Abstract
This article presents the first results of the zooarchaeological analysis of Chalcolithic levels of the Barrio del Castillo site (Torrejón de Ardoz), located within the Aldovea complex, which also contains evidence from other prehistoric periods. The Barrio del Castillo sector reveals an occupation pattern characterized by pit and silo fields, a common phenomenon in the Tagus Middle Valley’s Prehistory. This study focuses on the Chalcolithic phase, which exhibits two types of faunal accumulations: one linked to domestic activities (referred to as domestic assemblages), and another with a more symbolic character, involving the burial of articulated or semi-articulated animals, mainly dogs, designated as ritual deposits. The first type of accumulation is dominated by domestic species like caprines (sheep and goats) and cattle. Other domestic species, like pigs and dogs, alongside wild species, like horses and deer, appear in smaller quantities. On the other hand, ritual deposits are largely dominated by dogs in the Chalcolithic phase. This paper emphasizes the marked contrast between the two types of accumulations, domestic and ritual, and highlights the implications that these might have toward a better understanding of the world of the first metallurgical communities of the inner Iberian Peninsula.
1. Introduction
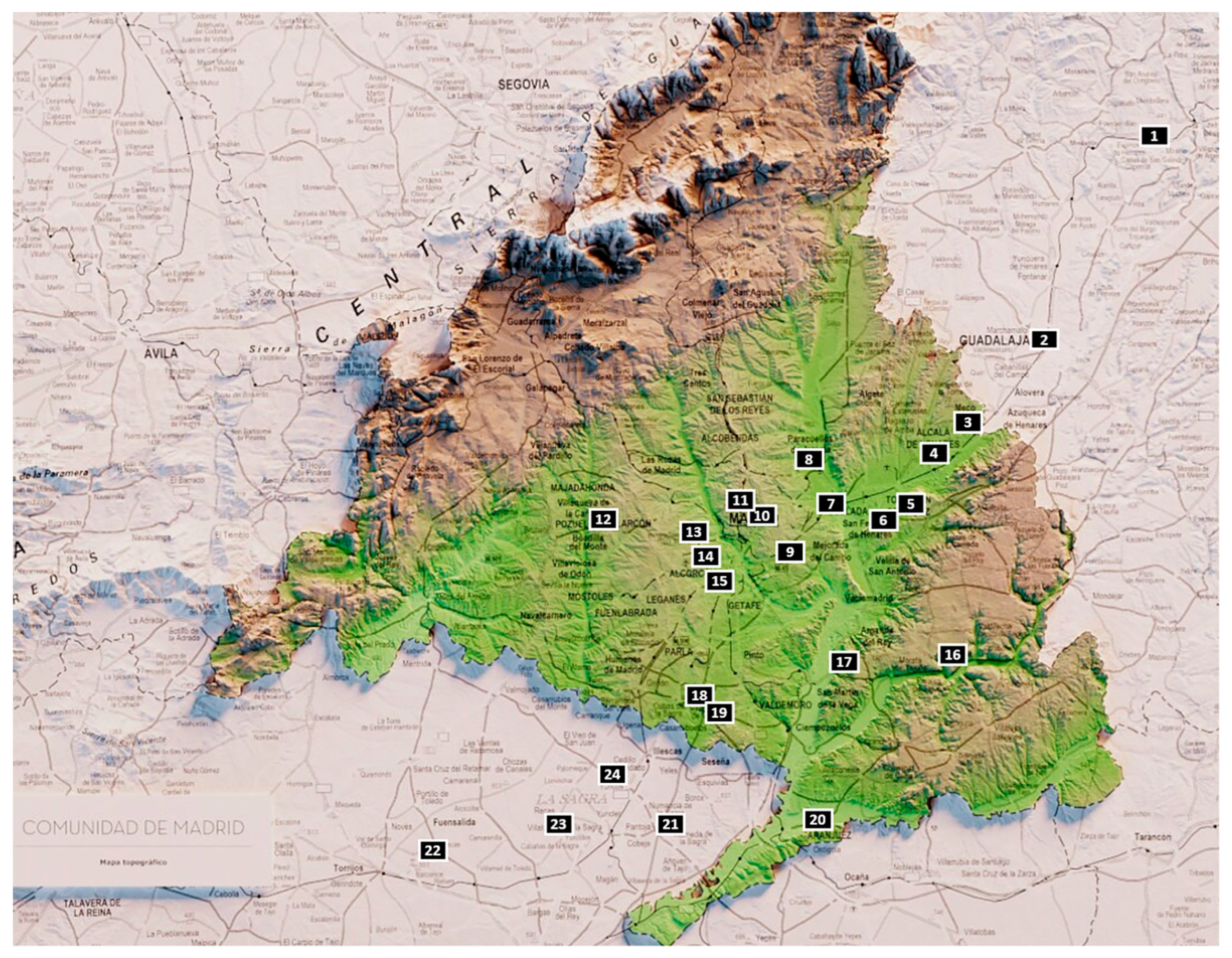
Until recently, zooarchaeological evidence for the Chalcolithic period in the interior of the Iberian Peninsula was scarce, primarily due to limited excavations and small sample sizes [1,2]. However, research carried out in recent years has substantially expanded the available dataset. Significant contributions have emerged from recently excavated Chalcolithic sites, such as Zanjillas [3], Aldovea [4], Camino de las Yeseras [5], Soto de Henares [6], and La Loma del Lomo III [7], among others (Figure 1). These sites have enabled more in-depth investigations into how Chalcolithic communities in the region managed and exploited faunal resources.
Figure 1.
Locations of Chalcolithic sites with faunal remains for the middle Tagus Valley: 1. Loma del lomo (Cogolludo), 2. Aguas Vivas (Guadalajara), 3. La Esgaravita (Alcalá de Henares), 4. Las Matillas (Alcalá de Henares), 5. Soto de Henares (Torrejón de Ardoz), 6. Aldovea and Barrio del Castillo (Torrejón de Ardoz), 7. Camino de las Yeseras (Coslada), 8. El Capricho (Barajas), 9. Los Berrocales (Vicálvaro), 10. Loma de Chicalana (Madrid), 11. Angosto de los Moncebos (Madrid), 12. La Cabeceras (Pozuelo de Alarcón), 13. El Espinillo (Villaverde), 14. Fábrica de Euskalduna and Tejar del Sastre (Villaverde), 15. El Ventorro (Villaverde), 16. Barranco del Herrero (San Martín de la Vega), 17. Juan Barbero (Tielmes de Tajuña), 18. La Cuesta and Humanejos (Torrejón de Velasco), 19. Las Zanjillas (Torrejón de Velasco), 20. Huerta de los Cabreros (Aranjuez), 21. Las Mayores (Numancia de la Sagra, Toledo), 22. Las Vegas (Huecas, Toledo), 23. Cerrocuquillo (Villaluenga de La Sagra, Toledo); 24. Entreviñas (Toeldo).
Current evidence suggests that early metallurgical societies in the interior of the Iberian Peninsula practiced agropastoral economies, with a particular emphasis on cereal cultivation [8] and livestock herding—especially mixed flocks of goats, sheep, and cattle [1,2,3,4].
This article presents new data from the recently excavated site of Barrio del Castillo, part of the Aldovea Complex in Torrejón de Ardoz, excavated under the direction of one of the authors (M.M.). The site has yielded substantial prehistoric faunal remains, with the Chalcolithic assemblages being particularly significant.
What distinguishes this assemblage is the presence of animal remains in two clearly differentiated contexts. The first is associated with domestic activities, characterized by a high degree of bone fragmentation and co-occurrence with lithic tools and ceramic sherds. The second context appears to be symbolic in nature, involving the burial of fully or partially articulated animals. This article explores the significance of these contrasting assemblages and their broader implications for understanding Chalcolithic societies in the region.
2. Barrio del Castillo
The Barrio del Castillo site is one of three that comprise the broader Aldovea complex, located in Torrejón de Ardoz in the province of Madrid [9] (Figure 1 and Figure 2). The complex includes the sites of Camino de Baracalde, Aldovea (analyzed in [4,10]), and Barrio del Castillo.

Figure 2.
(A): Location of the Aldovea area in Torrejón de Ardoz, and the three corresponding sectors: Aldovea, Barrio del Castillo, and Camino de Baracalde. (B): Orthophoto of Barrio del Castillo at the end of the excavation process, where all the excavated silos can be seen. (C,D): Examples of silo fields in Barrio del Castillo. (Photos: M.M). See orthophoto of Figure 2B in Supplementary File.
At Barrio del Castillo, more than 800 circular features—primarily silos and pits—have been documented, spanning several cultural phases from the Neolithic to the Bronze Age. Of these, 223 have been attributed to the Chalcolithic phase, based on associated material culture and primarily ceramics (Figure 3 and Figure 4). Additionally, a radiocarbon date was obtained from a dog skeleton found from ritual deposit SU 6830, yielding a calibrated date of 4570 ± 15 BP. This date aligns well with the established chronology of the Chalcolithic period in the Iberian Peninsula [11]. The settlement pattern identified at Barrio del Castillo is characteristic of sites located in fluvial valleys along the middle Tagus basin. These sites are commonly referred to as “pit-fields” or “silo-fields” across a broad chronological range [12,13,14,15,16]. The earliest Neolithic phase provides only limited evidence of occupation, with just a few silos identified from this period. In contrast, the post-Chalcolithic phase examined in this study reveals a substantial number of pits and basins, which can be attributed to the Protocogotas and Cogotas I cultural phases.

Figure 3.
Pottery from the Chalcolithic levels of Barrio del Castillo (Torrejón de Ardoz). (A,C): Plain globular bowls; (B): Fragment of a cheese strainer; (D): Sherd with impressed and incised decoration; (E): Rim with impressed decoration; (F): Bowl base. (Photo: M.M.).
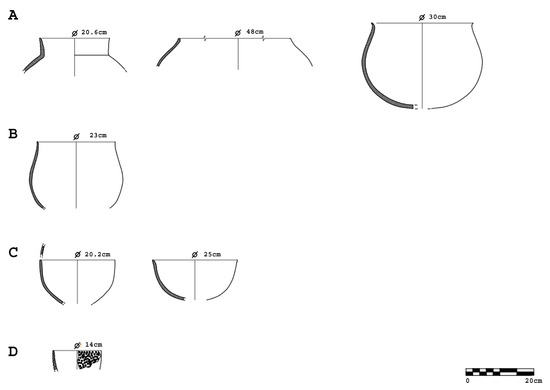
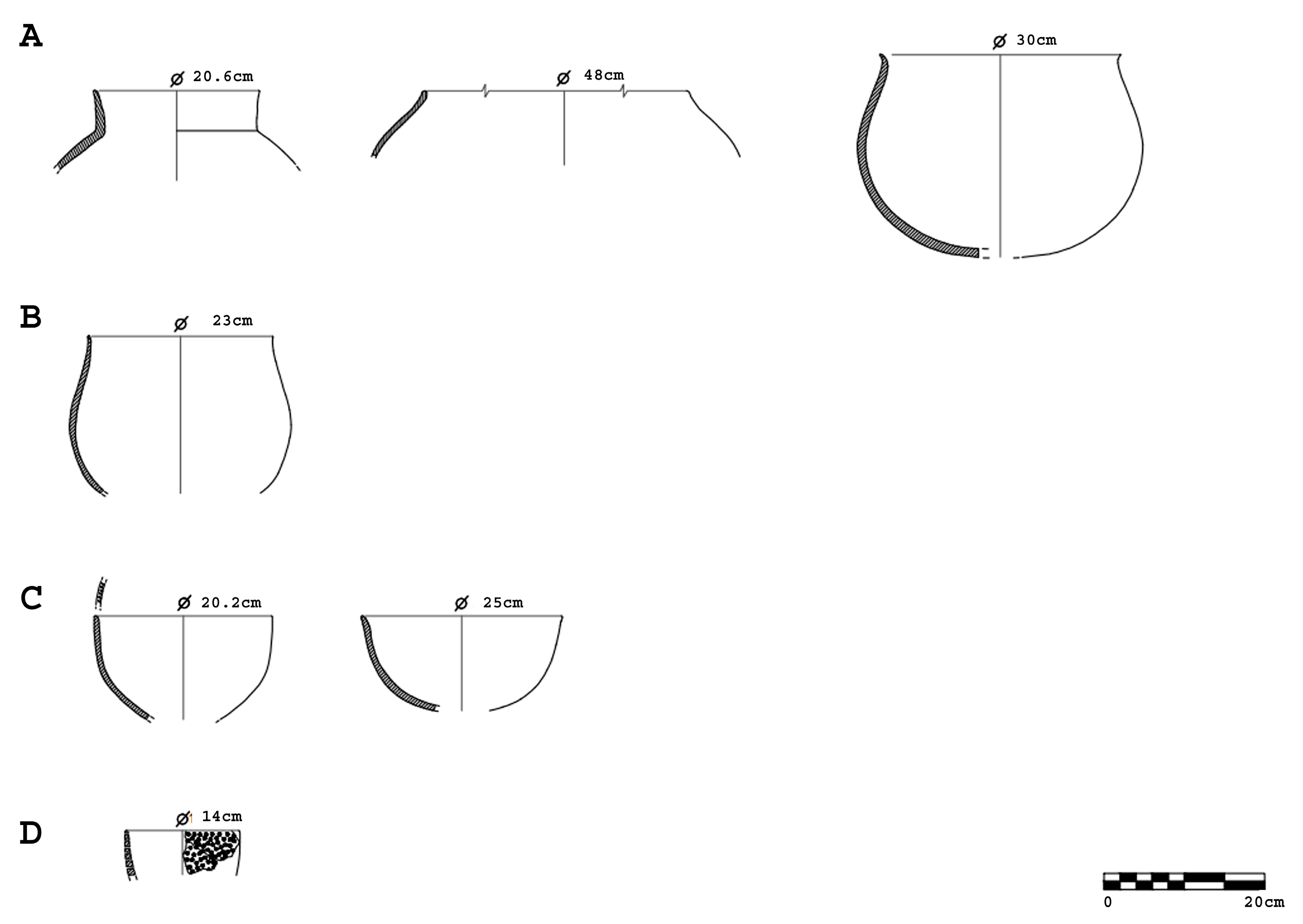
Figure 4.
Pottery forms from the Chalcolithic levels of Barrio del Castillo. (A): Storage jars; (B): cooking pot; (C): bowls; and (D): cheese strainer. (Diagram: M.M.).
The ceramic assemblage from the Chalcolithic levels at Barrio del Castillo comprises 43,124 fragments, of which 7902 are diagnostic. These include complete and semi-complete vessels, rims, handles, bases, carinations, mamelons (diagnostic rounded appliqués), and decorated fragments (Figure 3). The analysis of this assemblage reveals a predominance of globular storage and cooking vessels (80%), with the remainder consisting of bowls, small pots, plates, cheese strainers, and cups (Figure 4).
The ceramics were primarily fired in reducing or mixed atmospheres. A total of 95.5% of the assemblage is undecorated, while 4.5% displays various decorative techniques, including impressions, incisions, and appliqués (Figure 3).
Lithic artifacts are also abundant at the site, with notable examples including projectile points from SU 2891, a fibrolite adze from SU 2381, and polished adzes from SU 6471. Both flint and quartzite were used as raw materials, with quartzite being more prevalent, often in the form of cobbles used for structural purposes (Figure 5). These were primarily employed in the construction of pit and silo bases, or as sealing elements. In SU 6836, however, quartzite cobbles were notably used to cover the remains of a juvenile dog in a ritual context. Finally, granite artifacts are also present, mainly in the forms of hand mills and grinding stones.
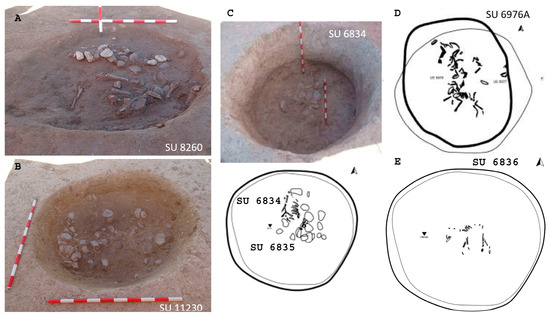
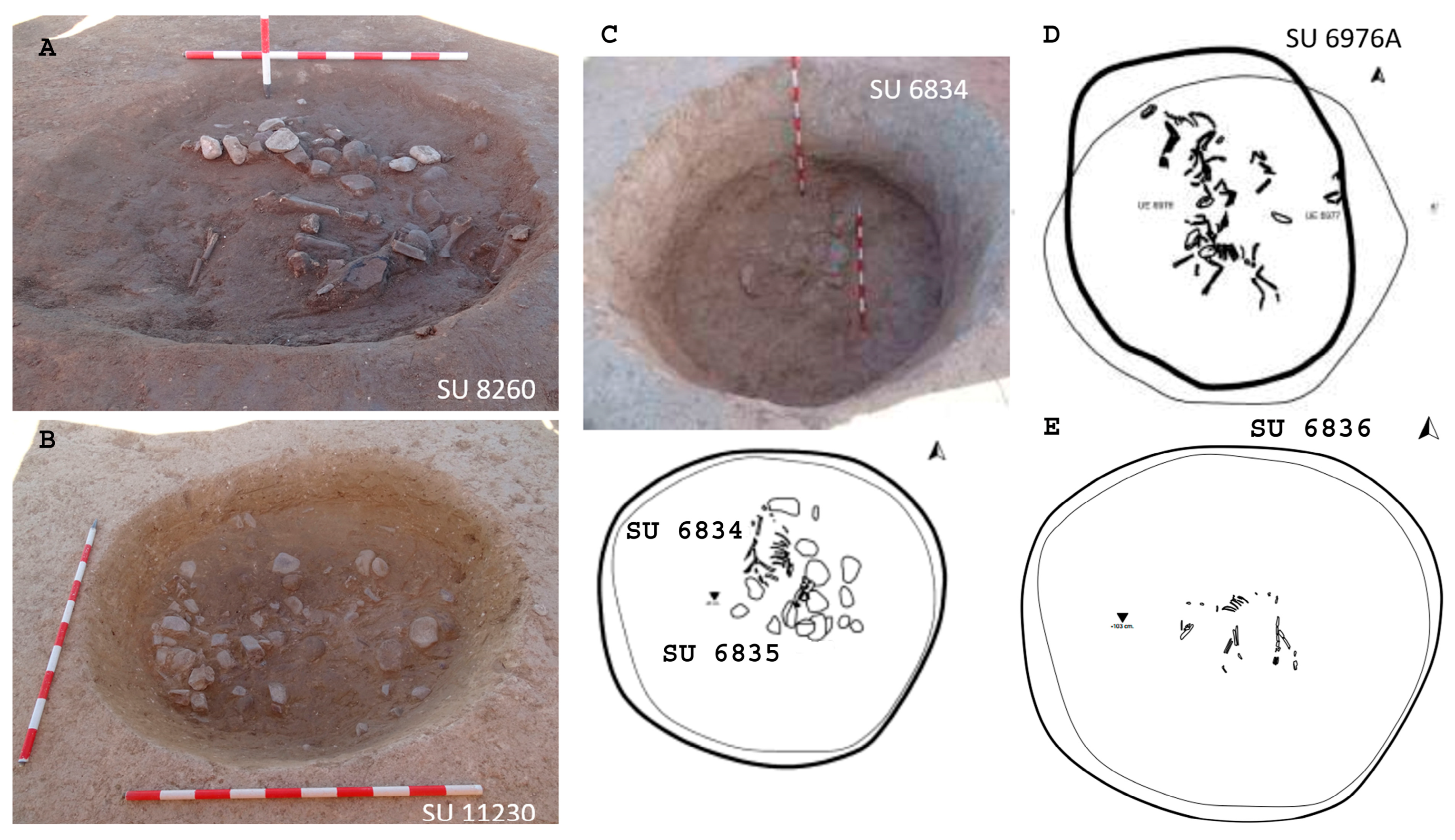
Figure 5.
Pits from Barrio del Castillo. (A,B): Domestic pits containing faunal remains and quartzite cobbles. (C–E): Pits corresponding to SU 6834 (C), SU 6976A (D), and SU 6836 (E). (Photo: M.M.).
3. Materials and Methods
This study focuses on the analysis of faunal remains excavated in 2019 by the private archaeological company Áqaba Arqueólogos S.L., which directed that year’s fieldwork campaign. The animal bone assemblages were classified into two categories, those recovered from domestic contexts and those from ritual deposits, the latter of which was defined by carefully arranged, articulated, or semi-articulated skeletons.
In total, 22,771 faunal fragments were analyzed. Of these, 3978 fragments originated from Chalcolithic domestic contexts, while 2992 came from ritual deposits associated with the same period.
A taxonomic identification approach was employed to assess species representation and mortality patterns. These data also allowed for a comparative analysis of the two types of faunal accumulations: domestic and ritual. Taxonomic identification was conducted using the reference collection housed in the Faculty of Geography and History at the Universidad Complutense de Madrid, as well as several reference atlases [17,18,19,20,21]. Additional identification aids were consulted for specific taxa, such as for distinguishing between sheep and goats [22,23,24,25], and between domestic pigs and wild boars [26].
Following taxonomic identification, fragments were quantified by the number of identified specimens (NISP) and the minimum number of individuals (MNI). Mortality profiles were established by classifying individuals as adults or juveniles/infants.
Fragments that could not be assigned to a specific taxon were grouped into size categories: large (e.g., Equus sp., Bos taurus), medium (e.g., Cervus elaphus), and small (e.g., Ovis aries, Capra hircus, Capreolus capreolus, Sus scrofa/domesticus, and Canis familiaris). Fragments were only considered indeterminate when they could not be identified taxonomically, anatomically, or by size.
Mortality analysis was based on tooth eruption, tooth wear, and epiphyseal fusion. For dental aging, established criteria were followed for goats [27], sheep [28], cattle [29], and pigs [30,31]. Age estimations based on epiphyseal fusion followed the guidelines provided in [17]. Four mortality categories were differentiated: neonates; infants, with the emergence of either the upper or lower M2; juveniles, with the emergence of either the upper or lower permanent M3 and P4; and adults when all dentition was present.
Skeletal profiles were also analyzed. Appendicular elements include limb bones, such as the humerus, femur, radius, ulna, tibia, and metapodials. Axial elements comprise ribs, vertebrae, scapulae, and pelvic bones, following the classification proposed in [32]. Cranial elements refer to all bones of the skull. A distinction was made between forelimbs (scapula, humerus, radius, ulna, carpals, and metacarpals) and hindlimbs (pelvis, femur, tibia, tarsals, and metatarsals). Finally, taphonomic alterations were assessed using jeweler’s loupes with 10–20× magnification.
4. Results
The Chalcolithic phase at Barrio del Castillo yielded a substantial quantity of faunal remains, with 3978 bone fragments recovered from 148 distinct stratigraphic units associated with domestic contexts.
The results indicate that domestic fauna are significantly more important than wild fauna, which represented 8% of the MNI and 3.5% of the NISP. Among domestic taxa, caprines and cattle are the most represented taxa, both in terms of the total number of identified specimens (NISP) and the minimum number of individuals (MNI). Specifically, cattle account for 41.9% of the NISP and 35.1% of the MNI, while caprines represent over 44% of both indices, NISP and MNI. However, the relatively high proportion (10.3%) of large indeterminate fragments may suggest an underrepresentation of cattle (Table 1).

Table 1.
Taxonomic representation by NISP (number of identified specimens) and MNI (minimum number of individuals) in domestic contexts and ritual deposits of Barrio del Castillo during its Chalcolithic phase.
Other domestic species, such as pigs and dogs, are also present, but in considerably lower proportions, each constituting less than 10% of the NISP and MNI. Nevertheless, these species are the predominant taxa in ritual deposits. Additionally, small numbers of wild taxa—including horses and deer—have been identified, though their representation remains low, comprising less than 10% of the MNI (Table 1). Their presence suggests that hunting may have complemented subsistence strategies at the site.
In stark contrast to domestic contexts, the 21 stratigraphic units associated with ritual deposits show a clear predominance of dogs, in terms of both NISP and MNI. Dogs account for 55% of the total sample and 45% of the MNI. They are followed in frequency by pigs and cattle. Notably, while pigs ranked second in NISP, caprines occupied the second position in MNI (Table 1), due to the fact that not all species identified within ritual deposits are necessarily the result of intentional deposition. Some may represent incidental inclusions. Table 2 presents only those animals that were found fully or partially articulated and are thus interpreted as intentional ritual deposits.
Table 2.
Faunal representation in ritual deposits, with animals stated as complete or partial. Additionally, fragments of other species associated with the deposits, but not found to be intentional in their nature. Bos refers to cattle; Canis to dog; Sus to pig; and Ovis/Capra to Caprine: sheep and goat.
According to these criteria, the intentional ritual deposits include ten dogs, one complete and one partial cattle, one complete and two partial pigs, and one partial sheep. This clearly demonstrates the importance that dogs must have held for the people in Barrio del Castillo. Other faunal remains found in these stratigraphic units—some even associated with human burials, as in the case of SU 10893—were considered incidental and not deliberately interred.
Certain contexts present interpretive challenges. For instance, SU 6976C yielded highly fragmented and disarticulated bones. Although there is some evidence suggestive of a dog deposit, the absence of securely identifiable axial elements has led to the exclusion of this stratigraphic unit from the list of confirmed ritual deposits.
Similarly, in SU 6076I and SU 6976J, a small number of appendicular elements were recovered in a semi-articulated state. Due to this partial articulation, these contexts are currently classified as potential ritual deposits, pending further analysis.
Taken together, these observations reveal a marked distinction between the two contexts: domestic assemblages are dominated by mixed herds of cattle and caprines (sheep and goats), while ritual deposits are overwhelmingly characterized by the presence of dogs (Table 1 and Table 2).
Mortality profiles also reveal notable differences between the two types of faunal assemblages (Table 3). In domestic contexts, adult individuals predominate across all taxa. In contrast, ritual deposits are characterized by a high proportion of infant dogs and pigs. which constitute a significant portion of the overall assemblage.
Table 3.
Mortality patterns observed in the Chalcolithic phases of Barrio del Castillo.
In summary, from a taxonomic standpoint, there is a distinct contrast between the faunal assemblages from domestic contexts and those from ritual deposits. Domestic contexts are predominantly composed of adult individuals from mixed herds of sheep, goats, and cattle. Opposingly, ritual deposits show a marked prevalence of infant dogs, along with both adult and infant pigs, while cattle and sheep are represented only by adults (Table 3).
In domestic contexts, all skeletal elements are represented. However, there is an anatomical predominance of cranial elements, accounting for 59.1% of Bos remains and 46.4% of Sus remains (Table 4). In contrast, postcranial elements are markedly underrepresented. Among the appendicular elements, forelimb bones are more prevalent than hindlimb bones, especially in caprines and cattle, the most abundant taxa. This overrepresentation of forelimb elements and underrepresentation of hindlimb elements may be attributed to the significant fragmentation of these assemblages. Consequently, there is a notable increase in the number of indeterminate fragments—1801 remains—particularly diaphyseal fragments that could not be assigned to specific anatomical elements.
Table 4.
Skeletal profiles of domestic contexts. Total O/C makes reference to Ovis, Capra, and Ovis/Capra together; Sus makes reference to Sus sp. and Sus domesticus. See Supplementary Material for more details on the skeletal profiles of each species in each unit.
The skeletal profiles from ritual deposits vary depending on the specific circumstances of each deposit. This can be seen, for example, in the differences between forelimb–hindlimb proportions for cattle in SU 7551 and 8261 (Table 5). Generally, all skeletal elements are represented with certain elements predominating, depending on the species involved. In the case of dogs, axial elements are the most common (Table 5), while cranial elements are more frequent in the deposits of cattle and pigs. This pattern reflects the fact that the deposits of cattle and pigs are often partial. For example, SU 8231 contains only a pig cranium, and, in the case of SU 7551 there is a shortage of appendicular elements from what would be expected. In other cases, the absence of elements may be due to the fact that they could not be assigned with certainty to one taxon or another; this is the case for SU 6976J, in which numerous axial bones could not be assigned to either dogs or pigs (Table 5).
Table 5.
Skeletal profiles of main animals for ritual deposits.
Regarding bone surface modifications, the bones from domestic contexts show considerable fragmentation and poor preservation of cortical surfaces. The primary visible alterations include root etching, biochemicals, and diagenetic changes. As a result, cut marks were observed on only 1% of the faunal assemblages from domestic contexts. In ritual deposits, the poor preservation of cortical surfaces hindered the identification of any evidence of human-induced bone surface modifications (Figure 6). Furthermore, in both types of accumulations, bones exhibiting epiphyseal collapse, likely caused by carnivores, possibly dogs, have been observed (Figure 6). As with the cut marks, the poor state of bone preservation has also hindered the identification of tooth marks.

Figure 6.
(A,C): Mandible with bad bone surface preservation. (B): Large bone with collapse of epiphysis.
5. Discussion
The zooarchaeological assemblage from the Chalcolithic levels of Barrio del Castillo reveals two distinct types of faunal contexts: domestic contexts, dominated by mixed herds of caprines and cattle; and ritual deposits, in which dogs are the most prominent species, followed by pigs and cattle.
These assemblages also differ in their mortality patterns. Domestic contexts are primarily composed of adult individuals, while ritual deposits are dominated by infant dogs and pigs. Another key distinction lies in the preservation of skeletal remains: domestic contexts show a higher level of fragmentation and no articulated bones, whereas ritual deposits are characterized by low fragmentation and the presence of complete, articulated skeletons.
These findings align with those from other archaeological sites in the middle Tagus Valley, such as Aldovea [4], where domestic assemblages were mainly composed of caprines and cattle, while ritual deposits featured a high number of dogs. In both sites, mortality patterns are consistent, with domestic contexts predominantly containing adult individuals, while ritual deposits consist primarily of infant dogs and pigs, along with adult cattle.
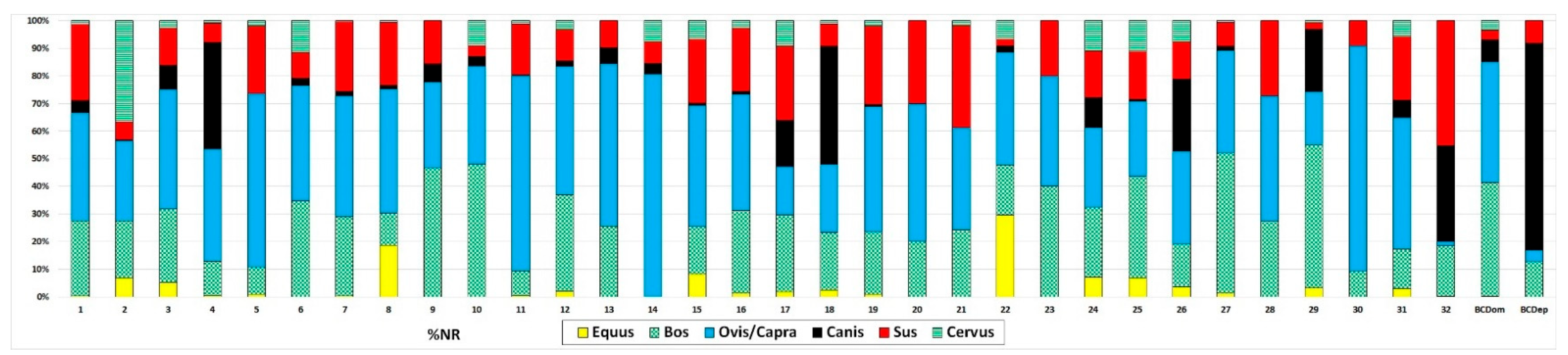
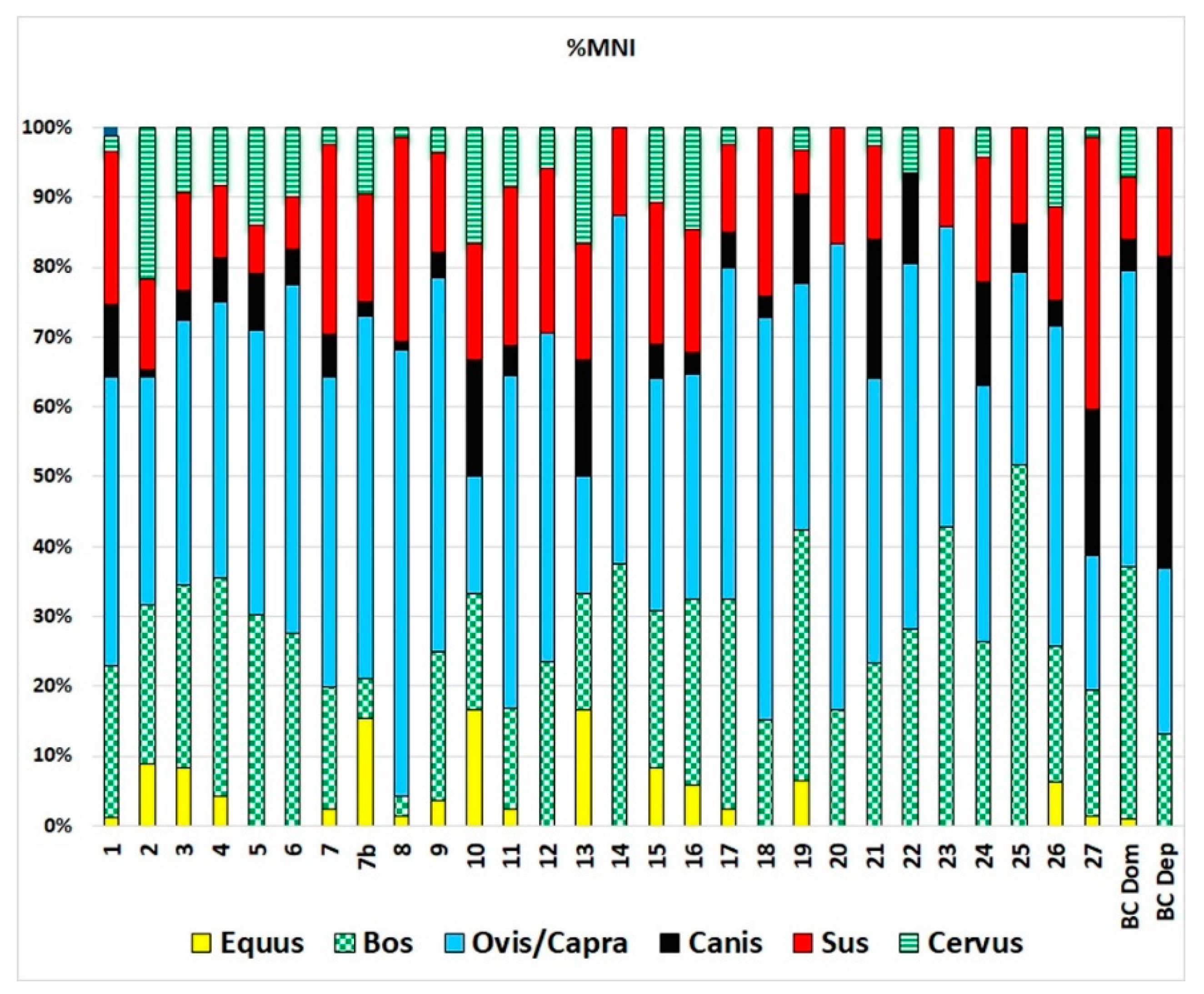
The taxonomic distribution observed at Barrio del Castillo also mirrors patterns found at other sites in the middle Tagus Valley, as seen in the number of identified specimens (NISP; Figure 7) and minimum number of individuals (MNI; Figure 8). In line with most other sites, caprines—including sheep and goats—are the most important species, followed by cattle, pigs, dogs, and wild game such as deer. Ritual deposits, though rare, display the same general characteristics as those at Barrio del Castillo, with a predominance of dogs, followed by pigs, and then cattle, which contrasts sharply with the taxonomic profiles observed in domestic contexts.
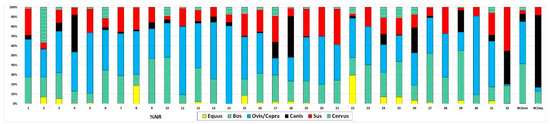
Figure 7.
Taxonomic representation of animals most yielded by Chalcolithic sites of the middle Tagus Valley according to N. 1: Loma del Lomo; 2: Aguas Vivas sect; 2, 3. Aguas Vivas sect; 0, 4: Aguas Vivas sect. Vía pecuaria; 5: La Esgaravita; 6: Las Matillas; 7: Camino de las Yeseras Southern area, 8: Camino de las Yeseras central area; 9: Camino de las Yeseras cabin E; 10: Aguas Vivas; 11: Capricho, 12: Loma de Chiclana; 13: Tejar del Sastre; 14: Priego de los Mancebos, 15: Las Cabeceras; 16: El Espinillo, Chalcolithic phase; 17: El Espinillo, Bell Beaker phase; 18: El Espinillo, Chalcolithic–Bronze phase; 19: Ventorro; 20: Fábrica de Euskalduna; 21: Barranco del Herrero; 22: Juan Barbero; 23: La Cuesta; 24: Humanejos, Chalcolithic phase; 25: Humanejos, Bell Beaker phase; 26: Humanejos, Chalcolithic–Bronze phase; 27: Zanjillas; 28: Huerta de los Cabreros; 29: Entreviñas; 30: Cerro Cuquillo; 31: Aldovea, domestic assemblages; 32: Aldovea deposits, BCDom: Barrio del Castillo domestic contexts, BCDep: Barrio del Castillo deposits.
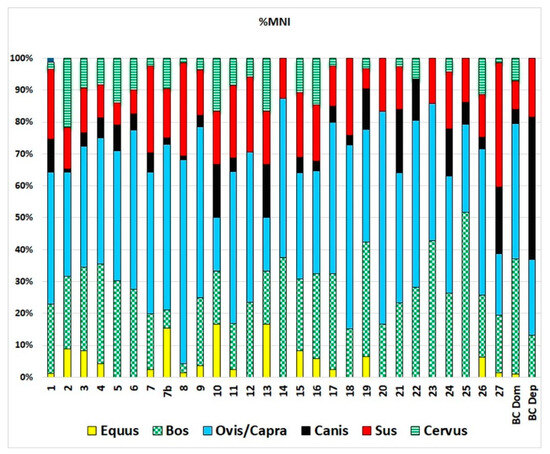
Figure 8.
Taxonomic representation of animals most represented in Chalcolithic sites of the middle Tagus Valley according to MNI. 1: Loma del Lomo; 2: Aguas Vivas sect. Vía pecuaria; 3: Aguas Vivas sect; 2, 4: Aguas Vivas sect; 0, 5: Aguas Vivas sect. 1, 6: Matillas; 7: Camino de las Yeseras, southern area; 7b: Camino de las Yeseras, central area; 8: Capricho; 9: Loma Chiclana; 10: Priego de los Mancebos; 11: Ventorro; 12: Barranco del Herrero; 13: Juan Barbero; 14: La Cuesta; 15: Humanejos, Chalcolithic phase; 16: Humanejos, Bell Beaker phase; 17: Zanjillas; 18: Huerta de los Cabreros; 19: Entreviñas; 20: Cerro Cuquillo; 21: Soto de Henares, early Chalcolithic phase; 22: Soto de Henares, Chalcolithic phase; 23: Soto de Henares, middle Chalcolithic phase; 24: Soto de Henares; Chalcolithic–Bronze phase; 25: Aldovea, domestic contexts; 26: Aldovea deposits, BCDom: Barrio del Castillo domestic contexts, BCDep: Barrio del Castillo deposits.
Ritual deposits from other sites, such as Aldovea [4], exhibit similar mortality patterns to those found at Barrio del Castillo, with a prevalence of infant dogs and pigs, as well as adult cattle. These patterns contrast with those observed in the domestic contexts of Barrio del Castillo, Aldovea [4], El Barranco del Herrero [33], Las Cabeceras [34], Loma Chiclana [1], Humanejos [35], Aguas Vivas (except for pigs) [36,37], and Zanjillas [3], the last of which shows an adult predominance across all taxa. High frequencies of non-adult caprines and pigs are seen in a limited number of sites, such as El Capricho [1], El Ventorro [1,38,39], and Camino de Yeseras [5]. Similarly, Aguas Vivas and Zanjillas present higher frequencies of non-adult pigs [3,36], while El Capricho stands out for its higher representation of non-adult cattle [1].
The mortality patterns observed at Barrio del Castillo, which show a clear predominance of adult caprines and cattle, suggest a livestock management system focused on optimizing animal resources throughout their lifespans. In this system, meat production appears to be secondary, with greater emphasis placed on secondary products, such as dairy and wool from caprines, as well as dairy products and labor from cattle. These patterns are also observed at other sites in the middle Tagus Valley [1,3,4,5,33,34,36,37]. The consistent importance of cattle across these sites highlights their key role in the economies of these societies. Not only are they one of the principal species in all of these sites, but they are also typically among the most frequent species found in ritual deposits, behind only dogs and pigs [4], with the distinction that they are generally represented by adults rather than infants.
Lastly, drawing on data from sites such as Aldovea [4] and Camino de las Yeseras [5], the evidence from Barrio del Castillo clearly highlights the significant role of dogs within Chalcolithic societies. Their high representation in ritual deposits suggests a symbolic importance, likely connected to their functional roles as hunting companions or as guardians and protectors within settlements.
Nevertheless, aside from the likely symbolic and ritualistic importance of dogs, it is entirely possible that dogs were used in complementary activities to hunting and herding. In sites like Aguas Vivas [37], cut marks have been documented on dog bones, which could suggest an exploitation of the animals for meat. Evidence like this has not been well registered in other sites along the middle Tagus Valley, presumably due to the poor preservation of bone surfaces in Barrio del Castillo, Zanjillas [3], Aldovea [4], and Camino de las Yeseras [5].
In any case, dogs seem to have a great significance for these societies and, although Chalcolithic communities had, by this time, largely consolidated agricultural and pastoral productive systems, the continued symbolic prominence of dogs may point to a cultural continuity rooted in earlier ways of life, in which hunting played a key role for sustenance. This symbolic association appears to diminish in later periods; evidence from other sites shows that, during the Bronze Age, the dominance of dogs in ritual contexts is gradually replaced by cattle, caprines, and pigs [4,40]. This shift may reflect broader socio-economic transformations: as agrarian and herding systems became increasingly dominant, the importance of hunting declined, along with the symbolic relevance of dogs.
This trend is also reflected in taxonomic patterns, where the frequency of wild game decreases progressively from the Chalcolithic to the Iron Age [4,35,41,42,43,44,45]. This symbolic transition is further suggested by the increased presence of caprines and cattle in ritual deposits during the Bronze and Iron Ages [3,4,40], which may indicate a shift in cultural values and ritual practices.
Other interpretations have also been proposed, including the notion of dogs as part of foundational or commemorative deposits [46,47,48]. In any case, regardless of the specific symbolic interpretation, the data from Barrio del Castillo—as well as other contemporary sites—strongly suggest that ritual deposits held significant symbolic meaning for the Chalcolithic communities of the middle Tagus Valley.
6. Conclusions
Predominance of Domestic Fauna: Across the entire assemblage, domestic fauna are significantly more prevalent than wild fauna. Game species are represented by less than 10% of the minimum number of individuals (MNI), a pattern that aligns with data from other Chalcolithic and Bronze Age sites in the interior of the Iberian Peninsula [1,2,4,10,49,50,51].
Predominance of Caprines and Cattle in Domestic Contexts: Domestic assemblages are primarily composed of caprines (sheep and goats) and cattle. In contrast, other domestic species, such as pigs and dogs, appear less frequently, typically accounting for under 10% of the assemblage. This contrasts sharply with ritual deposits, where dogs dominate during the Chalcolithic period, and pigs and cattle become more prominent in the Bronze age. These patterns are consistent with findings from the Chalcolithic and Bronze Age sectors of Aldovea [4,10], where dog and pig deposits decrease in later phases, while caprines—and especially cattle—gain symbolic and quantitative importance. A similar trend is observed in the Protocogotas Bronze age contexts presented in [10].
Symbolism and Mortality Patterns: The present taxonomic patterns, in conjunction with the mortality profiles, reinforce the symbolic nature of the ritual deposits. In Chalcolithic assemblages, the presence of infant pigs and dogs clearly suggests intentional, symbolic deposition. In contrast, adult cattle—present in both domestic and ritual contexts throughout the Chalcolithic and Bronze Ages—may have played a distinct symbolic role, perhaps different from those of pigs and dogs. In the Bronze Age, the marked increase in adult cattle, as seen in Aldovea [10] and the Protocogotas phase of Barrio del Castillo, points to a growing economic and symbolic significance of cattle.
The mortality profiles further underscore the symbolic distinctions between assemblages. Ritual deposits from the Chalcolithic phase include infant individuals (pigs and dogs) and adult cattle, whereas domestic contexts across all taxa exhibit a consistent pattern of adult individuals. The recurring presence of adult cattle in both domestic and ritual contexts suggests a unique and enduring symbolic status for this species within Chalcolithic and Bronze Age communities in the middle Tagus Valley.
7. Future Proposals and Final Remarks
This article presents the documentation of a new Chalcolithic settlement, featuring two clearly differentiated types of faunal assemblages. Domestic contexts at Barrio del Castillo are overwhelmingly dominated by caprines and cattle. This pattern aligns with that observed at numerous sites across the inner Iberian Peninsula, including Aguas Vivas [36,37], La Esgaravita [52], Las Matillas [8], Camino de las Yeseras [2,5,53], Aldovea [4], El Capricho [1], Loma de Chiclana [1], El Tejar del Sastre [1], Las Cabeceras [34], El Espinillo [41], El Ventorro [38], El Barranco del Herrero [33], Juan Barbero [54], Zanjillas [3], Huerta de los Cabreros [55], Soto de Henares [6], Angosta de los Mancebos [56], and La Loma del Lomo [7]. Nevertheless, many of these sites also feature assemblages that may be interpreted not as domestic refuse, but as ritual deposits, with distinct taxonomic compositions.
Ritual deposits have been the focus of various analyses e.g., [2,4,7,10,17,49,51], revealing unique and non-random taxonomic patterns, especially in the Chalcolithic phase. Dogs and pigs are often highly represented in these contexts, yet many aspects of their symbolic significance remain underexplored. Further analysis is needed, both through the study of additional sites and through focused investigations into the symbolic and cultural dimensions of these deposits.
The data from Barrio del Castillo contribute to the growing corpus of Chalcolithic ritual faunal assemblages in the Iberian interior, complementing and expanding upon previous studies, such as [2,4]. As documented in Aldovea [4,10], there appears to be a diachronic shift in the symbolic roles of animals: dogs, predominant in early ritual deposits, lose prominence by the end of the Chalcolithic phase, giving way to pigs, caprines, and cattle in the Bronze Age. This transformation may reflect the increasing consolidation of agricultural and herding systems, as well as the corresponding decline of hunting as both a subsistence strategy and symbolic practice in the middle Tagus Valley.
However, these patterns must be confirmed with additional data. While the progression observed at Aldovea suggests a broader regional trend, a more robust and nuanced understanding requires further zooarchaeological studies across a wider range of sites. Only through a more comprehensive and comparative approach can we begin to fully grasp the cultural significance of these animal deposits and their roles in the symbolic landscapes of prehistoric Iberian societies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/heritage8050181/s1, Supplementary File contains the the orthophoto of Barrio del Castillo and the skeletal profile data of domestic contexts from Chalcolithic levels of Barrio del Castillo.
Author Contributions
Conceptualization, V.E.-G. and J.Y.; data curation, V.E.-G. and J.Y.; formal analysis, V.E.-G., J.C.-M. and J.Y.; investigation, V.E.-G. and J.C.-M.; methodology, V.E.-G. and J.Y.; project administration, M.M.-G.; supervision, V.E.-G., M.M.-G. and J.Y.; validation, V.E.-G., M.M.-G. and J.Y.; visualization, V.E.-G., M.M.-G. and J.Y. writing—original draft, V.E.-G., J.C.-M. and J.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The samples are in the Regional Archaeological Museum of Madrid.
Acknowledgments
We thank the Complutense University of Madrid for the support provided to carry out this research and appreciate the suggestions of the anonymous reviewers. We are grateful for the opportunity, provided by the company Áqaba Arqueólogos S.L., to conduct a zooarchaeological study of this site in 2018–2022. We also want to express our appreciation to the journal Heritage for offering us the chance to publish this research.
Conflicts of Interest
Author Mónica Major-González is employed by the company Áqaba Arqueólogos S.L.
References
- Morales, A.; Liesau von Lettow-Vorbeck, C. Arqueozoología del Calcolítico en Madrid. In El Horizonte Campaniforme de la región de Madrid en el Centenario de Ciempozuelos; Blasco, C., Ed.; Ensayo Crítico de Síntesis: Madrid, Spain, 1994; pp. 227–247. [Google Scholar]
- Liesau von Lettow-Vorbeck, C. Fauna in Living and Funerary Contexts of the 3rd Millennium BC in Central Iberia. In Key Resources and Sociocultural Developments in the Iberian Chalcolithic; Bartelheim, M., Bueno Ramirez, P., Kunst, M., Eds.; Academia: Berlin, Germany, 2017; pp. 107–128. [Google Scholar]
- Estaca-Gómez, V.; López, G.; Morín, J.; Yravedra, J. Estudio Zooarqueológico y Tafonómico del Yacimiento Calcolítico Las Zanjillas (Torrejón de Velasco, Madrid). Munibe Antropoligía-Arkeol. 2023, 74, 87–104. [Google Scholar] [CrossRef]
- Estaca Gómez, V.; de la Torre García, A.; Señoran Martín, J.M.; Martínez Granero, A.B.; Major González, M.; Yravedra Sainz de los Terreros, J. Aprovechamiento de recursos animales en el yacimiento calcolítico precampaniforme de Aldovea (Torrejón de Ardoz, Madrid). Complutum 2023, 34, 31–55. [Google Scholar] [CrossRef]
- Liesau von Lettow-Vorbeck, C. La Arqueozoología, un elemento clave en la concepción espacial de Camino de las Yeseras. Los restos de mamíferos del ámbito doméstico y funerario. In Yacimientos Calcolíticos con Campaniforme de la región de Madrid. Nuevos Estudios; Blasco, C., Liesau, C., Ríos, P., Eds.; Patrimonio Arqueológico de Madrid, 6; Universidad Autónoma de Madrid: Madrid, Spain, 2011; pp. 167–170. [Google Scholar]
- Galindo-Pellicena, M.Á.; San José, V.; Sánchez, M.; Sánchez, M.; Lorente, M. Soto del Henares. Aproximación a un poblado de recintos. In Actas de las Cuartas Jornadas de Patrimonio Arqueológico de la Comunidad de Madrid; Jordana, N.B., Benito López, J.E., Eds.; Museo Arqueológico Regional: Alcalá de Henares, Spain, 2009; pp. 263–271. [Google Scholar]
- Valiente Malla, J. La Loma del Lomo III, Cogolludo Guadalajara; Patrimonio Historico; Arqueología: Castilla la Mancha, Spain, 2001; Volume 17, 304p. [Google Scholar]
- Díaz del Río, P. Campesinado y gestión pluriactiva del ecosistema: Un marco teórico para el análisis del III y el II milenios a.C. en la Meseta peninsular. Trab. Prehist. 1995, 54, 99–109. [Google Scholar] [CrossRef]
- Major-González, M.; Señorán Martín, J.M.; López-López, G.; Martínez Granero, A.B.; Izquierdo Zamora, A. Los yacimientos prehistóricos de Aldovea; Actas de la RAM: Torrejón de Ardoz, Madrid; Madrid, Spain, 2019; pp. 50–59. [Google Scholar]
- Estaca Gómez, V.; de la Torre, A.; Tardaguila-Giacomozzi, S.; Major González, M. Zooarchaeological study of Aldovea (Torrejón de Ardoz, Madrid), a new Bronze Age archaeological site from the Iberian Peninsula inland. J. Archaeol. Sci. Rep. 2024, 57, 104616. [Google Scholar] [CrossRef]
- Blázquez-Orta, R.; Rodríguez, L.; Major González, M.; Estaca-Gómez, V.; De Gaspar, I.; Feranec, R.S.; Carretero, J.M.; Arsuaga, J.L.; García, N. Dogs from the past: Exploring morphology in mandibles from Iberian archaeological sites using 3D geometric morphometrics. J. Archaeol. Sci. Rep. 2024, 57, 104660. [Google Scholar] [CrossRef]
- Bellido, A. Los campos de hoyos. In Inicios de la Economía Agrícola en la Submeseta Norte; Studia Archaeológica, 85; Universidad de Valladolid: Valladolid, Spain, 1996. [Google Scholar]
- Díaz del Río, P. La Formación del Paisaje Agrario: Madrid en el III y II Milenios BC; Arqueología, Paleontología Y Etnografía, 9; Comunidad de Madrid: Madrid, Spain, 2001; 397p, Available online: https://www.madrid.org/bvirtual/BVCM002085.pdf (accessed on 16 May 2025).
- Díaz del Río, P. Recintos de fosos del III milenio ac en la meseta peninsular. Trab. Prehist. 2003, 60, 61–78. [Google Scholar] [CrossRef]
- Alameda Cuenca-Romero, M.C.; Carmona Ballestero, E.; Pascual Blanco, S.; Martínez Díez, G.; Díez Pastor, C. El “campo de hoyos” calcolítico de Fuente Celada (Burgos): Datos preliminares y perspectivas. Complutum 2011, 22, 47–69. [Google Scholar] [CrossRef]
- González, J.M. Interpretación arqueológica de un “Campo de Hoyos” en Forfoleda (Salamanca). Zephyrus 2009, 46, 309–313. [Google Scholar]
- Barone, R. Anatomie Comparée des Mammifères Domestiques 1; Ostéologie-Paris Laboratoire d’Anatomie, Ecole Nationale Vétérinaire: Paris, France, 1986; 879p. [Google Scholar]
- Pales, L.; Lambert, C. Atlas Osteologique Pour Servir à l’Identification des Mamiferes du Quaternaire; Éditions du Centre national de la Recherche Scientifique: Paris, France, 1971; 177p. [Google Scholar]
- Prummel, W. Distinguishing Features en Postcranial SKELETAL elememnts of Cattle, Bos primigenius, Bos taurus and Red deer, Cervus elaphus; Schriften aus der Archäologisch-Zoologischen Arbeitsgruppe Schleswig-Kiel; Heft 12; Institut für Haustierkunde Neue Universität: Paris, France, 1988; pp. 5–52. [Google Scholar]
- Schmid, E. Atlas of Animal Bones for Prehistorians, Archaeologist and Quaternary Geologist; Elsevier Publishing Company: Amsterdan, The Netherland; London, UK; New York, NY, USA, 1972; 156p. [Google Scholar]
- Hilson, S. Mammal Bones and Teeth: An Introductory Guide to Methods of Identification; London Institute of Archaeology: London, UK, 1992; 119p. [Google Scholar]
- Boesseneck, J. Osteological Differences between Sheep (Ovis aries Linné) and Goats (Capra hircus Linné). In Science in Archaeology; Praeger: New York, NY, USA, 1969; pp. 331–358. [Google Scholar]
- Fernández, H. Osteologie Comparee des Petties Ruminants Eurasiatiques Sauvages et Domestiques (Genres Rupicapra, Ovis, Capra et Capreolus): Diagnose Differentialle du Squelette Apendiculaire. Ph.D. Thesis, Facultat de Ciencies, Universite de Geneva, Geneva, Switzerland, 2001. [Google Scholar] [CrossRef]
- Payne, S. Morphological distinction between the mandibular teeth of young sheep, Ovis and goats, Capra. J. Archaeol. Sci. 1985, 12, 139–147. [Google Scholar] [CrossRef]
- Prummel, W.; Frisch, H.J. A guide for the distinction of species, sex and body size in bones of sheep and goat. J. Archaeol. Sci. 1986, 13, 567–577. [Google Scholar] [CrossRef]
- Rolett, B.V.; Chiu, M. Age Estimation of Prehistoric Pigs (Sus scrofa) by Molar Eruption and Attrition. J. Archaeol. Sci. 1994, 21, 377–386. [Google Scholar] [CrossRef]
- Pérez Ripoll, M. Estudio de la secuencia del desgaste de los molares de Capra pyrenaica de los yacimientos prehistóricos. Arch. Prehist. Levantina 1988, 18, 83–128. [Google Scholar]
- Payne, S. Kill-off pattern in sheep and goats: The mandibles of Açvan Kale. Anatol. Stud. 1973, 23, 281–303. [Google Scholar] [CrossRef]
- Ducos, P. L’Origine des Animaux Domestiques en Palestine. Publications de l’Institut de Pr´ehistoire de l’Universite´ de Bordeau/M´emoire, Volume 6; Delmas: Bordeaux, France, 1968; 191p. [Google Scholar]
- Grant, A. The use of tooth wear as a guide to the age of domestic ungulates. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; BAR International Series 109; BAR Publishing: Oxford, UK, 1982; pp. 91–108. [Google Scholar]
- O’Connor, T.P. Husbandry decisions: Age at death. In The Analysis of Urban Animal Bones Assemblages: A Hand Book for Archaeologists; O’Connor, T.P., Ed.; Council for British Archaeology: York, UK, 2003; pp. 157–165. [Google Scholar]
- Yravedra, J. Tafonomía Aplicada a Zooarqueología; Aula Abierta: Madrid, Spain, 2006; 412p. [Google Scholar]
- Yravedra, J. Arqueozoología y Tafonomía del Yacimiento Calcolítico del Barranco del Herrero (San Martín de la Vega, Madrid). Estud. Prehist. Arqueol. Madrileñas 2007, 14–16, 427–440. [Google Scholar]
- Moreno-García, M.; y Cantalapiedra, V. Sobre el aprovechamiento de recursos de origen animal en la región de Madrid durante el III milenio cal. AC: La fauna de los contextos calcolíticos del Sector 3 de Las Cabeceras (Pozuelo de Alarcón, Madrid). Boletín Semin. Estud. Arte Arqueol. (BSAA) 2021, LXXXV–LXXXVI, 177–218. [Google Scholar] [CrossRef]
- Estaca Gómez, V.; Yravedra, J. Informe Zooarqueológico de Humanjos; Parla: Comunidad de Madrid, Spain, 2015; (Unpublished). [Google Scholar]
- García Somoza, P. Zooarqueología de los sectores 0 y Vía Pecuaria del yacimiento Ampliación Aguas Vivas. In El yacimiento arqueológico de Aguas Vivas. Prehistoria Reciente en el Valle del Río Henares (Guadalajara); Cantalapiedra-Jiménez, V., Ísmodes-Ezcurra, A., Eds.; La Ergástula Ediciones: Madrid, Spain, 2010; Volume 1, pp. 133–161. [Google Scholar]
- Yravedra, J. Zooarqueología de los sectores 1 y 2 del Yacimiento Ampliación Aguas Vivas. In El yacimiento arqueológico de Aguas Vivas. Prehistoria Reciente en el Valle del Río Henares (Guadalajara); Cantalapiedra-Jiménez, V., Ísmodes-Ezcurra, A., Eds.; La Ergástula Ediciones: Madrid, Spain, 2010; Volume 1, pp. 121–132. [Google Scholar]
- Priego, Mª.C.; y Quero, S. El Ventorro, un Poblado Prehistórico de los Albores de la Metalurgia; Estudios de Prehistoria y Arqueología Madrileñas: Madrid, Spain, 1992; Volume 8, 381p. [Google Scholar]
- Morales, A.; Villegas, C. La fauna de mamíferos del yacimiento de “El Ventorro”. Pecado-tesis osteológica de la campaña de 1981. Estud. Prehist. Arqueol. Madrileñas 1994, 9, 35–56. [Google Scholar]
- Liesau von Lettow-Vorbeck, C. Depósitos con ofrendas de animales en yacimientos Cogotas I: Antecedentes y características. In Cogotas I, Una cultura de la Edad del Bronce en la Península Ibérica; Rodríguez Marcos, J.A., Fernández Manzano, J., Eds.; Ediciones Universidad de Valladolid: Valladolid, Spain, 2012; pp. 219–257. [Google Scholar]
- Cerdeño, M.E.; Herráez, E. Estudio de la fauna del Yacimiento del Espinillo (Villaverde, Madrid). In El Espinillo: Un Yacimiento Calcolítico y de la edad del Bronce en las Terrazas del Manzanares; Baquedano, E., de la Torre, M.P., Marín, A.B., Eds.; Arqueología, Paleontología y Etnografía: Madrid, Spain, 2000; Volume 8, pp. 141–149. [Google Scholar]
- Baquedano, M.L.; Blanco, F.; Alonso, P.; Álvarez, D. El Espinillo: Un Yacimiento Calcolítico y de la Edad del Bronce en las Terrazas del Manzanares; Arqueología, Paleontología y Etnografia: Madrid, Spain, 2000; Volume 8, 164p. [Google Scholar]
- Estaca-Gómez, V. La Zooarqueología Durante la Edad del Hierro en el Valle Medio del Tajo; AUDEMA; UCM: Madrid, Spain, 2017; 328p. [Google Scholar]
- Estaca-Gómez, V. Prácticas socioeconómicas de la fauna doméstica de la Edad del Hierro en el Valle Medio del Tajo. Complutum 2018, 29, 381–406. [Google Scholar] [CrossRef]
- Estaca-Gómez, V.; Linares-Matás, G.J. Husbandry practices among Iron Age communities in the centre of the Iberian Peninsula. Archaeol. Anthropol. Sci. 2019, 11, 5009–5012. [Google Scholar] [CrossRef]
- Cámara, J.A.; Lizcano, R. Ritual y sedentarización en el yacimiento del Polideportivo de Martos (Jaén). Actas del I Congrés del Neolitic a la Península Ibérica, Gavá-Bellaterra, 1995. Rubricantum 1996, 1, 313–322. [Google Scholar]
- Liesau, C.; Vega, J.; Daza, A.; Ríos, P.; Menduiña, R.; Blasco, C. Manifestaciones simbólicas en el acceso Noreste del Recinto 4 de Foso en Camino de las Yeseras (San Fernando de Henares, Madrid). Saldvie 2014, 13/14, 53–69. [Google Scholar] [CrossRef]
- Márquez-Romero, J.E. Sobre los depósitos estructurados de animales en yacimientos de fosos del Sur de la Península Ibérica. Animais na Pré-hisória e Arqueología da Península Ibérica. In Proceedings of the Actas do IV Congreso de Arqueología Peninsular, Braga, Portugal, 14–19 September 2004; pp. 15–25. [Google Scholar]
- Liesau, C.; Blasco, C.; Ríos, P.; Vega, J.; Menduiña, R.; Blanco, J.F.; Baena, J.; Herrera, T.; Petri, A.; Gómez, J.L. Un espacio compartido por vivos y muertos: El poblado calcolítico de fosos de Camino de las Yeseras (San Fernando de Henares, Madrid). Complutum 2008, 18, 97–120. [Google Scholar]
- Galindo-Pellicena, M.Á.; Pérez-Romero, A.; Gómez-Felipe, A.; Romero-Ruiz, M.; Blázquez-Orta, R.; Andreu-Alarcón, S.; Benítez de Lugo Enrich, L. Animals for the Deceased: Zooarchaeological Analysis of the Bronze Age in the Castillejo del Bonete Site (Terrinches, Ciudad Real, Spain). Animals 2025, 15, 680. [Google Scholar] [CrossRef]
- Estaca Gómez, V.; Cruz-Alcázar, R.; Tardaguila-Giacomozzi, S.; y Yravedra, J. New Evidence for the Bronze Age Zooarchaeology in the Inland Area of the Iberian Peninsula through the Analysis of Pista de Motos (Villaverde Bajo, Madrid). Animals 2024, 14, 413. [Google Scholar] [CrossRef] [PubMed]
- Martínez Navarrete, M.I. El yacimiento de La Esgaravita (Alcalá de Henares, Madrid) y la cuestión de los llamados fondos de cabaña del Valle del Manzanares. Trab. Prehist. 1979, 36, 83–118. [Google Scholar]
- Ríos, P.; Daza, A.; Ortiz, I.; de Chorro, M.Á.; y Liesau, C. La Cabaña ‘E’ del yacimiento de Camino de las Yeseras. Nuevos datos sobre el espacio doméstico en un Poblado de Hoyos. Cuad. Prehist. Arqueol. 2016, 2, 73–105. [Google Scholar]
- Molero Gutiérrez, G.; Brea López, P.; y Bustos Pretel, V. Estudio faunístico de la cueva de Juan Barbero (Tielmes, Madrid). Trab. Prehist. 1984, 41, 105–112. [Google Scholar]
- Liesau von Lettow-Vorbeck, C. Análisis faunístico de los yacimientos de “Huerta de los Cabreros”, “Cantera de la Flamenca” y “Puente Largo del Jarama” (Aranjuez, Madrid). In El Poblamiento desde el Neolítico Final a la Primera Edad del Hierro en la Cuenca Media del río Tajo; Muñoz, K., Ed.; Univesidad Complutense de Madrid: Madrid, Spain, 1998; pp. 1418–1444. [Google Scholar]
- Priego Fernández, C.M. El yacimiento de Angosta de los Mancebos. Nueva contribución al conocimiento de la Edad del Bronce madrileño. Estud. Prehist. Arqueol. Madrileños 1994, 9, 91–97. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).