Summer

 ,
,
Abstract
1. Introduction
2. Study Population and Methods
2.1. Participants
2.2. Questionnaires
2.3. Sleep Logs (SL)
2.4. Light Recording
2.5. Melatonin Measurements
2.6. Statistical Analysis
3. Results
3.1. Light Exposure
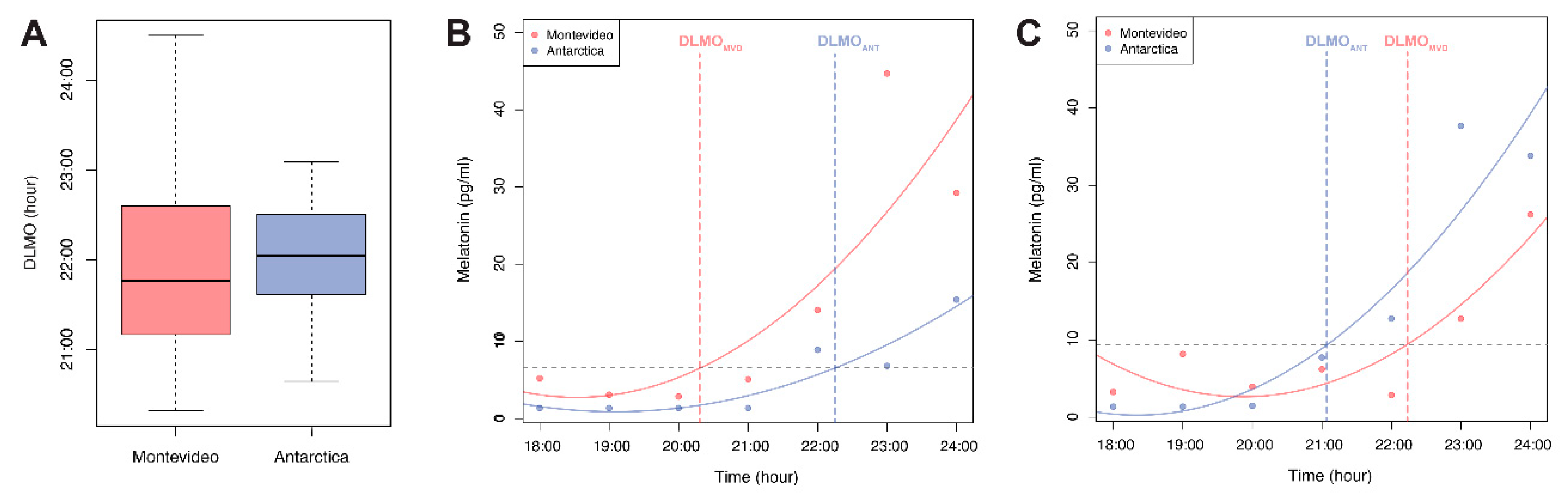
3.2. Melatonin Levels and DLMO
3.3. Chronotype-Dependence of Melatonin Levels and Consequences on Sleep Habits
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DLMO | Dim Light Melatonin Onset |
| MEQ | Mornningness-Eveningness Questionnaire |
| MCTQ | Munich Chrono-Type Questionnaire |
| MSFsc | Mid-Sleep Free days (sleep corrected) |
| SO | Sleep Onset |
| SL | Sleep Log |
| IBML | Individual Basal Melatonin Level |
References
- Wever, R.A. The Circadian System of Man; Schaefer, K.E., Ed.; Springer: Berlin/Heidelberg, Germany, 1979; ISBN 9781461261445. [Google Scholar]
- Duffy, J.F.; Wright, K.P. Entrainment of the human circadian system by light. J. Biol. Rhythm. 2005, 20, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Paranjpe, D.A.; Sharma, V.K. Evolution of temporal order in living organisms. J. Circadian Rhythm. 2005, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Chandrashekaran, M.K. Zeitgebers (time cues) for biological clocks. Curr. Sci. 2005, 89, 1136–1146. [Google Scholar]
- Roenneberg, T.; Kantermann, T.; Juda, M.; Vetter, C.; Allebrandt, K.; Vetter, C.; Allebrandt, K.V. Light and the Human Circadian Clock. In Circadian Clocks; Kramer, A., Merrow, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 217, ISBN 9783642259494. [Google Scholar]
- Roenneberg, T. The human sleep project. Nature 2013, 498, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.A. Casting light on sleep deficiency. Nature 2013, 497, 445–448. [Google Scholar] [CrossRef] [PubMed]
- de la Iglesia, H.O.; Moreno, C.; Lowden, A.; Louzada, F.; Marqueze, E.; Pilz, L.K.; Valeggia, C.; Czeisler, C.A. Ancestral sleep. Curr. Biol. 2016, 26, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin: Characteristics, concerns, and prospects. J. Biol. Rhythm. 2005, 20, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Sack, R.L. The dim light melatonin onset as a marker for circadian position. Chronobiol. Int. 1989, 6, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.L.; Blood, M.L.; Lewy, A.J. Melatonin rhythms in night shift workers. Sleep 1992, 15, 434–441. [Google Scholar] [CrossRef]
- Folkard, S. Do permanent night workers show circadian adjustment? A review based on the endogenous melatonin rhythm. Chronobiol. Int. 2008, 25, 215–224. [Google Scholar] [CrossRef]
- Lewy, A.J.; Wehr, T.A.; Goodwin, F.; Newsome, D.; Markey, S. Light suppresses melatonin secretions in humans. Science 1980, 210, 1267–1269. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Tokura, H. Bright light exposure during the daytime affects circadian rhythms of urinary melatonin and salivary immunoglobulin A. Chronobiol. Int. 1999, 16, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Mishima, K.; Okawa, M.; Shimizu, T.; Hishikawa, Y. Diminished melatonin secretion in the elderly caused by insufficient environmental illumination. J. Clin. Endocrinol. Metab. 2001, 86, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J. The dim light melatonin onset, melatonin assays and biological rhythm research in humans. Biol. Signals Recept. 1999, 8, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Kohsaka, M.; Nakamura, K.; Honma, H.; Honma, S.; Honma, K. Midday exposure to bright light changes the circadian organization of plasma melatonin rhythm in humans. Neurosci. Lett. 1997, 221, 89–92. [Google Scholar] [CrossRef]
- Takasu, N.N.; Hashimoto, S.; Yamanaka, Y.; Tanahashi, Y.; Yamazaki, A.; Honma, S.; Honma, K. Repeated exposures to daytime bright light increase nocturnal melatonin rise and maintain circadian phase in young subjects under fixed sleep schedule. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, 1799–1807. [Google Scholar] [CrossRef] [PubMed]
- Kozaki, T.; Kubokawa, A.; Taketomi, R.; Hatae, K. Light-induced melatonin suppression at night after exposure to different wavelength composition of morning light. Neurosci. Lett. 2016, 616, 1–4. [Google Scholar] [CrossRef]
- Khalsa, S.B.S.; Jewett, M.E.; Cajochen, C.; Czeisler, C.A. A phase response curve to single bright light pulses in human subjects. J. Physiol. 2003, 549, 945–952. [Google Scholar] [CrossRef]
- Gooley, J.J.; Chamberlain, K.; Smith, K.A.; Khalsa, S.B.S.; Rajaratnam, S.M.W.; Van Reen, E.; Zeitzer, J.M.; Czeisler, C.A.; Lockley, S.W. Exposure to room light before bedtime suppresses melatonin onset and shortens melatonin duration in humans. J. Clin. Endocrinol. Metab. 2011, 96, E463–E472. [Google Scholar] [CrossRef]
- Akacem, L.D.; Wright Jr, K.P.; LeBourgeois, M.K. Sensitivity of the circadian system to evening bright light in preschool-age children. Physiol. Rep. 2018, 6, e13617. [Google Scholar] [CrossRef]
- Wehr, T.A. The durations of human melatonin secretion and sleep respond to changes in daylength (photoperiod). J. Clin. Endocrinol. Metab. 1991, 73, 1276–1280. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Biological rhythms during residence in polar regions. Chronobiol. Int. 2012, 29, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Caci, H.; Adan, A.; Bohle, P.; Natale, V.; Pornpitakpan, C.; Tilley, A. Transcultural properties of the composite scale of morningness: The relevance of the “morning affect” factor. Chronobiol. Int. 2005, 22, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.S.; Folkard, S.; Schmieder, R.A.; Parra, L.F.; Spelten, E.; Almiral, H.; Sen, R.N.; Sahu, S.; Perez, L.M.; Tisak, J. Investigation of morning-evening orientation in six countries using the preferences scale. Pers. Individ. Differ. 2002, 32, 949–968. [Google Scholar] [CrossRef]
- Randler, C. Morningness-eveningness comparison in adolescents from different countries around the world. Chronobiol. Int. 2008, 25, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Tonetti, L.; Sahu, S.; Natale, V. Circadian preference in Italy and India: A comparative study in young adults. Pers. Individ. Differ. 2012, 53, 355–358. [Google Scholar] [CrossRef]
- Adan, A.; Archer, S.N.; Hidalgo, M.P.; Di Milia, L.; Natale, V.; Randler, C. Circadian typology: A comprehensive review. Chronobiol. Int. 2012, 29, 1153–1175. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Kuehnle, T.; Juda, M.; Kantermann, T.; Allebrandt, K.; Gordijn, M.; Merrow, M. Epidemiology of the human circadian clock. Sleep Med. Rev. 2007, 11, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Hida, A.; Aritake, S.; Higuchi, S.; Enomoto, M.; Kato, M. Validity of the Japanese version of the Munich ChronoType Questionnaire. Chronobiol. Int. 2014, 1–6. [Google Scholar] [CrossRef]
- Kantermann, T.; Sung, H.; Burgess, H.J. Comparing the Morningness-Eveningness Questionnaire and Munich ChronoType Questionnaire to the dim light melatonin onset. J. Biol. Rhythm. 2015, 30, 449–453. [Google Scholar] [CrossRef]
- Burgess, H.J.; Eastman, C. The dim light melatonin onset following fixed and free sleep schedules. J. Sleep Res. 2005, 14, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.R.C.; Vasconcelos, S.; Marqueze, E.C.; Lowden, A.; Middleton, B.; Fischer, F.M.; Louzada, F.M.; Skene, D.J. Sleep patterns in Amazon rubber tappers with and without electric light at home. Sci. Rep. 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.P.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Figueiro, M.G.; Plitnick, B.; Rea, M.S. The effects of chronotype, sleep schedule and light/dark pattern exposures on circadian phase. Sleep Med. 2014, 15, 1554–1564. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T.; Steer, R.A.; Carbin, M.G. Psychometric properties of the Beck Depression Inventory: Twenty-five years of evaluation. Clin. Psychol. Rev. 1988, 8, 77–100. [Google Scholar] [CrossRef]
- Roenneberg, T.; Wirz-Justice, A.; Merrow, M. Life between clocks: Daily temporal patterns of human chronotypes. J. Biol. Rhythm. 2003, 18, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Adan, A.; Almirall, H. Adaptation and standarization of a Spanish version of the morningness-eveningness questionnaire: Individual differences. Pers. Individ. Differ. 1990, 11, 1123–1130. [Google Scholar] [CrossRef]
- Roenneberg, T.; Merrow, M. Entrainment of the human circadian clock. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 293–299. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kuehnle, T.; Pramstaller, P.P.; Ricken, J.; Havel, M.; Guth, A.; Merrow, M. A marker for the end of adolescence. Curr. Biol. 2004, 14, R1038–R1039. [Google Scholar] [CrossRef]
- Horne, J.A.; Östberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Wehr, T.A.; Aeschbach, D.; Duncan Jr, W.C. Evidence for a biological dawn and dusk in the human. J. Physiol. 2001, 535, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Klerman, E.B.; Gershengorn, H.B.; Duffy, J.F.; Kronauer, R.E. Comparisons of the variability of three markers of the human circadian pacemaker. J. Biol. Rhythm. 2002, 17, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Voultsios, A.; David, J.; Dawsont, D. Salivary melatonin as a circadian phase marker: Validation and comparison to plasma melatonin. J. Biol. Rhythm. 2015, 12, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Molina, T.A.; Burgess, H.J. Calculating the dim light melatonin onset: The impact of threshold and sampling rate. Chronobiol. Int. 2011, 28, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J. Melatonin and human chronobiology. Cold Spring Harb. Symp. Quant. Biol. 2007, LXXII, 623–636. [Google Scholar] [CrossRef]
- Nagare, R.; Plitnick, B.; Figueiro, M.G. Effect of exposure duration and light spectra on nighttime melatonin suppression in adolescents and adults. Light. Res. Technol. 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin and human rhythms. Chronobiol. Int. 2006, 23, 21–37. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Zavada, A.; Gordijn, M.C.M.; Beersma, D.G.M.; Daan, S.; Roenneberg, T. Comparison of the Munich Chronotype Questionnaire with the Horne-Östberg´s morningness-eveningness score. Chronobiol. Int. 2005, 22, 267–278. [Google Scholar] [CrossRef]
- Tassino, B.; Horta, S.; Santana, N.; Levandovski, R.; Silva, A. Extreme late chronotypes and social jetlag challenged by Antarctic conditions in a population of university students from Uruguay. Sleep Sci. 2016, 9, 20–28. [Google Scholar] [CrossRef]
- Roenneberg, T. Having trouble typing? What on Earth is chronotype? J. Biol. Rhythm. 2015, 30, 487–491. [Google Scholar] [CrossRef]
- Natale, V.; Alzani, A. Additional validity evidence for the composite scale of morningness. Pers. Individ. Differ. 2001, 30, 293–301. [Google Scholar] [CrossRef]
- Taillard, J.; Philip, P.; Chastang, J.; Bioulac, B. Validation of Horne and Ostberg Morningness-Eveningness Questionnaire in a middle-aged population of French workers. J. Biol. Rhythm. 2004, 19, 76–86. [Google Scholar] [CrossRef]
- Di Milia, L.; Adan, A.; Natale, V.; Randler, C. Reviewing the psychometric properties of contemporary circadian typology measures. Chronobiol. Int. 2013, 30, 1261–1271. [Google Scholar] [CrossRef]
- Monk, T.H.; Leng, V.C. Interactions between inter-individual and inter-task differences in the diurnal variation of human performance. Chronobiol. Int. 1986, 3, 171–177. [Google Scholar] [CrossRef]
- Natale, V.; Lorenzetti, R. Influences of morningness-eveningness and time of day on narrative comprehension. Pers. Individ. Differ. 1997, 23, 685–690. [Google Scholar] [CrossRef]
- Czeisler, C.A.; Allan, J.S.; Strogatz, S.H.; Ronda, J.M.; Sánchez, R.; Ríos, C.D.; Freitag, W.O.; Richardson, G.S.; Kronauer, R.E. Bright light resets the human ccircadian pacemaker independent of the timing of the sleep-wake cycle. Science 1986, 233, 667–671. [Google Scholar] [CrossRef]
- Honma, K.; Honma, S.; Wada, T. Phase-dependent shift of free-running human circadian rhythms in response to a single bright light pulse. Experientia 1987, 43, 1205–1207. [Google Scholar] [CrossRef]
- Burešová, M.; Dvořáková, M.; Zvolsky, P.; Illnerová, H. Early morning bright light phase advances the human circadian pacemaker within one day. Neurosci. Lett. 1991, 121, 47–50. [Google Scholar] [CrossRef]
- Minors, D.; Waterhouse, J.; Wirz-Justice, A. A human phase-response curve to light. Neurosci. Lett. 1991, 133, 36–40. [Google Scholar] [CrossRef]
- St Hilaire, M.A.; Gooley, J.J.; Khalsa, S.B.S.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Human phase response curve to a 1 h pulse of bright white light. J. Physiol. 2012, 590, 3035–3045. [Google Scholar] [CrossRef]
- Van Cauter, E.; Sturis, J.; Byrne, M.M.; Blackman, J.D.; Leproult, R.; Ofek, G.; L’Hermite-Baleriaux, M.; Refetoff, S.; Turek, F.W.; Van Reeth, O. Demonstration of rapid light-induced advances and delays of the human circadian clock using hormonal phase markers. Am. J. Physiol. Metab. 1994, 266, E953–E963. [Google Scholar] [CrossRef]
- Brzezinski, A. Melatonin in humans. N. Engl. J. Med. 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Yoneyama, S.; Hashimoto, S.; Honma, K. Seasonal changes of human circadian rhythms in Antarctica. Am. J. Physiol. 1999, 277, R1091–R1097. [Google Scholar] [CrossRef]
- Premkumar, M.; Sable, T.; Dhanwal, D.; Dewan, R. Circadian levels of serum melatonin and cortisol in relation to changes in mood, sleep, and neurocognitive performance, spanning a year of residence in Antarctica. Neurosci. J. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Broadway, J.; Arendt, J.; Folkard, S. Bright light phase shifts the human melatonin rhythm during the Antarctic winter. Neurosci. Lett. 1987, 79, 185–189. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Van Dorp, C.F. Free-running rhythms of melatonin, cortisol, electrolytes, and sleep in humans in Antarctica. Am. J. Physiol. 1991, 260, R1137–R1144. [Google Scholar] [CrossRef]
- Chen, N.; Wu, Q.; Xiong, Y.; Chen, G.; Song, D.; Xu, C. Circadian rhythm and sleep during prolonged Antarctic residence at Chinese Zhongshan Station. Wilderness Environ. Med. 2016, 27, 458–467. [Google Scholar] [CrossRef]
- Midwinter, M.J.; Arendt, J. Adaptation of the melatonin rhythm in human subjects following night-shift work in Antarctica. Neurosci. Lett. 1991, 122, 195–198. [Google Scholar] [CrossRef]
- Broadway, J.; Arendt, J. Seasonal and bright light changes of the phase position of the human melatonin rhythm in Antarctica. Arctic Med. Res. 1988, 47, 201–203. [Google Scholar]
- Arendt, J.; Middleton, B. Human seasonal and circadian studies in Antarctica (Halley, 75°S). Gen. Comp. Endocrinol. 2018, 258, 250–258. [Google Scholar] [CrossRef]
- Figueiro, M.G.; Rea, M.S. Lack of short-wavelength light during the school day delays dim light melatonin onset (DLMO) in middle school students. Neuro. Endocrinol. Lett. 2010, 31, 92–96. [Google Scholar]
- Gabel, V.; Maire, M.; Reichert, C.F.; Chellappa, S.L.; Schmidt, C.; Hommes, V.; Viola, A.U.; Cajochen, C. Effects of artificial dawn and morning blue light on daytime cognitive performance, well-being, cortisol and melatonin levels. Chronobiol. Int. 2013, 30, 988–997. [Google Scholar] [CrossRef]
- Czeisler, C.A.; Kronauer, R.E.; Allan, J.S.; Duffy, J.F.; Jewett, M.E.; Brown, E.N.; Ronda, J.M. Bright light induction of strong (type 0) resetting of the human circadian pacemaker. Science 1989, 244, 1328–1333. [Google Scholar] [CrossRef]
- Rosenthal, N.E.; Joseph-Vanderpool, J.R.; Levendosky, A.A.; Johnston, S.H.; Allen, R.; Kelly, K.A.; Souetre, E.; Schultz, P.M.; Starz, E. Phase-shifting effects of bright morning light as treatment for delayed sleep phase syndrome. Sleep 1990, 13, 354–361. [Google Scholar]
- Minors, D.S.; Waterhouse, J.M.; Wirz-Justicg, A. A human phase-response curve to light. Neurosci. Lett. 1991, 13, 36–40. [Google Scholar] [CrossRef]
- Pattyn, N.; Mairesse, O.; Cortoos, A.; Marcoen, N.; Neyt, X.; Meeusen, R. Sleep during an Antarctic summer expedition: New light on “polar insomnia”. J. Appl. Physiol. 2017, 122, 788–794. [Google Scholar] [CrossRef][Green Version]
- Lewy, A.J.; Cutler, N.L.; Sack, R.L. The endogenous melatonin profile as a marker for circadian phase position. J. Biol. Rhythm. 1999, 14, 227–236. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Smits, M.; Spence, W.; Srinivasan, V.; Cardinali, D.P.; Lowe, A.D.; Kayumov, L. Dim light melatonin onset (DLMO): A tool for the analysis of circadian phase in human sleep and chronobiological disorders. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2007, 31, 1–11. [Google Scholar] [CrossRef]
- Benloucif, S.; Guico, M.J.; Reid, K.J.; Wolfe, L.F.; L’Hermite-Balériaux, M.; Zee, P.C. Stability of melatonin and temperature as circadian phase markers and their relation to sleep times in humans. J. Biol. Rhythm. 2005, 20, 178–188. [Google Scholar] [CrossRef]
- Stothard, E.R.; McHill, A.W.; Depner, C.M.; Birks, B.R.; Moehlman, T.M.; Ritchie, H.K.; Guzzetti, J.R.; Chinoy, E.D.; LeBourgeois, M.K.; Axelsson, J.; et al. Circadian entrainment to the natural light-dark cycle across seasons and the weekend. Curr. Biol. 2017, 27, 508–513. [Google Scholar] [CrossRef]
- Claustrat, B.; Leston, J. Melatonin: Physiological effects in humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef]
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Iwamoto, J.; Okamoto, N.; Tomioka, K.; Nezu, S.; Ikada, Y.; Kurumatani, N. Positive effect of daylight exposure on nocturnal urinary melatonin excretion in the elderly: A cross-sectional analysis of the HEIJO-KYO study. J. Clin. Endocrinol. Metab. 2012, 97, 4166–4173. [Google Scholar] [CrossRef]
- Stephan, F.K. The “other” circadian system: Food as a zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Wehrens, S.M.T.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; Skene, D.J.; Johnston, J.D. Meal timing regulates the human circadian system. Curr. Biol. 2017, 27, 1768–1775. [Google Scholar] [CrossRef]
- Kräuchi, K.; Cajochen, C.; Werth, E.; Wirz-justice, A. Alteration of internal circadian phase relationships after morning versus evening carbohydrate-rich meals in humans. J. Biol. Rhythm. 2002, 17, 364–376. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Hashimoto, S.; Masubuchi, S.; Natsubori, A.; Nishide, S. Differential regulation of circadian melatonin rhythm and sleep-wake cycle by bright lights and nonphotic time cues in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R546–R557. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Skene, D.J. Social influences on mammalian circadian rhythms: Animal and human studies. Biol. Rev. 2004, 79, 533–556. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Skene, D.J. Nonphotic entrainment in humans? J. Biol. Rhythm. 2005, 20, 339–352. [Google Scholar] [CrossRef]
- Davidson, A.J.; Menaker, M. Birds of a feather clock together–sometimes: Social synchronization of circadian rhythms. Curr. Opin. Neurobiol. 2003, 13, 765–769. [Google Scholar] [CrossRef]
- Phillips, A.J.K.; Vidafar, P.; Burns, A.C.; McGlashan, E.M.; Anderson, C.; Rajaratnam, S.M.W.; Lockley, S.W.; Cain, S.W. High sensitivity and interindividual variability in the response of the human circadian system to evening light. Proc. Natl. Acad. Sci. USA 2019, 116, 12019–12024. [Google Scholar] [CrossRef]
- Leung, M.; Tranmer, J.; Hung, E.; Korsiak, J.; Day, A.G.; Aronson, K.J. Shift work, chronotype, and melatonin patterns among female hospital employees on day and night shifts. Cancer Epidemiol. Biomark. Prev. 2016, 25, 830–838. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Interval (h) | Location | Light Exposure (lx) | |||
|---|---|---|---|---|---|
| All Participants | Early Chronotypes | Late Chronotypes | p-Value | ||
| 7:00–9:59 | Montevideo | 869.13 ± 678.48 | 1070.66 ± 710.29 | 587.00 ± 581,89 | 0.24 |
| Antarctica | 2554.87 ± 187.32 | 2607.73 ± 177.56 | 2480.88 ± 193.56 | 0.27 | |
| p-value | 0.00002 | 0.002 | 0.002 | ||
| 10:00–12:59 | Montevideo | 2733.64 ± 1536.76 | 2943.78 ± 1923.95 | 2439.46 ± 869.84 | 0.60 |
| Antarctica | 8459.67 ± 957.59 | 8787.97 ± 959.46 | 8000.06 ± 829.56 | 0.17 | |
| p-value | 0.00001 | 0.001 | 0.0002 | ||
| 19:00–21:59 | Montevideo | 189.49 ± 76.38 | 170.00 ± 68.38 | 216.77 ± 86.22 | 0.32 |
| Antarctica | 699.81 ± 207.15 | 745.41 ± 258.33 | 653.96 ± 95.76 | 0.39 | |
| p-value | 0.00001 | 0.001 | 0.004 | ||
| 22:00–00:59 | Montevideo | 137.01 ± 91.62 | 100.41 ± 42.41 | 188.26 ± 121.49 | 0.10 |
| Antarctica | 200.12 ± 123.41 | 231.47 ± 140.14 | 156.23 ± 91.08 | 0.32 | |
| p-value | 0.16 | 0.044 | 0.50 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.; Simón, D.; Pannunzio, B.; Casaravilla, C.; Díaz, Á.; Tassino, B. Chronotype-Dependent Changes in Sleep Habits Associated with Dim Light Melatonin Onset in the Antarctic Summer. Clocks & Sleep 2019, 1, 352-366. https://doi.org/10.3390/clockssleep1030029
Silva A, Simón D, Pannunzio B, Casaravilla C, Díaz Á, Tassino B. Chronotype-Dependent Changes in Sleep Habits Associated with Dim Light Melatonin Onset in the Antarctic Summer. Clocks & Sleep. 2019; 1(3):352-366. https://doi.org/10.3390/clockssleep1030029
Chicago/Turabian StyleSilva, Ana, Diego Simón, Bruno Pannunzio, Cecilia Casaravilla, Álvaro Díaz, and Bettina Tassino. 2019. "Chronotype-Dependent Changes in Sleep Habits Associated with Dim Light Melatonin Onset in the Antarctic Summer" Clocks & Sleep 1, no. 3: 352-366. https://doi.org/10.3390/clockssleep1030029
APA StyleSilva, A., Simón, D., Pannunzio, B., Casaravilla, C., Díaz, Á., & Tassino, B. (2019). Chronotype-Dependent Changes in Sleep Habits Associated with Dim Light Melatonin Onset in the Antarctic Summer. Clocks & Sleep, 1(3), 352-366. https://doi.org/10.3390/clockssleep1030029