
The High Potential of Micro-Magnetic Resonance Imaging for the Identification of Archaeological Reeds: The Case Study of Tutankhamun

 ,
,  ,
, 
 ,
, 

Abstract
:1. Introduction
Reed Anatomy and Its Study
2. Materials and Methods
2.1. Sample Preparation
2.2. Optical Microscopy (OM)
2.3. Micro-Magnetic Resonance Imaging (μ-MRI)
3. Results
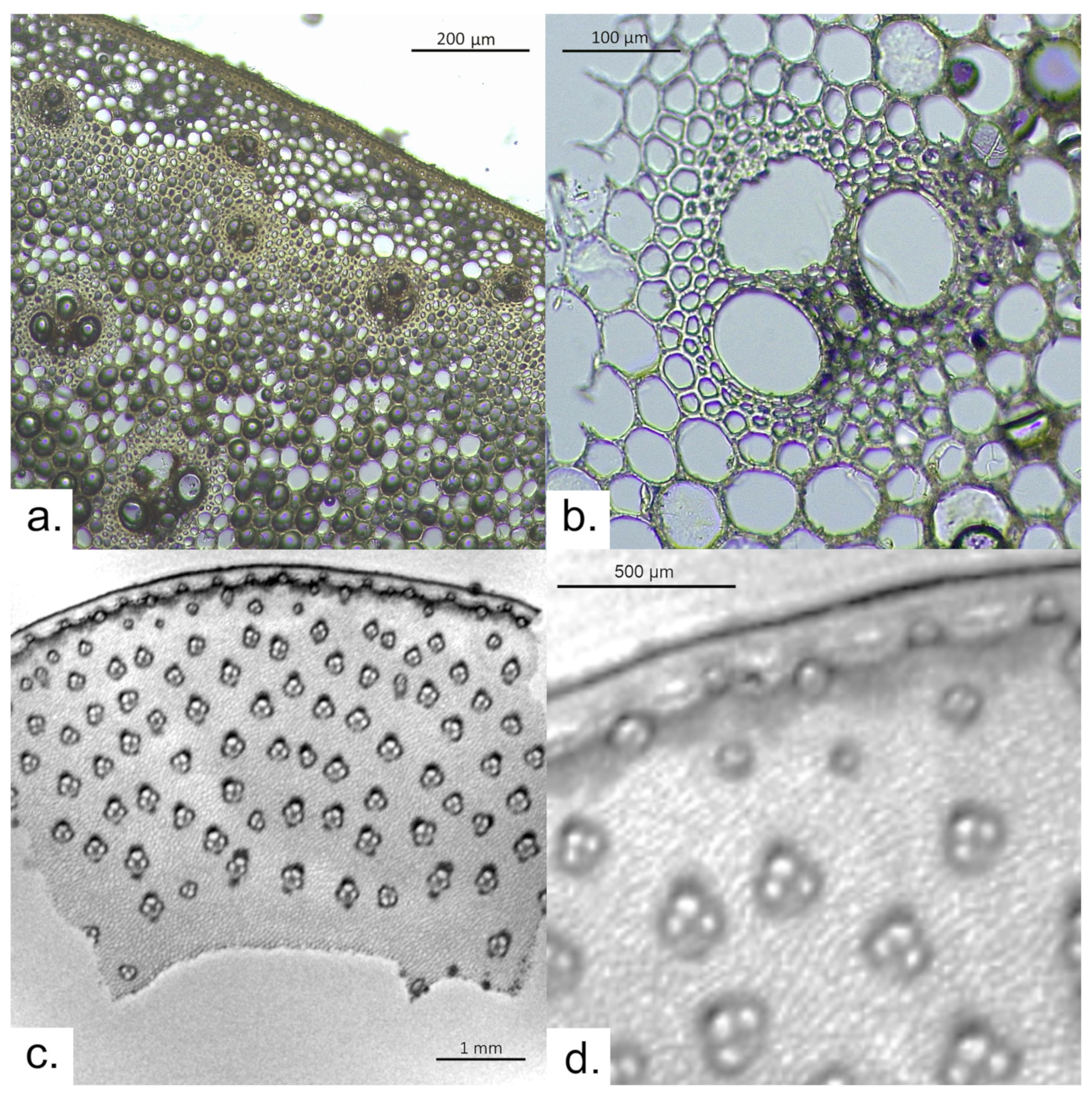
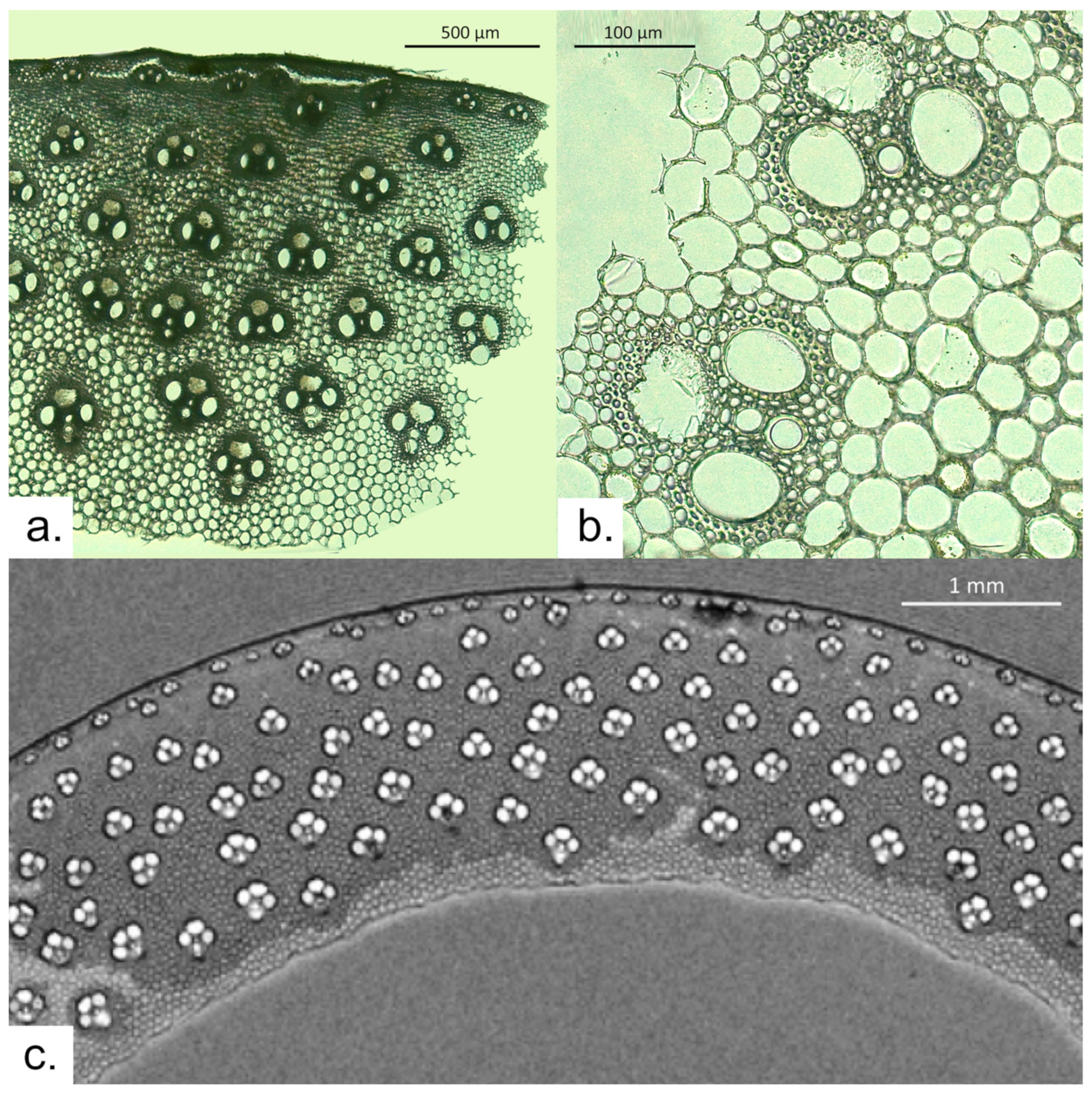
3.1. Arundo donax
3.2. Phragmites australis
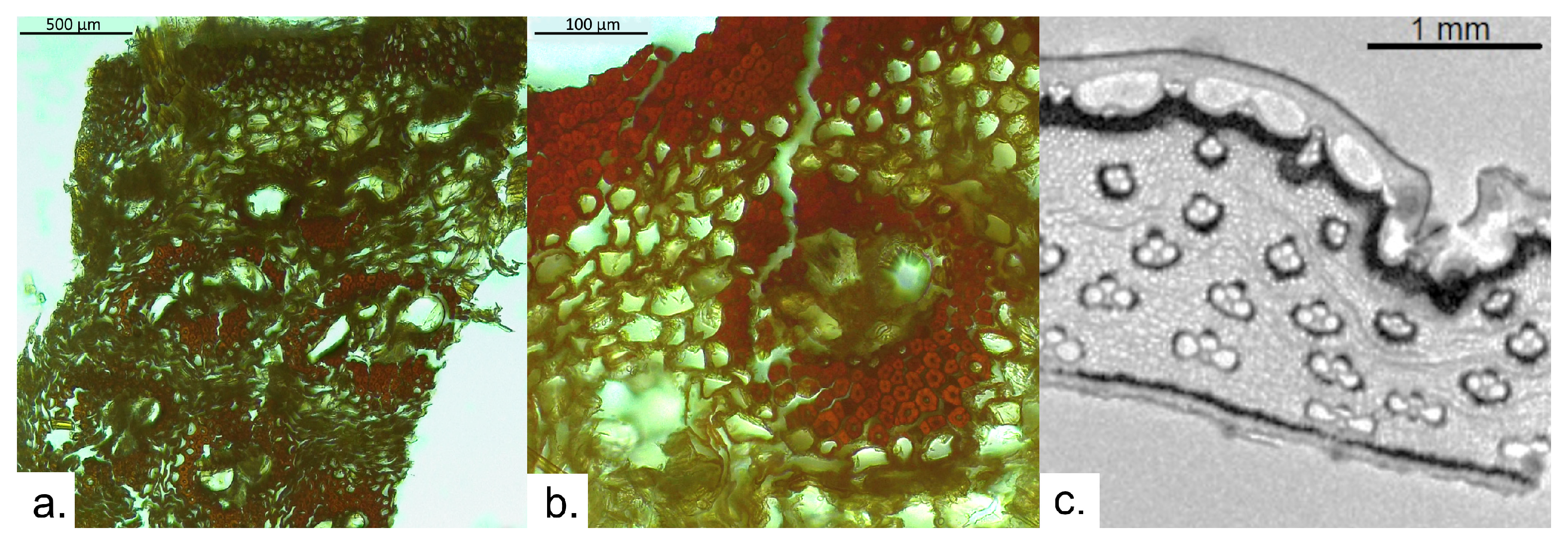
3.3. Archaeological Sample
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Kopold, P.; van Aken, P.A.; Maier, J.; Yu, Y. Energy storage materials from nature through nanotechnology: A sustainable route from reed plants to a silicon anode for lithium-ion batteries. Angew. Chem. 2015, 127, 9768–9772. [Google Scholar] [CrossRef]
- Enciclopedia Treccani. Canna. Enciclopedia Online. Available online: https://www.treccani.it/enciclopedia/canna#:~:text=Nome%20genericamente%20attribuito%20a%20numerose,che%20coniugano%20resistenza%20ed%20elasticit%C3%A0 (accessed on 10 September 2023).
- Ramsey, M.N.; Maher, L.A.; Macdonald, D.A.; Nadel, D.; Rosen, A.M. Sheltered by reeds and settled on sedges: Construction and use of a twenty-thousand-year-old hut according to phytolith analysis from Kharaneh IV, Jordan. J. Anthropol. Archaeol. 2018, 50, 85–97. [Google Scholar] [CrossRef]
- Oron, A.; Galili, E.; Hadas, G.; Klein, M. Early maritime activity on the Dead Sea: Bitumen harvesting and the possible use of reed watercraft. J. Marit. Archaeol. 2015, 10, 65–88. [Google Scholar] [CrossRef]
- Romano, L.; Celant, A.; Montorfani, M.V. Reed-swamps in the Sumerian material culture: Archaeological, archaeobotanical, and experimental insights from the Abu Tbeirah excavations. In Southern Iraq’s Marshes: Their Environment and Conservation; Jawad, L.A., Ed.; Springer: Cham, Switzerland, 2021; pp. 33–54. [Google Scholar]
- Kolesik, P.; Mills, A.; Sedgley, M. Anatomical characteristics affecting the musical performance of clarinet reeds made from Arundo donax L. (Gramineae). Ann. Bot. 1998, 81, 151–155. [Google Scholar] [CrossRef]
- Kawasaki, M.; Nobuchi, T.; Nakafushi, Y.; Nose, M.; Shiojiri, M. Structure and biomechanics of culms of Phragmites australis used for reeds of Japanese wind instrument “hichiriki”. Microsc. Res. Tech. 2015, 78, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Perdue, R.E. Arundo donax—Source of musical reeds and industrial cellulose. Econ. Bot. 1958, 12, 368–404. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Sadori, L.; Blasi, C. Editorial: Archaeobotany for cultural landscape and human impact reconstructions. Plant Biosyst. 2010, 144, 860–864. [Google Scholar] [CrossRef]
- Miksicek, C.H. Formation processes of the archaeobotanical record. Adv. Archaeol. Method Theory 1987, 10, 211–247. [Google Scholar]
- Boulos, L.; Fahmy, A.G.E.D. Grasses in ancient Egypt. Kew Bull. 2007, 62, 507–511. [Google Scholar]
- Noaman, S. Usage of Arundo donax L. as a sustainable material in interior design and architecture. In Cities’ Identity Through Architecture and Arts, Proceedings of the International Conference on Cities’ Identity through Architecture and Arts (CITAA 2017), Cairo, Egypt, 11–13 May 2017; Catalani, A., Nour, Z., Versaci, A., Hawkes, D., Bougdah, H., Sotoca, A., Ghoneem, M., Trapani, F., Eds.; Routledge: London, UK, 2018; p. 201. [Google Scholar]
- Kiviat, E. What reed (Phragmites) ecology tells us about reed management. Part 1. Confronting reed’s lurid reputation. News Hudsonia 2005, 20, 1–6. [Google Scholar]
- Thomas, E. Terrestrial marsh and solar mat. J. Egypt. Archaeol. 1959, 45, 38–51. [Google Scholar] [CrossRef]
- Barakat, H.N.; Aziz, A. Guide to Plants of Ancient Egypt; Bibliotheca Alexandrina: Alexandria, Egypt, 2010. [Google Scholar]
- Malek, J.; Moffett, J.; Hutchison, S.; Miles, E.; Magee, D.; Rawlinson, K.; Tutankhamun: Anatomy of an Excavation. Carter Archives—Main Object List: 001–049. Available online: http://www.griffith.ox.ac.uk/gri/carter/HomePage.html#042 (accessed on 15 September 2023).
- Abdrabou, A.; Abdallah, M.; Sultan, G.M.; Mostafa, M.; Bayoumi, H.; Magdy, R.; Abd El Kader, M.A.; Hamza, N.M.; Mamdouh, D.; Elsayed, H.M.; et al. Tutankhamun’s polychrome wooden shawabtis: Preliminary investigation for pigments and gilding characterization and indirect dating of previous restorations by the combined use of imaging and spectroscopic techniques. Open Archaeol. 2022, 8, 30–54. [Google Scholar] [CrossRef]
- Peruzzi, G.; Ciccola, A.; Bosi, A.; Serafini, I.; Negozio, M.; Hamza, N.M.; Moricca, C.; Sadori, L.; Favero, G.; Nigro, V.; et al. Applying Gel-Supported Liquid Extraction to Tutankhamun’s Textiles for the Identification of Ancient Colorants: A Case Study. Gels 2023, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Cutler, D.F.; Botha, C.E.J.; Stevenson, D.W. Plant Anatomy: An Applied Approach; Blackwell Publishing: Malden, MA, USA, 2008. [Google Scholar]
- Tian, S.; Yang, R.; Pan, Z.; Su, X.; Li, S.; Wang, P.; Huang, X. Anisotropic reed-stem-derived hierarchical porous biochars supported paraffin wax for efficient solar-thermal energy conversion and storage. J. Energy Storage 2022, 56, 106153. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Anatomy of European Woods; Haupt: Stuttgart, Germany, 1990. [Google Scholar]
- Martín-Seijo, M.; Teira-Brión, A.; Currás, A.; Rodríguez-Rellán, C. After the fire: The end of a house life-cycle at the Iron Age site of Nabás (North-western Iberia). Veg. Hist. Archaeobot. 2020, 29, 427–446. [Google Scholar] [CrossRef]
- Figueiral, I.; Bouby, L.; Buffat, L.; Petitot, H.; Terral, J.F. Archaeobotany, vine growing, and wine producing in Roman Southern France: The site of Gasquinoy (Béziers, Hérault). J. Archaeol. Sci. 2010, 37, 139–149. [Google Scholar] [CrossRef]
- Baruch, U. Identification of Charred Wood Remains from ‘En Gedi. Atiquot 2005, 49, 43–48. [Google Scholar]
- Stagno, V.; Moricca, C.; Sadori, L.; Dell’Aglio, E.; Reale, R.; Capuani, S. Evaluation of the efficacy of micro-Magnetic Resonance Imaging compared with light microscopy to investigate the anatomy of modern and ancient waterlogged wood. Magn. Reson. Imaging 2023, 102, 164–178. [Google Scholar] [CrossRef]
- Callaghan, P.T. Principles of Nuclear Magnetic Resonance Microscopy; Oxford University Press Inc.: New York, NY, USA, 1991. [Google Scholar]
- Cole-Hamilton, D.I.; Kaye, B.; Chudek, I.A.; Hunter, G. Nuclear magnetic resonance imaging of waterlogged wood. Stud. Conserv. 1995, 40, 41–50. [Google Scholar]
- Telkki, V.V. Wood Characterization by NMR & MRI of Fluids. eMagRes 2012, 1, 215–222. [Google Scholar]
- Longo, S.; Egizi, F.; Stagno, V.; Di Trani, M.G.; Marchelletta, G.; Gili, T.; Fazio, E.; Favero, G.; Capuani, S. A multi-parametric investigation on waterlogged wood using a magnetic resonance imaging clinical scanner. Forests 2023, 14, 276. [Google Scholar] [CrossRef]
- Germer, R. Die Pflanzenmaterialien aus dem grab des Tutanchamun; Gerstenberg Verlag: Hildesheim, Germany, 1989. [Google Scholar]
- El-Gawad, A.M.A.; El-Amier, Y.A. Anatomical features of three perennial swampy plants of Poaceae, grown on the water stream banks in Nile Delta, Egypt. J. Med. Bot. 2017, 1, 58–64. [Google Scholar] [CrossRef]
- Spatz, H.C.; Beismann, H.; Brüchert, F.; Emanns, A.; Speck, T. Biomechanics of the giant reed Arundo donax. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1997, 352, 1–10. [Google Scholar] [CrossRef]
- Yamauchi, T.; Shimamura, S.; Nakazono, M.; Mochizuki, T. Aerenchyma formation in crop species: A review. Field Crops Res. 2013, 152, 8–16. [Google Scholar] [CrossRef]
- Capuani, S.; Gambarini, G.; Guarnieri, R.; Di Pietro, G.; Testarelli, L.; Di Nardo, D. Nuclear magnetic resonance microimaging for the qualitative assessment of root canal treatment: An ex vivo preliminary study. Diagnostics 2021, 11, 1012. [Google Scholar] [CrossRef] [PubMed]
- Danin, A.; Naenny, W. Contribution to the recognition of reeds by their rhizome anatomy. Flora Mediterr. 2008, 18, 385–392. [Google Scholar]
- Armstrong, J.; Afreen-Zobayed, F.; Blyth, S.; Armstrong, W. Phragmites australis: Effects of shoot submergence on seedling growth and survival and radial oxygen loss from roots. Aquat. Bot. 1999, 64, 275–289. [Google Scholar] [CrossRef]
- Fogli, S.; Marchesini, R.; Gerdol, R. Reed (Phragmites australis) decline in a brackish wetland in Italy. Mar. Environ. Res. 2002, 53, 465–479. [Google Scholar] [CrossRef]
- Bleicher, N. An easy low-budget method to produce thin-sections of heavily decayed archaeological wood. Dendrochronologia 2008, 26, 9–11. [Google Scholar] [CrossRef]
- Stokes, D.J. Principles and Practice of Variable Pressure/Environmental Scanning Electron Microscopy (VP-ESEM); John Wiley and Sons: West Sussex, UK, 2008. [Google Scholar]
- Cartwright, C.R. Identifying the woody resources of Diepkloof Rock Shelter (South Africa) using scanning electron microscopy of the MSA wood charcoal assemblages. J. Archaeol. Sci. 2013, 40, 3463–3474. [Google Scholar] [CrossRef]
- Balzano, A.; Merela, M.; Čufar, K. Scanning electron microscopy protocol for studying anatomy of highly degraded waterlogged archaeological wood. Forests 2022, 13, 161. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Archaeological Unknown Reed | Arundo donax | Phragmites australis |
|---|---|---|---|
| N° slices | 4 | 3 | 4 |
| TR (ms) | 450 | 800 | 1000 |
| TE (ms) | 4.7 | 2.8 | 3.0 |
| STK (μm) | 500 | 500 | 500 |
| FOV (cm) | 0.85 | 0.90 | 0.95 |
| MTX (mm2) | 512 | 512 | 512 |
| NS | 128 | 256 | 128 |
| AT (h) | 8 | 29 | 18 |
| R (μm2) | 17 × 17 | 17 × 17 | 18 × 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moricca, C.; Stagno, V.; Hamza, N.M.; Favero, G.; Sadori, L.; Capuani, S. The High Potential of Micro-Magnetic Resonance Imaging for the Identification of Archaeological Reeds: The Case Study of Tutankhamun. Heritage 2023, 6, 7170-7180. https://doi.org/10.3390/heritage6110375
Moricca C, Stagno V, Hamza NM, Favero G, Sadori L, Capuani S. The High Potential of Micro-Magnetic Resonance Imaging for the Identification of Archaeological Reeds: The Case Study of Tutankhamun. Heritage. 2023; 6(11):7170-7180. https://doi.org/10.3390/heritage6110375
Chicago/Turabian StyleMoricca, Claudia, Valeria Stagno, Nagmeldeen Morshed Hamza, Gabriele Favero, Laura Sadori, and Silvia Capuani. 2023. "The High Potential of Micro-Magnetic Resonance Imaging for the Identification of Archaeological Reeds: The Case Study of Tutankhamun" Heritage 6, no. 11: 7170-7180. https://doi.org/10.3390/heritage6110375
APA StyleMoricca, C., Stagno, V., Hamza, N. M., Favero, G., Sadori, L., & Capuani, S. (2023). The High Potential of Micro-Magnetic Resonance Imaging for the Identification of Archaeological Reeds: The Case Study of Tutankhamun. Heritage, 6(11), 7170-7180. https://doi.org/10.3390/heritage6110375