Environmental and Vegetative Controls on Soil CO2 Efflux in Three Semiarid Ecosystems

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil CO2 Efflux and Environmental Measurements
2.3. Ecosystem Photosynthesis
2.4. Data Analysis
2.5. Model Development
3. Results
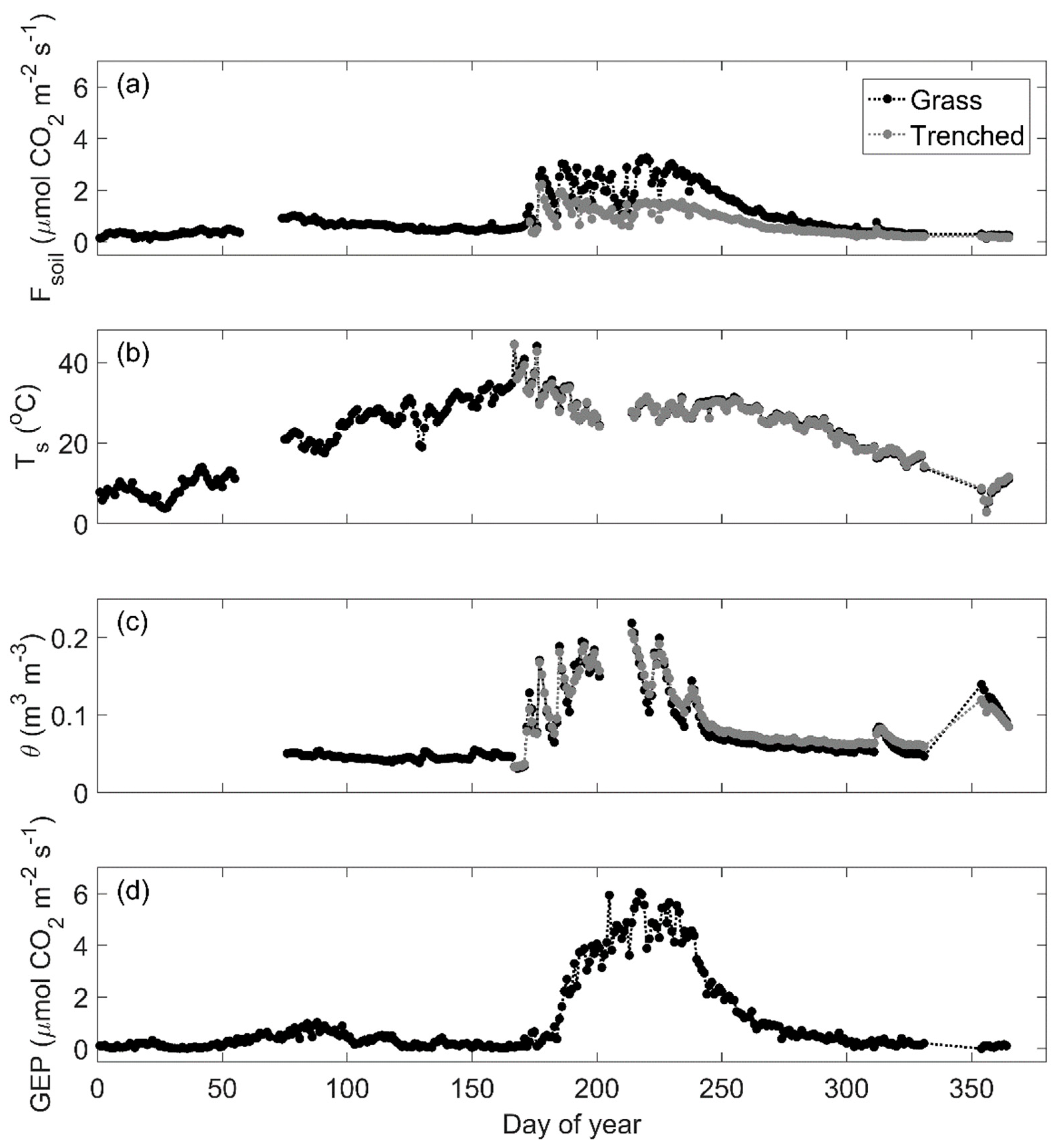
3.1. Seasonality of Soil CO2 Efflux
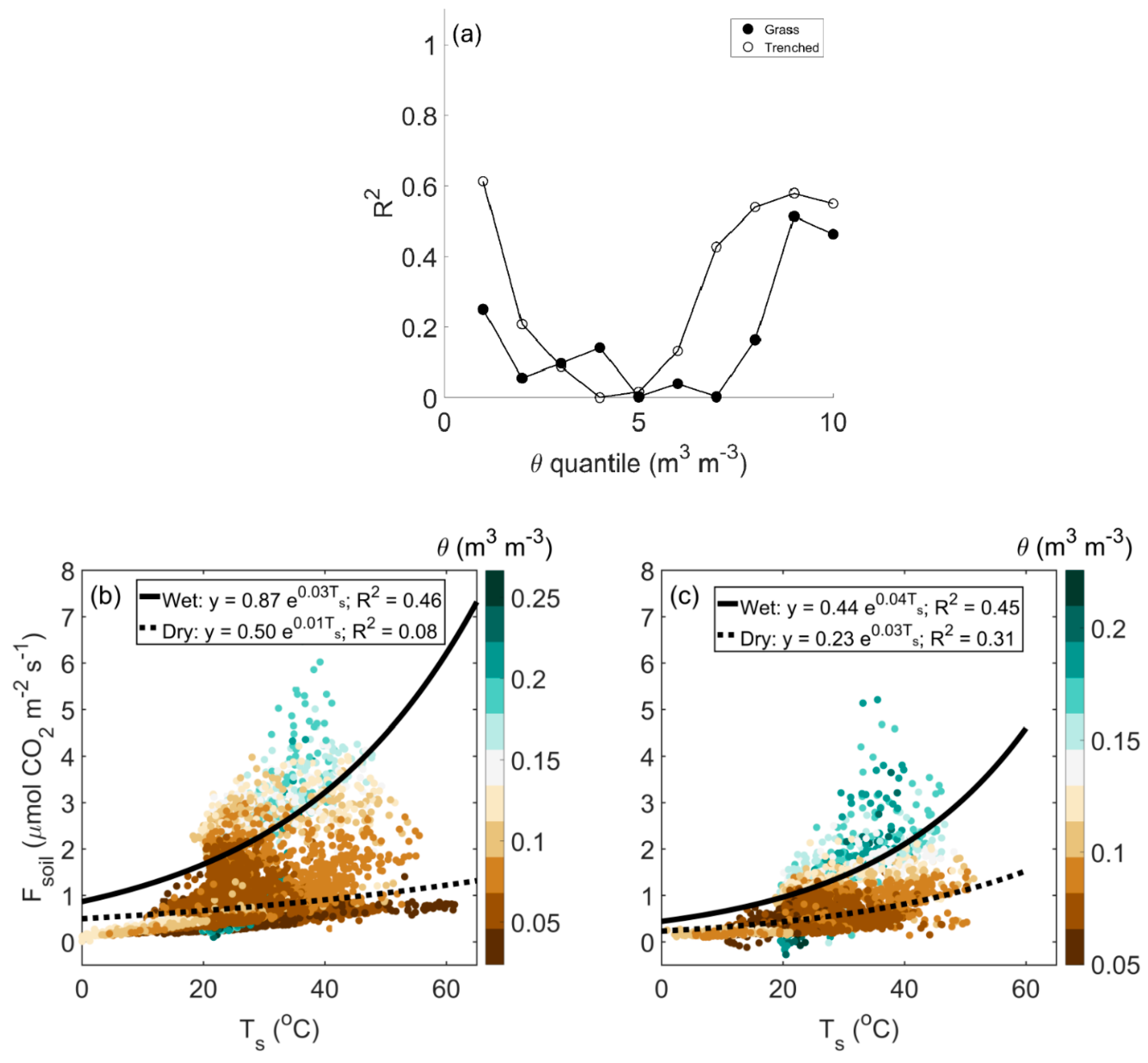
3.2. Environmental Controls on Soil CO2 Efflux
3.3. Physiological Controls on Soil CO2 Efflux
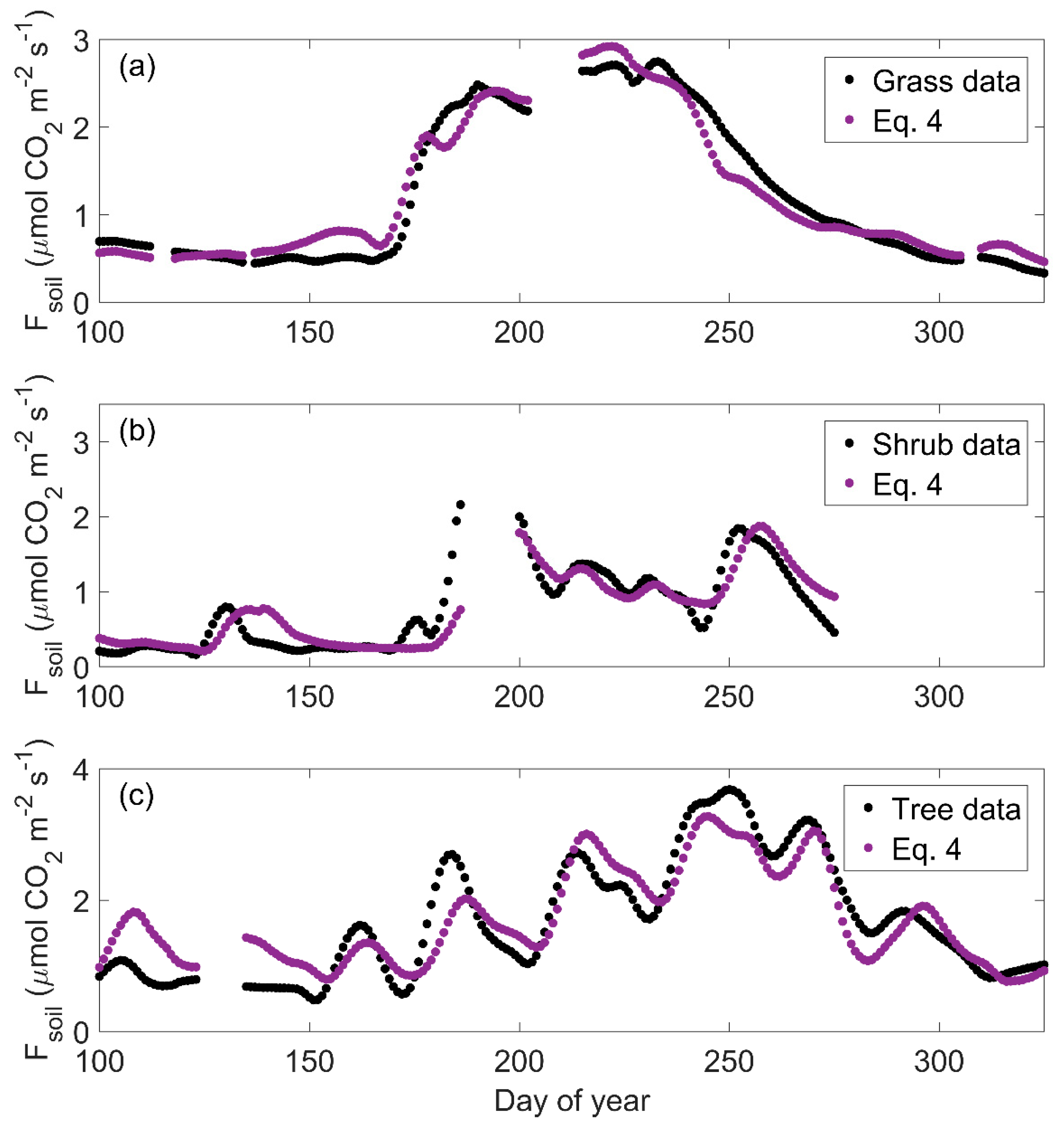
3.4. Model Performance
4. Discussion
4.1. Water Availability Limits Autotrophic and Heterotrophic Respiration
4.2. Ecosystem Photosynthesis Stimulates Soil CO2 Efflux
4.3. Moisture and Photosynthesis Terms Improve Modeled Carbon-Water Dynamics
4.4. Vegetation Activity and Structure Influence the Relative Importance of Soil CO2 Efflux Drivers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y.; Running, S.W.; et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Ahlström, A. The dominant role of semiarid ecosystems in the trend and variability of the land CO2 sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Valentini, R.; Matteucci, G.; Dolman, A.J.; Schulze, E.; Rebmann, C.; Moors, E.J.; Granier, A.; Gross, P.; Jensen, N.O. Respiration as the main determininant of carbon balance in European forests. Nature 2000, 404, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.L.; Hamerlynck, E.P.; Jenerette, G.D.; Moran, M.S.; Gafford, G.A.B. Carbon dioxide exchange in a semidesert grassland through drought-induced vegetation change. J. Geophys. Res. 2010, 115, 1–12. [Google Scholar] [CrossRef]
- Hamerlynck, E.P.; Scott, R.L.; Sánchez-Cañete, E.P.; Barron-Gafford, G.A. Nocturnal soil CO2 uptake and its relationship to subsurface soil and ecosystem carbon fluxes in a Chihuahuan Desert shrubland. J. Geophys. Res. Biogeosci. 2013, 118, 1593–1603. [Google Scholar] [CrossRef]
- Ryan, M.G.; Law, B.E. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 2005, 73, 3–27. [Google Scholar] [CrossRef]
- Sánchez-Cañete, E.P.; Scott, R.L.; van Haren, J.; Barron-Gafford, G.A. Improving the accuracy of the gradient method for determining soil carbon dioxide efflux: Accurate Long-Term Fsoil Based on the GM. J. Geophys. Res. Biogeosci. 2017, 122, 50–64. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the temperature dependence of soil respiration. Source Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent of confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Xu, M.; Qi, Y. Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.D. Spatial–temporal variation in soil respiration in an oak–grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 2005, 73, 183–207. [Google Scholar] [CrossRef]
- Sponseller, R.A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Glob. Chang. Biol. 2007, 13, 426–436. [Google Scholar] [CrossRef]
- Cable, J.M.; Barron-Gafford, G.A.; Ogle, K.; Pavao-Zuckerman, M.; Scott, R.L.; Williams, D.G.; Huxman, T.E. Shrub encroachment alters sensitivity of soil respiration to temperature and moisture. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Reichstein, M.; Rey, A.; Freibauer, A.; Tenhunen, J.; Valentini, R.; Banza, J.; Casals, P.; Cheng, Y.; Grünzweig, J.M.; Irvine, J.; et al. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Wang, B.; Zha, T.S.; Jia, X.; Gong, J.N.; Bourque, C.; Feng, W.; Tian, Y.; Wu, B.; Qing Zhang, Y.; Peltola, H. Soil water regulates the control of photosynthesis on diel hysteresis between soil respiration and temperature in a desert shrubland. Biogeosciences 2017, 14, 3899–3908. [Google Scholar] [CrossRef]
- Vargas, R.; Allen, M.F. Environmental controls and the influence of vegetation type, fine roots and rhizomorphs on diel and seasonal variation in soil respiration. New Phytol. 2008, 179, 460–471. [Google Scholar] [CrossRef]
- Xu, M.; Shang, H. Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef]
- Xu, L.; Baldocchi, D.D.; Tang, J. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Feng, S.; Fu, Q. Expansion of global drylands under a warming climate. Atmos. Chem. Phys. 2013, 13, 10081–10094. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Scott, R.L.; Huxman, T.E. Whole ecosystem metabolic pulses following precipitation events. Funct. Ecol. 2008, 22, 924–930. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Gavrichkova, O. Time lag between photosynthesis and carbon dioxide efflux from soil: A review of mechanisms and controls. Glob. Chang. Biol. 2010, 16, 3386–3406. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- Cable, J.M.; Ogle, K.; Lucas, R.W.; Huxman, T.E.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Ewers, B.E.; Pendall, E.; Welker, J.M.; et al. The temperature responses of soil respiration in deserts: A seven desert synthesis. Biogeochemistry 2011, 103, 71–90. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. A global database of soil respiration data. Biogeosciences 2010, 7, 1915–1926. [Google Scholar] [CrossRef]
- Yan, Z.; Bond-Lamberty, B.; Todd-Brown, K.E.; Bailey, V.L.; Li, S.; Liu, C.; Liu, C. A moisture function of soil heterotrophic respiration that incorporates microscale processes. Nat. Commun. 2018, 9, 2562. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Baldocchi, D.D.; Xu, L. Tree photosynthesis modulates soil respiration on a diurnal time scale. Glob. Chang. Biol. 2005, 11, 1298–1304. [Google Scholar] [CrossRef]
- Conant, R.T.; Klopatek, J.M.; Klopatek, C.C. Environmental factors controlling soil respiration in three semiarid ecosystems. Soil Sci. Soc. Am. J. 2000, 64, 383–390. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Novak, M.D.; Gaumont-Guay, D.; Nesic, Z. Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas-fir stand. Glob. Chang. Biol. 2008, 14, 1305–1318. [Google Scholar] [CrossRef]
- Luo, Y.; Ahlström, A.; Allison, S.D.; Batjes, N.H.; Brovkin, V.; Carvalhais, N.; Chappell, A.; Ciais, P.; Davidson, E.A.; Finzi, A.; et al. Toward more realistic projections of soil carbon dynamics by Earth system models. Glob. Biogeochem. Cycles 2016, 30, 40–56. [Google Scholar] [CrossRef]
- Zhang, Q.; Phillips, R.P.; Manzoni, S.; Scott, R.L.; Oishi, A.C.; Finzi, A.; Daly, E.; Vargas, R.; Novick, K.A. Changes in photosynthesis and soil moisture drive the seasonal soil respiration-temperature hysteresis relationship. Agric. For. Meteorol. 2018, 259, 184–195. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdina, N.; David, T.S.; David, J.S.; Ferreira, M.I.; Dohnal, M.; Tesař, M.; Gartner, K.; Leitgeb, E.; Nadezhdin, V.; Cermak, J.; et al. Trees never rest: The multiple facets of hydraulic redistribution. Ecohydrology 2010, 3, 431–444. [Google Scholar] [CrossRef]
- Barron-Gafford, G.A.; Scott, R.L.; Jenerette, G.D.; Huxman, T.E. The relative controls of temperature, soil moisture, and plant functional group on soil CO2 efflux at diel, seasonal, and annual scales. J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- Curiel Yuste, J.; Baldocchi, D.D.; Gershenson, A.; Goldstein, A.; Misson, L.; Wong, S. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Glob. Chang. Biol. 2007, 13, 2018–2035. [Google Scholar] [CrossRef]
- Cable, J.M.; Ogle, K.; Williams, D.G.; Weltzin, J.F.; Huxman, T.E. Soil texture drives responses of soil respiration to precipitation pulses in the sonoran desert: Implications for climate change. Ecosystems 2008, 11, 961–979. [Google Scholar] [CrossRef]
- Barron-Gafford, G.A.; Cable, J.M.; Bentley, L.P.; Scott, R.L.; Huxman, T.E.; Jenerette, G.D.; Ogle, K. Quantifying the timescales over which exogenous and endogenous conditions affect soil respiration. New Phytol. 2014, 202, 442–454. [Google Scholar] [CrossRef]
- Suseela, V.; Conant, R.T.; Wallenstein, M.D.; Dukes, J.S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Glob. Chang. Biol. 2012, 18, 336–348. [Google Scholar] [CrossRef]
- Sampson, D.A.; Janssens, I.A.; Curiel Yuste, J.; Ceulemans, R. Basal rates of soil respiration are correlated with photosynthesis in a mixed temperate forest. Glob. Chang. Biol. 2007, 13, 2008–2017. [Google Scholar] [CrossRef]
- Scott-Denton, L. Spatial and temporal controls of soil respiration rate in a high-elevation, subalpine forest. Soil Biol. Biochem. 2003, 35, 525–534. [Google Scholar] [CrossRef]
- Moore, D.J.P.; Trahan, N.A.; Wilkes, P.; Quaife, T.; Stephens, B.B.; Elder, K.; Desai, A.R.; Negron, J.; Monson, R.K. Persistent reduced ecosystem respiration after insect disturbance in high elevation forests. Ecol. Lett. 2013, 16, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Trahan, N.A.; Dynes, E.L.; Pugh, E.; Moore, D.J.P.; Monson, R.K. Changes in soil biogeochemistry following disturbance by girdling and mountain pine beetles in subalpine forests. Oecologia 2015, 177, 981–995. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.L.; Biederman, J.A.; Hamerlynck, E.P.; Barron-gafford, G.A. The carbon balance pivot point of southwestern U.S. semiarid ecosystems: Insights from the 21st century drought. J. Geophys. Res. Biogeosci. 2015, 120, 2612–2624. [Google Scholar] [CrossRef]
- Cox, J.R.; Frasier, G.W.; Renard, K.G. Biomass distribution at grassland and shrubland sites. Rangelands 1986, 8, 67–68. [Google Scholar]
- Scott, R.L.; Jenerette, G.D.; Potts, D.L.; Huxman, T.E. Effects of seasonal drought on net carbon dioxide exchange from a woody-plant-encroached semiarid grassland. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Barba, J.; Cueva, A.; Bahn, M.; Barron-Gafford, G.A.; Bond-Lamberty, B.; Hanson, P.J.; Jaimes, A.; Kulmala, L.; Pumpanen, J.; Scott, R.L.; et al. Comparing ecosystem and soil respiration: Review and key challenges of tower-based and soil measurements. Agric. For. Meteorol. 2018, 249, 434–443. [Google Scholar] [CrossRef]
- Andrade, J.M.; Estévez-Pérez, M.G. Statistical comparison of the slopes of two regression lines: A tutorial. Anal. Chim. Acta 2014, 838, 1–12. [Google Scholar] [CrossRef]
- Savage, K.; Davidson, E.A.; Richardson, A.D.; Hollinger, D.Y. Three scales of temporal resolution from automated soil respiration measurements. Agric. For. Meteorol. 2009, 149, 2012–2021. [Google Scholar] [CrossRef]
- Yan, L.; Chen, S.; Huang, J.; Lin, G. Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Glob. Chang. Biol. 2010, 16, 2345–2357. [Google Scholar] [CrossRef]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Selected Papers of Hirotugu Akaike; Springer: New York, NY, USA, 1998; pp. 199–213. [Google Scholar]
- Zeng, X.; Song, Y.; Zhang, W.; He, S. Spatio-temporal variation of soil respiration and its driving factors in semi-arid regions of North China. Chin. Geogr. Sci. 2018, 28, 12–24. [Google Scholar] [CrossRef]
- Davidson, E.A.; Richardson, A.D.; Savage, K.E.; Hollinger, D.Y. A distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce-dominated forest. Glob. Chang. Biol. 2006, 12, 230–239. [Google Scholar] [CrossRef]
- Moyano, F.E.; Manzoni, S.; Chenu, C. Responses of soil heterotrophic respiration to moisture availability: An exploration of processes and models. Soil Biol. Biochem. 2013, 59, 72–85. [Google Scholar] [CrossRef]
- Wang, B.; Zha, T.S.; Jia, X.; Wu, B.; Zhang, Y.Q.; Qin, S.G. Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. Biogeosciences 2014, 11, 259–268. [Google Scholar] [CrossRef]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Jenerette, G.D.; Barron-Gafford, G.A.; Guswa, A.J.; McDonnell, J.J.; Villegas, J.C. Organization of complexity in water limited ecohydrology. Ecohydrology 2012, 5, 184–199. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Xu, K.; Metzger, S.; Desai, A.R. Upscaling tower-observed turbulent exchange at fine spatio-temporal resolution using environmental response functions. Agric. For. Meteorol. 2017, 232, 10–22. [Google Scholar] [CrossRef]
- Savage, K.; Davidson, E.A.; Tang, J. Diel patterns of autotrophic and heterotrophic respiration among phenological stages. Glob. Chang. Biol. 2013, 19, 1151–1159. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garrten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Gomez-Casanovas, N.; Matamala, R.; Cook, D.R.; Gonzalez-Meler, M.A. Net ecosystem exchange modifies the relationship between the autotrophic and heterotrophic components of soil respiration with abiotic factors in prairie grasslands. Glob. Chang. Biol. 2012, 18, 2532–2545. [Google Scholar] [CrossRef]
- Vargas, R.; Sánchez-Cañete, P.E.; Serrano-Ortiz, P.; Curiel Yuste, J.; Domingo, F.; López-Ballesteros, A.; Oyonarte, C. Hot-moments of soil CO2 efflux in a water-limited grassland. Soil Syst. 2018, 2, 47. [Google Scholar] [CrossRef]
- Baldocchi, D.; Tang, J.; Xu, L. How switches and lags in biophysical regulators affect spatial-temporal variation of soil respiration in an oak-grass savanna. J. Geophys. Res. Biogeosci. 2006, 111. [Google Scholar] [CrossRef]
- Vargas, R.; Baldocchi, D.D.; Allen, M.F.; Bahn, M.; Black, T.A.; Collins, S.L.; Yuste, J.C.; Hirano, T.; Jassal, R.S.; Pumpanen, J.; et al. Looking deeper into the soil: Biophysical controls and seasonal lags of soil CO2 production and efflux. Ecol. Appl. 2010, 20, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Kurc, S.A.; Small, E.E. Soil moisture variations and ecosystem-scale fluxes of water and carbon in semiarid grassland and shrubland. Water Resour. Res. 2007, 43. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Scott, R.L.; Barron-Gafford, G.A.; Huxman, T.E. Gross primary production variability associated with meteorology, physiology, leaf area, and water supply in contrasting woodland and grassland semiarid riparian ecosystems. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Walnut Gulch Kendall Grassland (Grassland) | Lucky Hills Shrubland (Shrubland) | Santa Rita Mesquite Savanna (Savanna) | |
|---|---|---|---|
| Year | 2017 | 2012 | 2015 |
| Latitude, longitude (°) | 31.7378° N, 109.9428° W | 31.749° N, 110.052° W | 31.822° N, 110.867° W |
| Elevation (m) | 1530 | 1370 | 1120 |
| Mean air temp. (°C) | 15.6 | 17.6 | 19.0 |
| Mean annual precip. (1971–2010; mm) | 346 | 320 | 384 |
| Mean annual GEP (g C m−2 year−1) | 227 | 159 | 331 |
| Mean LAI (MODIS) | 0.30 | 0.25 | 0.37 |
| Woody cover (%) | 3 | 40 | 35 |
| Perennial grass/forb cover (%) | 37 | 3 | 15 |
| Soil type | Very gravelly, sandy to fine sandy, and clayey loams | Gravelly sandy loams | Deep loamy sands |
| Plots | Grass (n = 4) Trenched (n = 4) | Shrub (n = 4) Intercanopy (n = 4) | Under tree (n = 2) Intercanopy (n = 3) |
| Site | Plot | Model | Drivers | b | c | n | n | R2 | AIC | RMSE | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grassland | Grass | 1 | Ts | 0.44 | 0.04 | - | - | 244 | 0.12 | 568 | 0.80 |
| Grass | 2 | Ts, θ | 0.75 | 0.04 | 69.38 | - | 244 | 0.75 | 268 | 0.42 | |
| Grass | 3 | Ts, GEP | 1.10 | 0.04 | - | 0.23 | 244 | 0.77 | 252 | 0.41 | |
| Grass | 4 | Ts, θ, GEP | 0.99 | 0.04 | 56.54 | 0.84 | 244 | 0.86 | 142 | 0.33 | |
| Grassland | Trenched | 1 | Ts | 0.18 | 0.06 | - | - | 154 | 0.37 | 169 | 0.42 |
| Trenched | 2 | Ts, θ | 0.43 | 0.04 | 95.93 | - | 154 | 0.79 | 3 | 0.24 | |
| Trenched | 3 | Ts, GEP | 0.35 | 0.06 | - | 0.53 | 154 | 0.65 | 79 | 0.31 | |
| Trenched | 4 | Ts, θ, GEP | 0.47 | 0.04 | 89.34 | 3.80 | 154 | 0.80 | −3 | 0.24 | |
| Shrubland | Shrub | 1 | Ts | 0.76 | 0.00 | - | - | 164 | 0.00 | 293 | 0.59 |
| Shrub | 2 | Ts, θ | 2.61 | −0.02 | 103.63 | - | 164 | 0.66 | 119 | 0.35 | |
| Shrub | 3 | Ts, GEP | 2.11 | 0.00 | - | 0.24 | 164 | 0.50 | 182 | 0.42 | |
| Shrub | 4 | Ts, θ, GEP | 3.02 | −0.02 | 98.56 | 2.03 | 164 | 0.68 | 108 | 0.34 | |
| Shrub * | 3 * | Ts, GEP * | 1.76 | 0.01 | - | 0.18 | 163 | 0.63 | 130 | 0.36 | |
| Shrub * | 4 * | Ts, θ, GEP * | 3.18 | −0.02 | 96.89 | 1.25 | 163 | 0.74 | 76 | 0.31 | |
| Savanna | Tree | 1 | Ts | 0.85 | 0.03 | - | - | 255 | 0.05 | 711 | 0.98 |
| Tree | 2 | Ts, θ | 1.42 | 0.03 | 55.50 | - | 255 | 0.54 | 526 | 0.68 | |
| Tree | 3 | Ts, GEP | 2.82 | 0.01 | - | 0.18 | 255 | 0.41 | 591 | 0.77 | |
| Tree | 4 | Ts, θ, GEP | 3.04 | 0.01 | 47.92 | 0.61 | 255 | 0.64 | 466 | 0.60 | |
| Tree * | 3 * | Ts, GEP * | 3.07 | 0.01 | - | 0.11 | 253 | 0.52 | 532 | 0.69 | |
| Tree * | 4 * | Ts, θ, GEP * | 3.18 | 0.01 | 44.11 | 0.41 | 253 | 0.69 | 423 | 0.56 |
| Site | Lag (Days) | R2 | R2 |
|---|---|---|---|
| GEP-Fsoil | GEP-θ No lag | GEP-θ with lag | |
| Grassland | 0 | 0.53 | - |
| Shrubland | 1 | 0.34 | 0.47 |
| Savanna | 2 | 0.14 | 0.26 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roby, M.C.; Scott, R.L.; Barron-Gafford, G.A.; Hamerlynck, E.P.; Moore, D.J.P. Environmental and Vegetative Controls on Soil CO2 Efflux in Three Semiarid Ecosystems. Soil Syst. 2019, 3, 6. https://doi.org/10.3390/soilsystems3010006
Roby MC, Scott RL, Barron-Gafford GA, Hamerlynck EP, Moore DJP. Environmental and Vegetative Controls on Soil CO2 Efflux in Three Semiarid Ecosystems. Soil Systems. 2019; 3(1):6. https://doi.org/10.3390/soilsystems3010006
Chicago/Turabian StyleRoby, Matthew C., Russell L. Scott, Greg A. Barron-Gafford, Erik P. Hamerlynck, and David J. P. Moore. 2019. "Environmental and Vegetative Controls on Soil CO2 Efflux in Three Semiarid Ecosystems" Soil Systems 3, no. 1: 6. https://doi.org/10.3390/soilsystems3010006
APA StyleRoby, M. C., Scott, R. L., Barron-Gafford, G. A., Hamerlynck, E. P., & Moore, D. J. P. (2019). Environmental and Vegetative Controls on Soil CO2 Efflux in Three Semiarid Ecosystems. Soil Systems, 3(1), 6. https://doi.org/10.3390/soilsystems3010006