
Exploitation of Pigs during the Late Medieval and Early Modern Period in Croatia

 ,
,
Abstract
1. Introduction
2. Materials and Methods
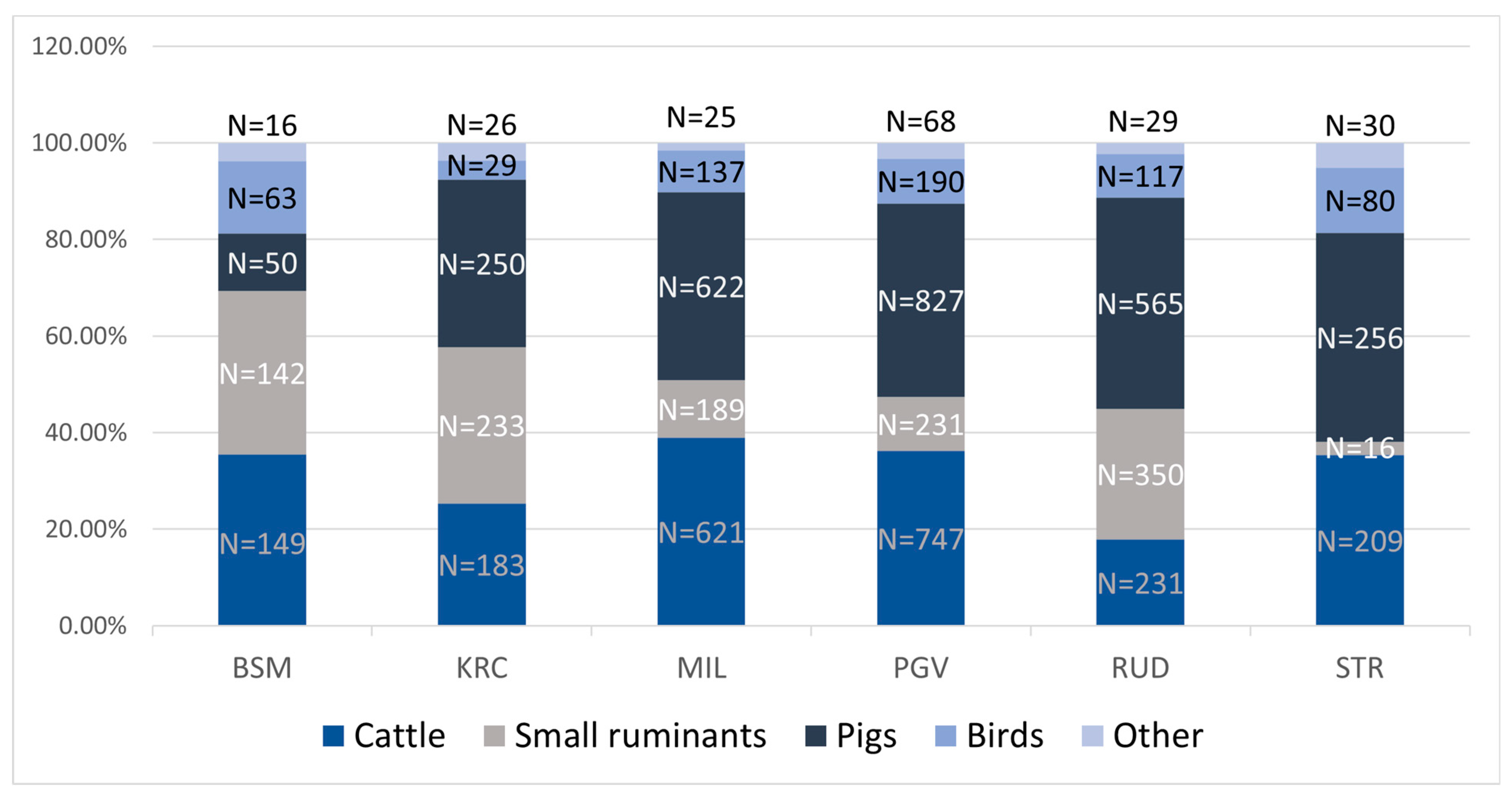
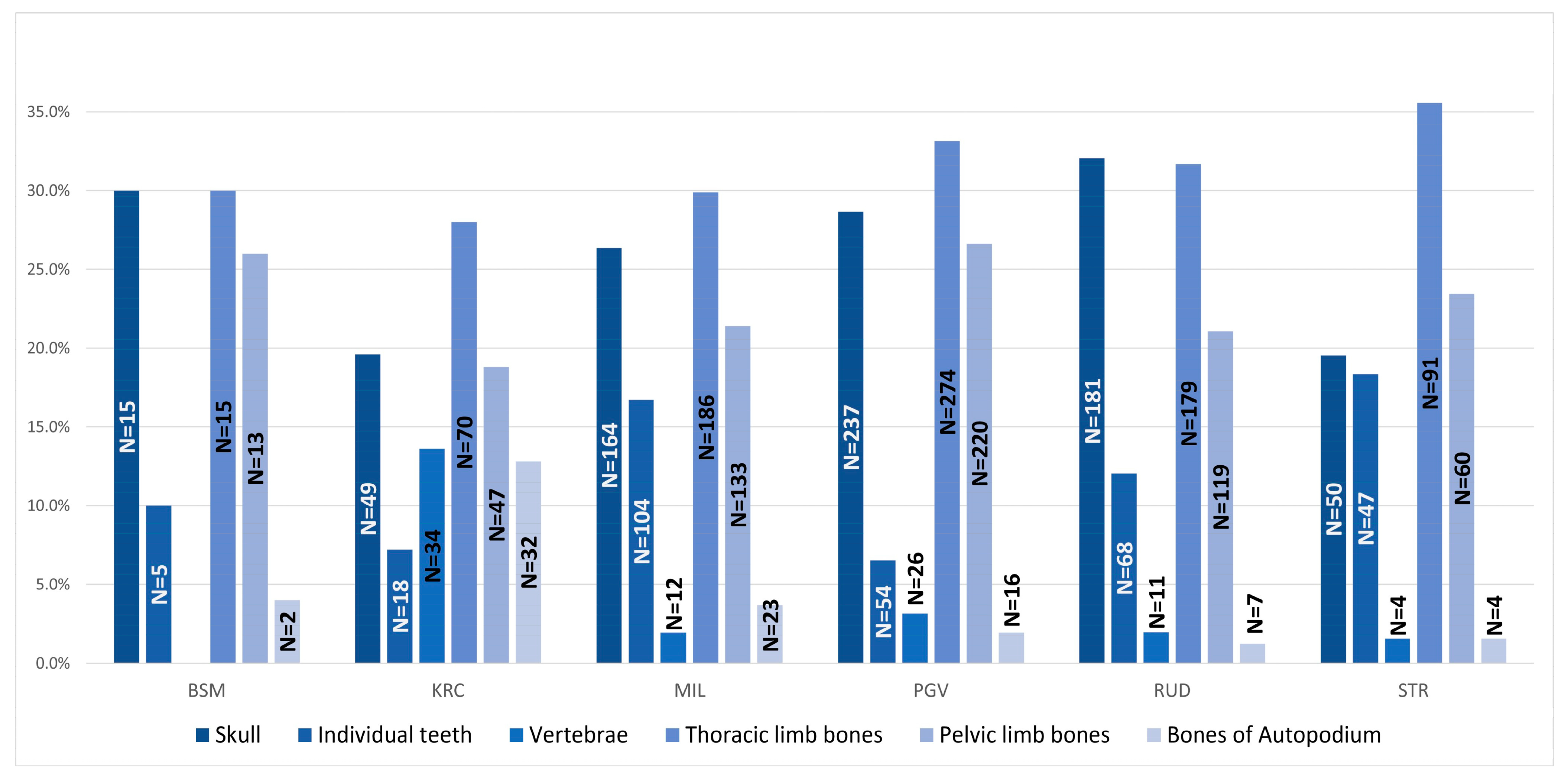
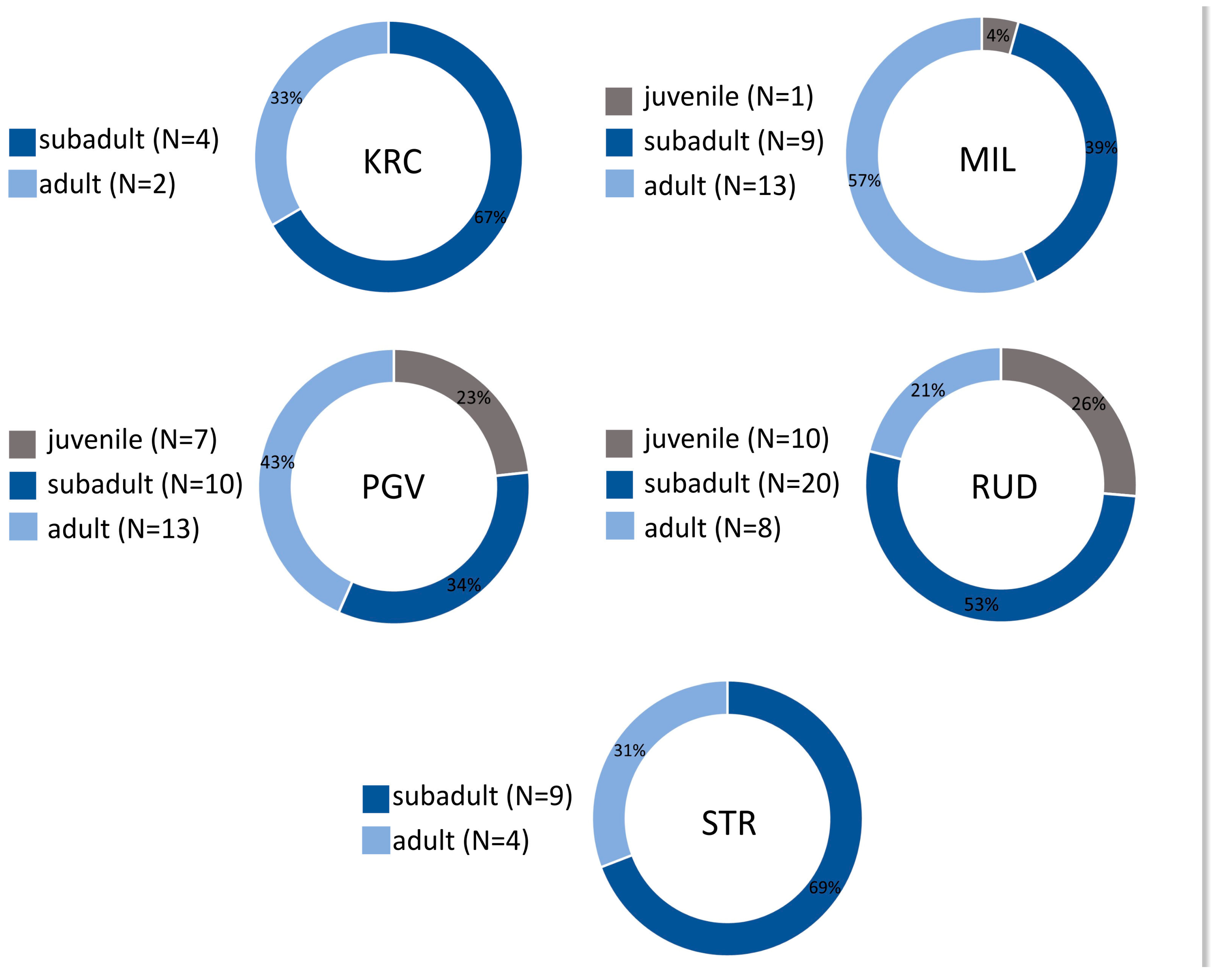
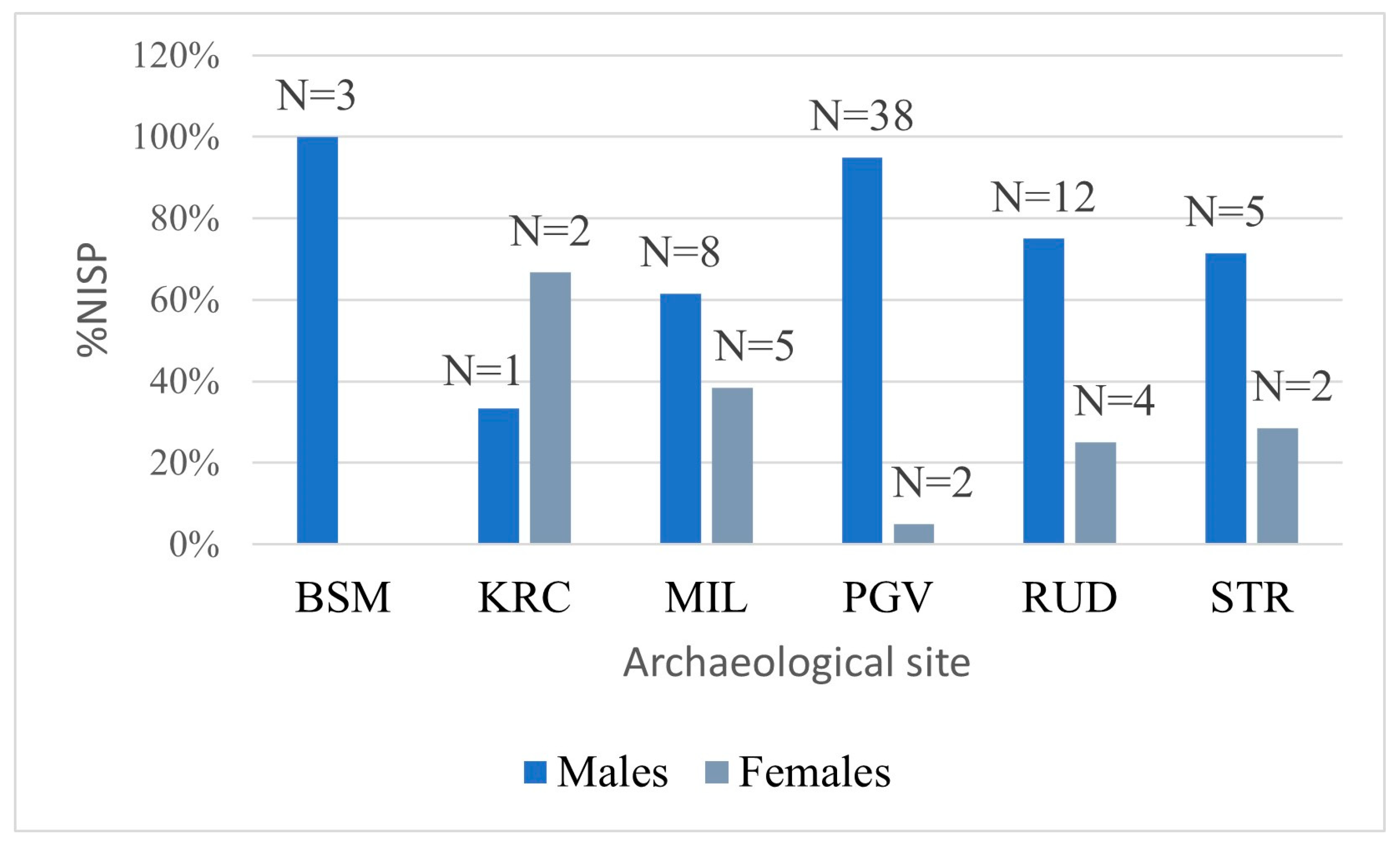
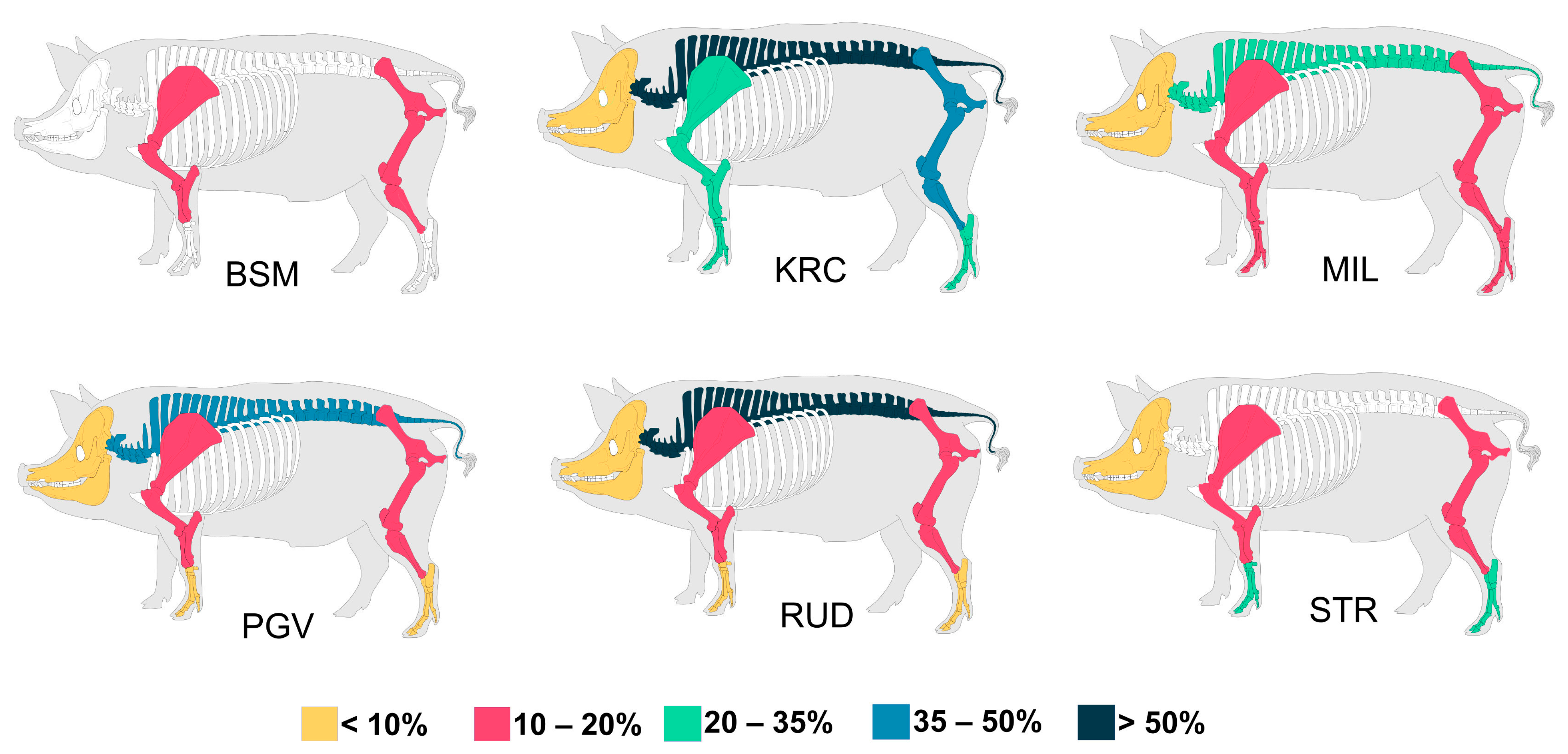
3. Results
4. Discussion
4.1. Diet in Medieval Croatia
4.2. Pork Consumption on European Sites
4.3. The Influence of Saint Benedictine Rule?
4.4. Pig Production Practices and Meat Consumption
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, A. Food, status and religion in England in the Middle Ages: An archaeozoological perspective. Anthropozoologica 1988, 2, 139–146. [Google Scholar]
- Ervynck, A.; Van Neer, W. Beef, pork and mutton: An archaeological survey of meat consumption in medieval and postmedieval towns in the southern Low Countries (Flanders & Brussels, Belgium). Quat. Int. 2017, 460, 65–73. [Google Scholar] [CrossRef]
- O’Connor, T.P. Provisioning urban communities: A topic in search of a model. Anthropozoologica 1992, 16, 101–106. [Google Scholar]
- Mladenović, T. Animal management in the Medieval Banat: Faunal remains from the settlement at the Pančevo—Livade site (Serbia). Mater. Și Cercet. Arheol. 2020, 16, 245–265. [Google Scholar] [CrossRef]
- Osypińska, M.; Żurawski, B.T. Pork for pilgrims: Livestock breeding and meat consumption at medieval Banganarti, Nubia. Antiquity 2020, 94, 164–178. [Google Scholar] [CrossRef]
- Wapnish, P.; Hesse, B. Urbanization and the organization of animal production at Tell Jemmeh in the Middle Bronze Age levant. J. Near East. Stud. 1988, 47, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Bartosiewicz, L. Animal Husbandry and medieval settlement in Hungary. Beitr. Mittelalterarchäol. Österr. 1999, 15, 139–155. [Google Scholar]
- Albarella, U.; Tagliacozzo, A.; Dobney, K.; Rowley-Conwy, P. Pig hunting and husbandry in Prehistoric Italy: A contribution to the domestication debate. Proc. Prehist. Soc. 2006, 72, 193–227. [Google Scholar] [CrossRef]
- Cesana, D.; Giovinazzo, R.; Marrazzo, D.; Melli, P.; Spinetti, A. The medieval diet in Genova (N.-W. Italy) through the analysis of faunal remains from archaeological sites. In Proceedings of the 4th International Congress of Medieval and Modern Archaeology—Medieval Europe, Paris, France, 3–8 September 2007; Harrassowitz Verlag: Leipzig, Germany, 2007; pp. 1–11. [Google Scholar]
- De Venuto, G. To eat and to be in medieval southern Italy: The zooarchaeological contribution from religious, rural, and fortified sites. In Bestial Mirrors—Using Animals to Construct Human Identities in Medieval Europe, Animals as Material Culture in the Middle Ages; Luskowski, A., Kunst, G.K., Kucera, M., Bietak, M., Hein, I., Eds.; Vienna Institute for Archaeological Science: Vienna, Austria, 2010; pp. 85–99. [Google Scholar]
- Albarella, U. Size, power, wool and veal: Zooarchaeological evidence for late medieval innovations. In Proceedings of the Papers of the “Medieval Europe Brugge 1997” Conference, Bruges, Belgium, 14–17 July 1997; Verhaeghe, F., Ed.; Instituut Voor Het Archeologisch Patrimonium: Zellik, Belgium, 1997; pp. 19–30. [Google Scholar]
- Rowley-Conwy, P.; Albarella, U.; Dobney, K. Distinguishing wild boar from domestic pig in Prehistory: A review of approaches and recent results. J. World Prehist. 2012, 25, 1–44. [Google Scholar] [CrossRef]
- Kužir, S.; Trbojević Vukičević, T. Životinjski nalazi s arheološkog lokaliteta Torčec—Gradić. Podravina 2004, 3, 116–121. [Google Scholar]
- Trbojević Vukičević, T.; Štilinović, A. Arheozoološka analiza kostiju s utvrde Čanjevo. In Utvrda Čanjevo–Istraživanja 2003–2007; Bekić, L., Ed.; Općina Visoko: Visoko, Croatia, 2008; pp. 253–259. [Google Scholar]
- Radović, S. Prehrana posade srednjovjekovne utvrde Paka. In Burg Paka; Šimek, M., Ed.; Gradski Muzej Varaždin, Državni Arhiv u Varaždinu: Varaždin, Croatia, 2021; pp. 215–242. [Google Scholar]
- Tkalčec, T.; Trbojević Vukičević, T. Archaeozoological evidence of dietary habits of small castle inhabitants in medieval Slavonia. In Castrum Bene 16, Castrum and Economy 124; Pisk, S., Ed.; Moslavina Museum Kutina, Historical Association Moslavina: Popovača, Croatia, 2021; pp. 124–152. [Google Scholar]
- Tkalčec, T. Burg Vrbovec u Klenovcu Humskome—Deset Godina Arheoloških Istraživanja; Horjan, G., Tomičić, Ž., Eds.; Muzeji Hrvatskog Zagorja, Institut za Arheologiju: Zagreb, Croatia, 2010; pp. 17–22. [Google Scholar]
- Sudec, K.; Hrelja, D. Milengrad—Zagonetna Utvrda; Državni arhiv u Varaždinu: Varaždin, Croatia, 2011; pp. 4–29. [Google Scholar]
- Kolar-Dimitrijević, M. Urbar pavlinskoga samostana u Strezi 1477. godine. Podravina 2003, 2, 103–123. [Google Scholar]
- Pleše, T.; Karlo, K. Monasterium omnium sanctorum de Ztreza ordinis s. Pauli primi eremitae. Opusc. Archaeol. 2009, 33, 183–205. [Google Scholar]
- Janeš, A. Arheološke Raziskave Benediktinskega Samostana sv. Margarete v Bijeli. Ph.D. Thesis, University of Ljubljana, Ljubljana, Slovenia, 2022. [Google Scholar]
- Janeš, A.; Sekulić, P. Rudina i Bijela: Benediktinski samostani kasnosrednjovjekovne Slavonije. Starohrv. Prosvj. 2014, 41, 185–204. [Google Scholar]
- Pleše, T.; Sekulić, P.; Mostarčić, B. Ususret četvrtom desetljeću od početka Istraživanja benediktinske opatije sv. Mihovila arkanđela na Rudini. Rad. Zavoda Znan. Umjetnički Rad Požegi 2018, 7, 91–135. [Google Scholar] [CrossRef]
- Kekez, H.; Pleše, T.; Sekulić, P. Plitvički Krčingrad i Babonići—Prilog kontekstualizaciji vremena gradnje. Povij. Pril. 2018, 37, 65–99. [Google Scholar]
- Miracle, P.T.; Pugsley, L. Vertebrate Faunal Remains from Pupićina Cave. In Prehistoric Herders of Northern Istria—The Archaeology of Pupićina Cave, 1st ed.; Miracle, P.T., Forenbaher, S., Eds.; Arheološki Muzej Istre—Pula: Pula, Croatia, 2006; pp. 259–400. [Google Scholar]
- Schmid, E. Atlas of Animal Bones for Prehistorians, Archaeologists, and Quaternary Geologists; Elsevier Publishing Company: Amsterdam, The Netherlands; London, UK; New York, NY, USA, 1972. [Google Scholar]
- Hillson, S. Teeth; Brothwell, D., Cunliffe, B., Fleming, S., Fowler, P., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 201–218. [Google Scholar]
- König, H.E.; Liebich, H.-G. Veterinary Anatomy of Domestic Animals, 7th ed.; Georg Thieme Verlag: Stuttgart, Germany; New York, NY, USA, 2020. [Google Scholar]
- Lyman, R.L. Quantitative Paleozoology; Cambridge University Press: New York, NY, USA, 2008; pp. 27–28. [Google Scholar]
- O’Connor, T.P. Bones from the General Accident Site, Tanner Row. In The Archaeology of York; Addyman, P.V., Ed.; Council for British Archaeology: London, UK, 1988; pp. 85–86. [Google Scholar]
- Reitz, E.J.; Wing, E.S. Zooarchaeology, 2nd ed.; Barker, G., Ed.; Cambridge University Press: New York, NY, USA, 2008; pp. 110–238. [Google Scholar]
- Statistica. Data Science Workbench, version 14.0.25; TIBCO Software Inc.: Palo Alto, CA, USA, 2020. [Google Scholar]
- Trbojević Vukičević, T.; Kučko, A. Procjena prehrambenih navika temeljem arheozoološke analize. In Stari Grad Barilović—10 Godina Arheoloških Istraživanja; Azinović Bebek, A., Krmpotić, M., Eds.; Hrvatski Restauratorski Zavod: Zagreb, Croatia, 2014; pp. 110–117. [Google Scholar]
- Adamček, J. Agrarni Odnosi u Hrvatskoj od Sredine XV do Kraja XVII Stoljeća; Sveučilišna Naklada Liber: Zagreb, Croatia, 1980. [Google Scholar]
- Kühtreiber, T. The medieval castle Lanzenkirchen in Lower Austria: Reconstruction of economical and ecological development of an average-sized manor (12th–15th century). Archaeol. Pol. 1999, 37, 135–144. [Google Scholar]
- Pasda, K. Tierknochen als Spiegel sozialer Verhältnisse im 8.–15. Jh. Bayerns. Ph.D. Thesis, Faculty of Science, Department of Geoscience, Eberhard Karls Universität Tübingen, Tübingen, Germany, 2003. [Google Scholar]
- Küchelmann, H.C. Noble meals instead of abstinence? A faunal assemblage from the Dominican Monastery of Norden, Northern Germany. In Proceedings of the General Session of the 11th International Council for Archaeozoology Conference, Pars, France, 23–28 August 2010; Lefèvre, C., Ed.; BAR Publishing: Oxford, UK, 2012; pp. 87–97. [Google Scholar]
- Albarella, U. Pig husbandry and pork consumption in Medieval England. In Food in Medieval England, 1st ed.; Woolgar, C.M., Serjeantson, D., Waldron, T., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2006; pp. 72–87. [Google Scholar]
- Powell, A.; Serjeantson, D.; Smith, P. Food consumption and disposal: The animal remains. In St. Gregory's Priory, Northgate, Canterbury. Excavations 1988–1991; Hicks, M., Hicks, A., Eds.; Canterbury Archaeological Trust: Canterbury, UK, 2001; pp. 289–333. [Google Scholar]
- Ayres, K.; Locker, A.; Serjeantson, D.; Hardy, A.; Dodd, A.; Keevill, C. Chapter 10: Mammal, bird and fish remains and oysters. In Aelfric’s Abbey: Excavations at Eynsham Abbey, Oxfordshire, 1989–1992; Hardy, A., Dodd, A., Keevill, C., Eds.; Oxford Archaeological Unit: Lancaster, UK, 2003; pp. 341–427. [Google Scholar] [CrossRef]
- Ervynck, A.; Van Neer, W.; Hüster Plogmann, H.; Schibler, J. Beyond affluence: The zooarchaeology of luxury. World Archaeol. 2003, 34, 428–441. [Google Scholar] [CrossRef]
- Gál, E. Remains of small domestic and game birds from medieval sites in Hungary. Quat. Int. 2020, 543, 99–107. [Google Scholar] [CrossRef]
- Gál, E. Archaeozoological assessment of the refuse deposit of the archiepiscopal residence in Esztergom. Antaeus 2021, 37, 421–472. [Google Scholar]
- Ginella, F.; Heigold-Stadelmann, A.; Ohnsorg, P.; Schibler, J. Ein Beitrag zur Nahrungswirtschaft und zur Verpflegung römischer Truppen im Legionslager Vindonissa/Windisch (CH). Archäozoologische Auswertung der Tierknochen aus der Grabung Vindonissa-Feuerwehrmagazin 1976. In Gesellschaft Pro Vindonissa, Jahresbericht 1999; Gesellschaft Pro Vindonissa: Brugg, Switzerland, 1999; pp. 3–26. [Google Scholar]
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: New York, NY, USA, 2001; pp. 294–314. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaeo Site/Species | Red Deer | Roe Deer | Hare | Squirrel | Vole | Horse | Bear | Cat | Dog | Mole | Ferret | Fox | Σ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BSM | 3 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 16 |
| KRC | 6 | 3 | 10 | 1 | 2 | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 26 |
| MIL | 2 | 5 | 14 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 25 |
| PGV | 12 | 20 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 4 | 68 |
| RUD | 7 | 7 | 5 | 0 | 0 | 8 | 1 | 1 | 0 | 0 | 0 | 0 | 29 |
| STR | 8 | 6 | 15 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 30 |
| Archaeological Site | NISP | NISP Pig | %NISP Pig |
|---|---|---|---|
| BSM | 414 | 50 | 12.2% |
| KRC | 722 | 250 | 34.6% |
| MIL | 1595 | 622 | 39% |
| PGV | 2079 | 827 | 39.8% |
| RUD | 1293 | 565 | 43.7% |
| STR | 591 | 256 | 43.3% |
| Archaeological Site | |||||||
|---|---|---|---|---|---|---|---|
| Bone | BSM | KRC | MIL | PGV | RUD | STR | Total |
| Mandible | 5 | 12 | 82 | 67 | 86 | 26 | 278 |
| Maxilla | 3 | 13 | 43 | 63 | 41 | 12 | 175 |
| Other | 7 | 24 | 39 | 107 | 54 | 12 | 243 |
| ΣCranial Bones | 15 | 49 | 164 | 237 | 181 | 50 | 696 |
| Individual Teeth | 5 | 18 | 104 | 54 | 68 | 47 | 296 |
| Vertebrae | 0 | 34 | 12 | 26 | 11 | 4 | 87 |
| Scapula | 3 | 13 | 37 | 70 | 27 | 18 | 168 |
| Humerus | 3 | 7 | 72 | 72 | 70 | 32 | 256 |
| Radius | 4 | 13 | 32 | 43 | 35 | 15 | 142 |
| Ulna | 5 | 15 | 35 | 38 | 36 | 19 | 148 |
| Ossa Carpi | 0 | 3 | 0 | 0 | 0 | 0 | 3 |
| MTC | 0 | 19 | 10 | 51 | 11 | 7 | 98 |
| ΣThoracic Limb Bones | 15 | 70 | 186 | 274 | 179 | 91 | 815 |
| Hip Bone | 5 | 9 | 21 | 57 | 27 | 17 | 136 |
| Femur | 4 | 11 + 1 * | 38 | 41 | 18 | 14 | 127 |
| Tibia | 3 | 11 | 42 | 61 | 48 | 20 | 185 |
| Fibula | 0 | 1 | 3 | 5 | 5 | 0 | 14 |
| Ossa Tarsi | 0 | 13 | 10 | 16 | 10 | 3 | 52 |
| MTT | 1 | 1 | 19 | 40 | 11 | 6 | 78 |
| ΣPelvic Limb Bones | 13 | 47 | 133 | 220 | 119 | 60 | 592 |
| Ph. prox. | 0 | 16 | 7 | 9 | 5 | 0 | 37 |
| Ph. med. | 0 | 2 | 7 | 3 | 1 | 0 | 13 |
| Ph. dist. | 0 | 3 | 1 | 0 | 0 | 0 | 4 |
| Metapodium | 2 | 11 | 8 | 4 | 1 | 4 | 30 |
| NISP | 50 | 250 | 622 | 827 | 565 | 256 | 2570 |
| Arch. Site/Age Group | Juvenile | Subadults | Adults | Total |
|---|---|---|---|---|
| BSM | 3 | 0 | 2 | 5 |
| KRC | 19 | 2 | 19 | 40 |
| MIL | 41 | 5 | 4 | 50 |
| PGV | 36 | 7 | 11 | 54 |
| RUD | 48 | 4 | 4 | 56 |
| STR | 9 | 2 | 3 | 14 |
| Total | 156 | 20 | 43 | 219 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korpes, K.; Piplica, A.; Đuras, M.; Trbojević Vukičević, T.; Kolenc, M. Exploitation of Pigs during the Late Medieval and Early Modern Period in Croatia. Heritage 2024, 7, 1014-1027. https://doi.org/10.3390/heritage7020049
Korpes K, Piplica A, Đuras M, Trbojević Vukičević T, Kolenc M. Exploitation of Pigs during the Late Medieval and Early Modern Period in Croatia. Heritage. 2024; 7(2):1014-1027. https://doi.org/10.3390/heritage7020049
Chicago/Turabian StyleKorpes, Kim, Aneta Piplica, Martina Đuras, Tajana Trbojević Vukičević, and Magdalena Kolenc. 2024. "Exploitation of Pigs during the Late Medieval and Early Modern Period in Croatia" Heritage 7, no. 2: 1014-1027. https://doi.org/10.3390/heritage7020049
APA StyleKorpes, K., Piplica, A., Đuras, M., Trbojević Vukičević, T., & Kolenc, M. (2024). Exploitation of Pigs during the Late Medieval and Early Modern Period in Croatia. Heritage, 7(2), 1014-1027. https://doi.org/10.3390/heritage7020049