Abstract
Fire is a key disturbance affecting plant biodiversity patterns and evolution. Although a wide range of studies have shown important impacts of fire on vegetation, most have focused on taxonomic diversity, with less emphasis on other aspects of biodiversity, such as functional and phylogenetic diversity. Therefore, we assessed the recovery of biodiversity facets across different times since the last fire in semiarid shrublands in Northeast Iran. We quantified changes in plant biodiversity facets, including taxonomic, functional, and phylogenetic diversity, and the diversity of seven functional traits in five ecologically comparable sites that have experienced wildfire disturbances at short-term (1 and 4 year sites) and long-term (10 and 20 year sites) intervals, in com- parison to an unburnt site. Our results showed significant changes in all biodiversity facets related to the year since the last fire, with a significant increase in biodiversity and diversity of functional traits under long-term rather than short-term conditions, and in comparison to the unburned site. We conclude that wildfire influences the presence of plant species with distant functional and evolutionary relatedness and causes an increase in plant species and diversity of functional traits de- pending on time intervals. Therefore, wildfire can promote positive effects on the recovery of bio- diversity aspects and the evolution of vegetation in semiarid shrublands.
1. Introduction
Fire is one of the main disturbances influencing vegetation, plant biodiversity, and the functioning of terrestrial ecosystems [1,2,3,4,5,6]. For instance, fire can drive significant changes in species composition and species richness through a decrease in the competitive impacts of some woody plants [7]. Short-term fire occurrence may increase the abundance and frequency of annual and perennial grasses [8]. Some evidence suggests that, in addition to species composition changes, functional traits such as life form, plant height, specific leaf area, and leaf carbon and nitrogen content can change with changes in fire occurrence [9]. Such changes in species composition and functional traits following the fire may or may not result in significant variation in evolutionary diversity, depending on how functional traits are conserved across phylogenies [2,10].
Vegetation restoration is one of the most important objectives of management and sustainable development of arid and semiarid shrublands [11,12]. However, restoration is highly dependent on disturbances, and on the temporal scales of imposed disturbances [2,10,13,14]. In this regard, some evidence suggests positive correlations between vegetation restoration and time intervals, such as increased time intervals generally promoting increased plant biodiversity, coverage percentage, and abundance of plant species [15]. However, it remains unknown how vegetation, plant functional traits, and evolutionary diversity have been restored across different time scales since last fire. Plant density or coverage may be better enhanced under longer-term than shorter-term intervals since the last fire [2]. As a result, in order to better understand the temporal recovery of vegetation after fire occurrence, we must first understand the structural complexity of biodiversity facets and their direct effects on ecosystem functioning and vegetation evolution across different times since the last fire.
Plant biodiversity is an important feature that may clearly explain post-fire vegetation recovery [3,9]. Plant biodiversity contains different components, or facets including taxonomic, functional, and phylogenetic diversity. Much of the literature has focused on the effects of disturbance on taxonomic diversity, with less attention being paid to functional and phylogenetic diversity [16,17,18,19]. Plant functional diversity (FD) characterizes the variability of functional traits within a community, while phylogenetic diversity represents the variability of evolutionary clades in a community [20,21,22,23]. For instance, if plant species from different functional groups or distinct evolutionary clades occur in a community, increased functional and phylogenetic diversity may be observed. Such increases in functional or evolutionary variability can influence ecosystem processes and stability [24,25]. The responses of phylogenetic diversity to fire disturbances can differ from those of functional diversity if functional traits are not phylogenetically conserved [26] or if phylogeny represents important functional traits about which no information has been measured [27]. Therefore, evaluating all facets of biodiversity across different post-fire recovery intervals can help us fill important gaps in our knowledge of fire and its impacts on plant communities and ecosystems.
Although biodiversity facets provide essential information about the effects of fire on vegetation, some evaluation of ecosystem functioning during post-fire recovery suggests the importance of functional trait diversity [9]. In this regard, some studies have shown important effects of fire on traits related to plant adaptation, including specific leaf area, life form, seed mass, and leaf dry matter content [10,28]. However, it is not clear how such functional traits emerge under different lengths of time since the last fire and if these functional traits are phenotypically emerging or can be transferred to the next generation [10].
We aimed to investigate post-fire recovery of plant biodiversity facets (i.e., taxonomic, functional, and phylogenetic diversity) and functional trait diversity at the community level in semiarid shrublands of Iran, which may serve as an appropriate example (based on vegetation type) to understand how vegetation changes over time. We gathered data on phylogenetic history and seven functional traits for all plant taxa found in the semiarid shrublands we studied and addressed the following questions: (1) Does post-fire recovery of plant biodiversity differ across different time intervals since the last fire for all biodiversity facets? (2) Do biodiversity facets and functional traits vary across different time intervals? (3) Which functional traits change with changes in ecosystem functioning or with vegetation evolution?
2. Materials and Methods
2.1. Study Area
Our study sites were both unburnt and burned shrubland growing in the Fereizi region located in Chenaran, Northeast Iran (between 36°20′ and 36°55′ N and 58°30′ and 59°40′ E) (Figure 1a). The mean elevation is 2000 m, and the mean annual precipitation ranges from 280 mm to 340 mm with a cold semiarid climate based on Emberger’s method (Supplementary Materials Table S1). The rainy season begins in late October and last until early spring, with the main season lasting from February to April and an average temperature of 7.5 °C. [29]. Altitude, precipitation, and temperature values were obtained from WorldClim [30] at 30″ resolution (Supplementary Materials Table S1).
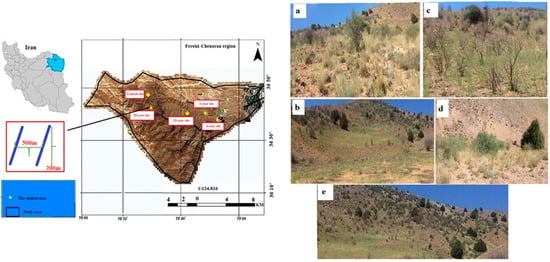
Figure 1.
The study region, with five study sites at different times since the last fire (i.e., 1-year, 4-year, 10-year, and 20-year sites, and a control site without fire occurrence) in the Fereizi-Chenaran shrublands in northeastern Iran, is represented in the above panel. Within each study site, two transects measuring 200 m in length and approximately 500 m apart were established, and 20 plots were placed on each one. Five study sites were surveyed in the Fereizi-Chenaran region, including (a) an unburnt site (i.e., a control site without fire occurrence), (b) 1-year sites (i.e., sites with the most recent fire occurring in the last year), (c) 4-year sites (i.e., sites with the most recent fire occurring 4 years prior), (d) 10-year sites and (e) 20-year sites (i.e., sites with the last fire occurring between 10 and 20 years prior) represented in the bottom panel.
The vegetation in the Fereizi region consists of grasses and shrubs in the form of semiarid shrubland landscapes. The dominant plant species consist of Juniperus polycarpos K. Koch (Cupressaceae), Lonicera nummulariifolia Jaub. & Spach. (Caprifoliaceae), Cotoneaster nummularius Lindl. (Rosaceae), Prunus microcarpa, Astragalus verus Olivier (Fabaceae), and some grasses, including Elymus hispidens (Opiz) Melderis (Poaceae), Phlomis cancellata Bunge (Lamiaceae), Poa bulbosa L. (Poaceae), and Taeniatherum caput-medusae (L.) Nevski (Poaceae).
2.2. Data Collection
2.2.1. Site Selection and Vegetation Sampling
We selected five sites based on documented wildfire histories in the area. These study sites occurred on north-facing slopes, and their geological formation was the same based on geological maps (Jurassic sandstone, shale, and conglomerate with marl and limestone) [31]. We selected one site for each of the following categories: (1) unburned site with no fire disturbance; (2) 1-year site (with the most recent fire occurring within the last year (in 2022)); (3) a 4-year site (most recent fire within 4 years before); (4) 10-year site (most recent fire within 10 years before); and (5) 20-year site (last fire having occurred within 20 years before) (Figure 1a). We interpret the noted times as representative of naturally occurring time intervals since the last fire, but we do not imply that these sites regularly have fires at the intervals noted.
To better recognize post-fire recovery of biodiversity facets, we categorized sites into three groups, including sites with short-term time intervals since the last fire [i.e., 1 and 4 year sites], sites with long-term time intervals since the last fire [i.e., 10 and 20 year sites], and the unburnt site. To investigate fire occurrence, we extracted wildfire histories for each site from aerial photos, and information on fire time recorded by environment department staff in the Fereizi region. Furthermore, we obtained comparable elevation and precipitation data from the WorldClim dataset for all of these sites (http://www.worldclim.org; (accessed on 1 February 2022) [32]) at 30″ resolution. Annual precipitation ranges from 271 mm to 327 mm (see details in Table S1 in Supplementary Materials).
In each of the five study sites, we established two transects of 200 m length, approximately 500 m apart, and 20 1 m × 1 m plots were placed 20 m apart on each transect. The number of plots and the appropriate area sampled for plots were assessed with species-area curves [33]. The vegetation was surveyed as a cover percentage for all present plants within each plot (Figure 1a). We identified 75 plant taxa in total at our study sites. We excluded 20 (out of 75) species with fewer than 5 individuals in the whole data set to avoid rare species effects [15] (Supplementary Materials Table S2).
We estimated fire intensity for each site based on classifications from Heydari et al. [34]. Under these classifications, fire intensity is calculated based on specific indicators, including: (i) obvious fire; (ii) herbaceous cover; (iii) the percentage of shrubs, herbs, and trees killed; and (iv) stem charring of tree height [34]. On the 1-year site, we found ground fire and scorched lower tree limbs. Herbaceous plants and some shrubs had been killed. Stem charring rose to about 70% of tree height. Between 25% and 30% of shrubs and herbs had been killed (Figure 1b). Finally, at the control site, there was no evidence of vegetation killed by fire, and no charring on tree stems was found. Based on such evidence, all burn sites indicated a comparable intermediate or moderate level of fire (Figure 1b). Therefore, we just focused on assessing the impacts of time since the last fire on plant biodiversity and the diversity of functional traits.
2.2.2. Plant Functional Traits
We assessed seven functional traits for plant species in our sites, including specific leaf area (SLA) (m 2.kg−1), leaf nitrogen (mg.g−1), carbon content (mg.g−1), plant height (cm), seed mass (mg), growth form, and life span (the last two traits); these traits were categorically ranked and included in the trait matrix for further analyses (see Table S2 for more details). These functional traits were introduced as key functional traits for plant species in burned ecosystems and are dependent upon biotic interactions and disturbance conditions [24,35]. For example, SLA generally reflects plant growth rate in fire-prone ecosystems, with increased SLA following fire because of the generation of open habitat that can increase the abundance of fast-growing and ruderal species relative to perennial species [36]. In addition, traits such as those related to plant regeneration (including seed mass, plant height, and plant growth) also represent plant response to wildfire [16]. Life span represents plant strategies and abilities, including competitive and facilitative abilities [37]. Leaf nitrogen/carbon content and life span can indicate post-fire resprouting in shrubs [38,39,40] and differentiate annual from perennial plant species in herbaceous communities [17].
We obtained information on these plant functional traits either from field measurements or from publicly available trait datasets (the BIEN package in R [41], TRY [42], and LEDA [43] and TR8 [44]). We considered plant height based on vegetative portions and measured it in the field directly, whereas other functional traits studied were obtained from publicly available datasets. When multiple measurements per species were available in these databases, we used the arithmetic mean of these observations to create a species mean trait value for each plot [45]. However, for some of our species, trait data were not available in existing databases. In those cases, we used the average of the trait values available for all the species at the genus level (see Supplementary Materials Table S2). Although not optimal, this is a solution to impute data for poorly studied taxa, somehow similar to what is commonly performed when evaluating phylogenetic relationships in less well-studied groups of species [22,46].
2.2.3. Phylogenetic Information
We constructed a phylogenetic tree for 55 plant species present in all studied sites using the “mega-tree” function in the V. PhyloMaker library in R [47] to generate a synthetic tree. This phylogenetic tree was generated by pruning and grafting taxa from an existing supertree [e.g., APG IV; 45], which is, as far as we know, the most extensive dated phylogeny for vascular plants, including 74, 533 species and all families of extant vascular plants [47]. We used the “scenario 3” approach (this scenario includes adding missing genus or species taxa to the phylogeny with known branch lengths [48] to add species to the phylogeny. This phylogenetic tree was then used to calculate phylogenetic diversity for our communities.
2.2.4. Measures of Taxonomic, Functional, and Phylogenetic Diversity
To assess post-fire recovery of biodiversity facets across different times since the last fire, we measured taxonomic, functional, and phylogenetic diversity for each 1 m2 plot in each site. To measure taxonomic diversity, we used the first two Hill numbers to estimate species richness (q = 0) and species diversity as the exponential of Shannon’s entropy (q = 1; referring to Shannon diversity) [49] for 1 m2 plots in each site. The calculation was based on the number of species, which is less affected by differences in total sampling effort than other methods [50].
To measure functional diversity, we assessed abundance-weighted mean pairwise distance (MPD) [51,52] as the most robust measure of functional relatedness at the species level for 1 m2 plots in each site. This index in the abundance-weighted case is equivalent to Rao’s Q and Hill numbers [52]. To better understand changes in functional traits depending on the time since the last fire, we also measured weighted means for each trait at the species level in each plot using a community-weighted mean index (CWM). CWM is quantified as the relative abundance of each plant species in terms of the trait values measured [53]. While wildfire disturbance is likely to result in intraspecific variability in functional traits, here we focus on species-level values of functional traits as a first-level test to detect if wildfire disturbance results in persistent changes to community functional traits [54]. We encourage future studies to collect trait data from individual plants across species to quantify how intraspecific trait variation changes with wildfire disturbance regimes. By using these multivariate functional indices at the plot level, we are able to compare communities’ characters before and after wildfire disturbance, as well as determine if community traits become more or less dispersed as a result of wildfire disturbance [54]. Before measuring functional diversity, we analyzed the correlation between pairs of functional traits via Pearson correlation analysis. We found no significant correlations between pairs of functional traits, so we considered all functional traits in further analyses and computed functional diversity via MPD.
We measured phylogenetic diversity as MPD, as used to measure phylogenetic relatedness at the species level, for each plot in each studied site. We calculated the standardized effect sizes of abundance-weighted MPD for functional (FSES.mpd) and phylogenetic (PSES.mpd) diversity to produce a phylogenetic and functional index of diversity that is independent of species richness. We used the independent-swap algorithm to draw a null distribution based on 999 randomizations and estimated standard effect sizes of MPD by comparing the observed community diversity to the null distribution of randomly assembled communities. Positive (SES values > 1.96) and negative (SES values < −1.96) values of SES.mpd indicate significant functional/phylogenetic over-dispersion and clustering patterns in the community, respectively. We calculated SES.mpd with the “ses.mpd” functions in the R package picante [53].
2.3. Statistical Analyses
We analyzed variation in taxonomic, functional, and phylogenetic diversity relative to the time since the last fire. We analyzed the response of taxonomic, functional, and phylogenetic diversity (using separate analyses for each) simultaneously with community-weighted means for functional traits within each plot (these variables were considered as response variables) relative to time since last fire (i.e., the fixed effect variable) via linear mixed effect models. We compared models with second-order Akaike information criteria (AIC) and R2adj values using the function ‘r.squaredGLMM’ in the package ‘MuMIn’ [55]. Finally, a one-way ANOVA and Tukey’s HSD multiple comparison test were applied to test whether time since the last fire resulted in significantly different plant biodiversity and diversity of functional traits [55].
The degree to which the studied traits showed a phylogenetic signal was assessed via the K-statistic to estimate the strength of phylogenetic conservatism for different traits using the ‘multiPhylosignal’ function in the R package picante [52]. The K-statistic was compared to randomized K values, with significant p-values indicating non-random correspondence between traits and phylogeny. Significant values of K greater than 1.0 indicate that traits are more conserved than expected by a Brownian motion evolutionary model [56]. Values lower than 1.0 indicate that traits are less conserved along the phylogeny than expected by this model.
3. Results
3.1. Plant Biodiversity Changes across Time after Wildfire
We found that time since last fire was a significant and effective factor on variation in plant biodiversity facets at the studied sites (Figure 2; q0, q1, FSES.mpd and PSES.mpd panels), with higher values of taxonomic, functional, and phylogenetic diversity in sites with longer-term time intervals since last fire (see results of Tukey’s HSD multiple comparison test and one-way ANOVA in q0, q1, FSES.mpd, and PSES.mpd panels of Figure 2). However, the results of linear mixed effect models and one-way ANOVA indicated greater differences in functional and phylogenetic diversity than taxonomic diversity between the studied sites (see mR2adj and p-values in Figure 2). Such findings suggest significant overdispersion patterns for functional and phylogenetic diversity over longer time intervals (Figure 2; see long-term site in FSES.mpd and PSES.mpd panels) and significant clustering patterns (i.e., less disturbed patterns) over short intervals and in unburned areas (Figure 2; see short-term and unburned site in FSES.mpd and PSES.mpd panels).
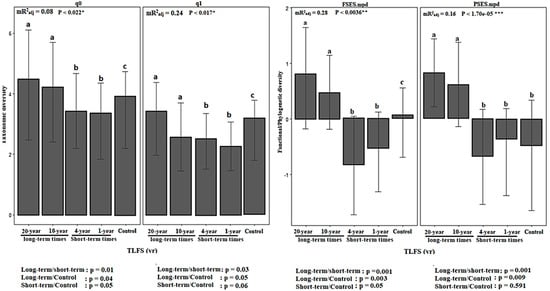
Figure 2.
Vertical bars showing taxonomic (i.e., species richness (q0) and Shannon’s diversity (q1)), functional (FSES.mpd), and phylogenetic (PSES.mpd) diversity at the studied sites across different times since last fire. Variation in taxonomic, functional, and phylogenetic indices across time since the last fire was assessed with linear mixed effect models. The marginal adjusted R2 explained by the fixed factors in our statistical models (mR2adj. are shown in each case) and the significance of time since last fire on variation in biodiversity facets were tested via ANOVA analysis. In addition, the results of the one-way ANOVA and Tukey’s HSD multiple comparisons (p < 0.05) represent differences between different time scales and within them, respectively. Error bars represent the uncertainty or variations in plant communities based on functional and evolutionary over-dispersion (higher values than zero) and under-dispersion (lower values than zero) patterns. Significant results of one-way ANOVA on biodiversity indices are shown in each panel: *, p < 0.05; **, p < 0.01; ***, p < 0.001. Letters (a, b, c) show the results of the Tukey tests for variation within each the time since last fire when significant.
3.2. Diversity of Functional Traits across Time Scales
The time since the last fire had a significant impact on the diversity of functional traits (Figure 3; see mR2adj and p-values for functional traits). In this regard, significant differences were observed between long-term sites and short-term and unburnt sites in terms of community-weighted mean values of some functional traits, including plant height, leaf carbon and nitrogen contents, and seed mass (see results of a one-way ANOVA for CWM-plantheight, CWM-C, CWM-N, and CWM-seedmass in Figure 3). Such differences were consistent with significant increases in the diversity of these functional traits in long-term sites (see Figure 3; CWM-plantheight, CWM-C, CWM-N, and CWM-seedmass panels). In contrast, higher community-weighted mean values of specific leaf area were found in sites with short-term intervals (Figure 3; CWM-SLA).
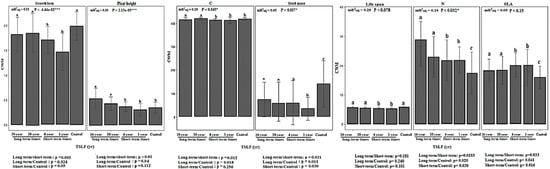
Figure 3.
Vertical bars showing the diversity of functional traits at the community level (i.e., CWM of traits), including growth form, plant height, leaf carbon (C) and nitrogen (N) content, seed mass, life span, and SLA across different times since the last fire. Variation in functional trait diversity across time since the last fire was assessed with linear mixed effect models. We also show the marginal adjusted R2 explained by the fixed factors in our statistical models (mR2adj). In addition, results of one-way ANOVA and Tukey’s HSD multiple comparisons (p < 0.05) represent differences between different time scales and within them, respectively. Error bars represent the uncertainty or presence of plant taxa with higher values of functional traits than mean values. Significant results of one-way ANOVA on biodiversity indices are shown in each panel: *, p < 0.05; ***, p < 0.001. Letters (a, b, c) show the results of the Tukey tests for variation within each the time since last fire when significant.
In addition, our results indicated a significant increase in the diversity of growth form and life span traits at long-term sites when compared to short-term and control sites (Figure 3; CWM-lifeform, CWM-lifespan). The results of statistical analyses showed significant effects of time since the last fire on post-recovery of functional traits including growth form, seed mass, C, N, and plant height (see mR2adj and p-values and results of Tukey’s HSD and one-way ANOVA in Figure 3). However, such effects were more consistent and positive for seed mass (CWM-seedmass; mR2adj = 0.45; p < 0.037), growth form (CWM-growth form; mR2adj = 0.31; p < 4.44 × 10−2), and plant height (CWM-plantheight; mR2adj = 0.30; p < 2.15 × 10−5) than for other traits (see Figure 3).
We found significant phylogenetic conservatism for seed mass, plant height, specific leaf area, leaf carbon content, and growth form in plant taxa present in studied semiarid shrublands across time since the last fire (Table 1). Generally, phylogenetic conservatism was much higher for growth form, leaf carbon content, and seed mass than plant height and specific leaf area for plant taxa in the studied sites (Table 1). In contrast, other functional traits, including life span, carbon/nitrogen ratio, and leaf nitrogen content, were not phylogenetically conserved in plant taxa studied across time since the last fire (Table 1). Overall, it seems that there is a strong and positive correlation between functional and phylogenetic diversity, with five out of seven traits showing significant conservatism.

Table 1.
Phylogenetic signal for the functional traits in the studied sites at different time scales since the last fire (Blomberg’s K-statistic for individual trait levels). Significant results are shown in bold (* p < 0.05; ** p < 0.01; ns: these traits do not show phylogenetic conservatism).
4. Discussion
4.1. Post-Fire Recovery of Plant Biodiversity Facets across Time since Last Fire
We found significant variation in plant biodiversity and functional traits across time since the last fire in the semi-arid shrublands. In this regard, we found significant increases in taxonomic, functional, and phylogenetic diversity across sites from which fire was absent for the longest time (i.e., 10 and 20 years). In contrast, functional/phylogenetic clustering patterns simultaneously associated with decreased taxonomic diversity were observed in sites with the shortest time since the last fire (i.e., 1 and 4 years). Although time since the last fire has promoted significant changes in plant biodiversity facets and functional traits, we found more consistent and positive effects of the temporal scale on functional and taxonomic diversity (q = 1 index) than phylogenetic diversity.
In agreement with our results, some previous evidence suggested that time since the last fire is a key factor in the post-fire recovery of vegetation [9,15]. However, the most effective recovery of vegetation was observed under long-term conditions, which directly confirms the intermediate disturbance hypothesis [10]. According to this hypothesis, although an increase in species diversity is strongly correlated with an increase in time intervals, intermediate and long time intervals can provide better conditions for recruitment of plant species through a relatively decreased fire disturbance [57]. Nonetheless, some previous studies have shown no effects of time scale on plant biodiversity recovery after fire disturbance [10]. It is possible that these neutral patterns in vegetation recovery observed in some studies were indirectly influenced by extreme environmental factors, including soil and climatic factors [16].
In our study region, significant variation in taxonomic diversity indices in sites where fire has been absent for the longest time suggests autosuccesion. In fire-prone ecosystems, almost all plant species in the post-fire community recorded before the fire were evident within 12 months of the fire [10,57] conforming to the ‘autosuccession’ [10,57] or ‘initial floristic composition’ [58] models of community succession. However, under long-term conditions, we found more species richness for fire-disturbed sites than in non-fire-disturbed sites, which supports the idea that fire provides suitable conditions for recruitment or survival of species that cannot survive without fire. The heat resulting from fire generally stimulates soil and canopy seed banks and can add some new plant species to the ecosystem [33]. Based on such phenomena, some functional or evolutionary changes may occur in burned ecosystems [10,17].
An increase in functional and phylogenetic diversity (i.e., significant functional and phylogenetic overdispesion patterns) with increasing time since the last fire may be related to a decrease in grass and herb dominance. It is clear that frequent wildfires promote the cover dominance of ruderal plants, grasses, and broadleaf herbs with strong competitive abilities to occupy disturbed and degraded ecosystems [16,59]. Grasses and herbs are generally favored under stressful conditions based on their competitive nature, shallow roots, and dense and small canopy areas [60]. However, when fire intervals decrease, woody species, including shrubs, trees, and cushion plants, may be replaced by such opportunistic plants [61,62]. Thus, since the last fire, more plant species with diverse growth forms may have coexisted under long-term conditions [63].
4.2. Post-Fire Recovery of Plant Biodiversity Consistent with Changes in Trait, Evolution, and Ecosystem Stability
The increase in functional and phylogenetic diversity in semi-arid shrubland sites where fire has been absent for decades promotes the presence of plant species with distant functional or evolutionary relatedness through a decrease in negative biotic interactions such as competition and an increase in positive interactions such as facilitation [25,64]. In fore-prone sites 10–20 years prior, we found plant species with different functional identities, including grasses, broadleaf herbs, and shrubs belonging to major functional groups or phylogenetic clades. In contrast, sites that burned 1–4 years prior were mostly covered by grasses and herbaceous plants. Based on such evidence, sites burned relatively recently (i.e., in the first 1–4 years post-fire) only provided suitable environments to recruit competitive and tolerant plant species with similar functional (e.g., low seed mass and high specific leaf area) or phylogenetic identities [64], thus causing a decrease in all facets of plant biodiversity. However, 10–20 years after the fire, positive interactions between distant plant taxa enhanced the presence of a wide range of plant species with different functional traits to recover all facets of plant biodiversity [63,65].
Our results on trait diversity indicated a significant increase in diversity of some functional traits, including seed mass, growth form, life span, leaf carbon, and nitrogen content, since the last fire. Growth form and life span diversity may explain the presence of different functional groups (i.e., grasses, herbs, and shrubs) to enhance all facets of plant biodiversity over long periods of time [66]. In contrast, under short-term conditions, more values of specific leaf area traits were found than under long-term conditions, indicating the dominance of grasses and broadleaf herbs at such sites [67]. In this regard, plant species in fire-disturbed sites showed phylogenetic conservatism in their regeneration niches, which was related to some functional traits such as seed mass, growth form, leaf carbon and nitrogen content, and specific leaf area, demonstrating the important effects of fire on vegetation in changing trait and evolutionary diversity [67]. Positive correlations between evolutionary change in vegetation and significant phylogenetic conservatism for five out of seven traits could explain post-fire vegetation recovery, which is consistent with the secondary succession hypothesis and autosuccession [65]. Therefore, we conclude that depending on the length of time intervals since the last fire, post-fire recovery of plant biodiversity facets occurred, and fire generally promoted significant changes in evolutionary lineages [28] and functional trait diversity to enhance ecosystem stability over long periods of time. However, our results regarding trait diversity and recovery of functional diversity across time since the last fire should be considered with caution. We propose this to be interpreted as a working hypothesis to be evaluated with a database that is enriched by plant traits of Mediterranean and semi-arid species and in light of recurrent disturbances and ongoing climate change.
4.3. Management Implications
Managing fire in a cost-effective manner may help conserve vegetation in mountainous semiarid shrublands by decreasing the loss of plant biodiversity and community composition [15]. Although we found that a long time interval may lead to increases in biodiversity facets and potentially even change the evolution of vegetation in our study area, some biological wildfire prevention tools to decrease fire intensity or fire occurrence may be effective to prevent the loss of plant biodiversity at short intervals [68]. For example, some studies have shown that wildlife grazing may decrease fuel load and fire intensity in the context of increasing drought under climate change [69]. Therefore, low to intermediate grazing intensity in sites with low herbaceous and grass species can ameliorate the negative impacts of fire intensity.
In addition, our results are consistent with the goals and strategies of nature conservation organizations to realize the United Nations’ recently adopted sustainable development goals (SDGs) [70]. We suggest that land managers of semiarid shrublands particularly focus on goals 13 and 15, which call to “Take urgent action to combat climate change and its impacts;” and “Focus on protecting, restoring and promoting sustainable use of terrestrial ecosystems and halting and reversing land degradation and halting biodiversity loss”, respectively (sustainabledevelopment.un.org) [71]. According to our results, this requires a reduction in fire occurrence in order to create short-term sites with lower fire risk [72] and to better maintain plant biodiversity and vegetation in post-fire recovery process over short time periods since last fire.
5. Conclusions
Taxonomic, functional, and phylogenetic diversity changed significantly depending on the time since the last fire. However, stronger differences in functional and phylogenetic diversity were observed than taxonomic differences between the studied sites, with overdispersion patterns found across long decades and clustering patterns in short times. In addition, fire significantly influenced trait diversity at the community level and trait evolution across phylogeny, with positive effects on some traits including seed mass, growth forms, life span, and leaf carbon and nitrogen content. It appears that the presence of distantly functionally or evolutionarily related taxa facilitated the post-fire recovery of plant biodiversity facets, with an increase in all diversity facets after decades. Such plant taxa can generally coexist when facilitative interactions between distantly related taxa are enhanced. In contrast, competitive interactions in communities are reduced via the presence of woody plant taxa. Based on these findings, we conclude that post-fire plant biodiversity recovery in the studied semiarid shrublands occurred in general, consistent with significant changes on functional traits and vegetation evolution following fire disturbance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fire6030103/s1, Table S1: Characteristics of the studied sites with more of five points: Longitude, Latitude, Altitude, Precipitation and Time since last fire; Table S2: A list of plant species presents in study sites and their functional trait values. Mean trait values were obtained in species level except for plant species marked by * (for these plant species, mean trait values were obtained in genus level).
Author Contributions
M.F. designed the study; M.B. and M.F. provided the data; data analyses and preparation of figures and tables were performed by M.B.; the paper was written by M.B., M.A. and R.P.S.; and all authors substantially contributed to the subsequent drafts. All authors have read and agreed to the published version of the manuscript.
Funding
This research is supported by the Iran National Science Foundation: INSF (Grant No. 4000679).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used to generate the results in this paper are available in Tables S1 and S2 in the Supplementary Materials File.
Acknowledgments
The authors wish to thank the Iran National Science Foundation. This research was conducted as part of the postdoctoral program of M.B. under the supervision of M.F. and was financially supported by the Iran National Science Foundation: INSF (Grant No. 4000679).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Badano, E.I.; Samour-Nieva, O.R.; Flores, J.; Flores-Flores, J.L.; Flores-Cano, J.A.; Rodas-Ortíz, J.P. Facilitation by nurse plants contributes to vegetation recovery in human-disturbed desert ecosystems. J. Plant Ecol. 2016, 9, 485–497. [Google Scholar] [CrossRef]
- Ren, H.; Yang, L.; Liu, N. Nurse plant theory and its application in ecological restoration in lower subtropics in China. Prog. Nat. Sci. 2008, 18, 137–142. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J.; Pither, J.; Thompson, J.; Zimmerman, J.K. The problem and promise of scale dependency in community phylogenetics. Ecology 2006, 87, 2418–2424. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Davies, T.J. Phylogenies in Ecology: A Guide to Concepts and Methods; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef]
- Yang, J.; Swenson, N.J.; Zhang, G.; Ci, X.; Cao, M.; Sha, L.; Lin, L. Local-scale partitioning of functional and phylogenetic beta diversity in a tropical tree assemblage. Sci. Rep. 2015, 5, 12731. [Google Scholar] [CrossRef]
- Piston, N.; Schob, C.; Armas, C.; Prieto, I. Contribution of co-occurring shrub species to community richness and phylogenetic diversity along an environmental gradient. Perspect. Plant Ecol. Evol. Syst. 2016, 19, 30–39. [Google Scholar] [CrossRef]
- Keesstra, S.D.; Bouma, J.; Wallinga, J.; Tittonell, P.; Smith, P.; Cerdà, A.; Quinton, J.N.; Pachepsky, Y.; van der Putten, W.H.; Bardgett, R.D. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2016, 2, 111–128. [Google Scholar] [CrossRef]
- Bowd, E.J.; Lindenmayer, D.B.; Banks, S.C.; Blair, D.P. Logging and fire regimes alter plant communities. Ecol. Applicat. 2018, 28, 826–841. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Perry, G.L.; Higgins, S.I.; Johnson, C.N.; Fuhlendorf, S.D.; Murphy, B.P. Pyrodiversity is the coupling of biodiversity and fire regimes in food webs. Philos. Trans. R. Soc. B. 2016, 371, 20150169. [Google Scholar] [CrossRef] [PubMed]
- Hurteau, M.D.; Bradford, J.B.; Fulé, P.Z.; Taylor, A.H.; Martin, K.L. Climate change, fire management, and ecological services in the southwestern US. For. Ecol. Manag. [CrossRef]
- Pausas, J.G.; Ribeiro, E. The global fire-productivity relationship. Glob. Ecol. Biogeog. 2013, 22, 728–736. [Google Scholar] [CrossRef]
- Vallego, V.R.; Arianoutsou, M.; Moreira, F. Fire Ecology and Post-Fire Restoration Approaches in Southern European Forest Types. Manag. For. Ecosyst. 2012, 24, 93–119. [Google Scholar] [CrossRef]
- Bahalkeh, K.; Abedi, M.; Tilaki, D.; Michalet, R. Fire slightly decreases on the short-term the competitive effects of a thorny cushion shrub in a semi-arid mountain steppe. Appl. Veg. Sci. 2021, 24, e12575. [Google Scholar] [CrossRef]
- Abedi, M.; Omidipour, R.; Hosseini, S.V.; Bahalkeh, K.; Gross, N. Fire disturbance effects on plant taxonomic and functional β-diversity mediated by topographic exposure. Ecol. Evol. 2022, 12, e8552. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2016, 16, 406–411. [Google Scholar] [CrossRef]
- Hernandez-Serrano, A.; Verdu, M.; Gonzalez-Martínez, S.C.; Pausas, J.G. Fire structures pine serotiny at different scales. Am. J. Bot. 2013, 100, 2349–2356. [Google Scholar] [CrossRef]
- Jankju, M.; Delavari, A.; Ganjali, A. Interseeding Bromus kopetdaghensis, in shrublands. Iran. J. Range Sci. 2008, 2, 314–328. [Google Scholar]
- Huston, M.A. Disturbance, productivity, and species diversity: Empiricism vs. logic in ecological theory. Ecology 2014, 95, 2382–2396. [Google Scholar] [CrossRef]
- Omidipour, R.; Tahmasebi, P.; Faizabadi, M.F.; Faramarzi, M.; Ebrahimi, A. Does β diversity predict ecosystem productivity better than species diversity? Ecol. Indic. 2021, 122, 107212. [Google Scholar] [CrossRef]
- Bashirzadeh, M.; Soliveres, S.; Farzam, M.; Ejtehadi, H. Plant–plant interactions determine taxonomic, functional and phylogenetic diversity in severe ecosystems. Glob. Ecol. Biogeog. 2022, 31, 649–662. [Google Scholar] [CrossRef]
- Pashirzad, M.; Ejtehadi, H.; Vaezi, J.; Sheferson, R.P. Spatial scale dependent phylogenetic signal in species distributions along geo graphic and elevation gradients in a mountainous rangeland. Ecol. Evol. 2018, 8, 10364–10373. [Google Scholar] [CrossRef]
- Soliveres, S.; Maestre, F.T.; Bowker, M.A.; Torices, R.; Quero, J.L.; Garcia-Gomez, M.; Noumi, Z. Functional traits determine plant co-occurrence more than environment or evolutionary relatedness in global drylands. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Soliveres, S.; Torices, R.; Maestre, F.T. Environmental conditions and biotic interactions acting together promote phylogenetic randomness in semi-arid plant communities: New methods help to avoid misleading conclusions. J. Veg. Sci. 2012, 23, 822–836. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Hubbell, S.P. The phylogenetic structure of a neotropical forest tree community. Ecology 2006, 87, S86–S99. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, Y.J.; Zhang, J.; Wang, X. Latitudinal gradients in phylogenetic relatedness of angiosperm trees in North America. Glob. Ecol. Biogeog. 2013, 22, 1183–1191. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Verdu, M. Facilitation can increase the phylogenetic diversity of plant communities. Ecol. Lett. 2007, 10, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Vega-Alvarez, J.; Garcia-Rodriguez, J.; Cayuela, L. Facilitation beyond specie richness. J. Ecol. 2019, 107, 722–734. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Barbosa, N.P.U.; Alberton, B.; Barbieri, A.; Dirzo, R.; Goulart, F.; Guerra, T.J.; Morellato, L.P.C.; Solar, R.R.C. The deadly route to collapse and the uncertain fate of Brazilian rupestrian grasslands. Biodiv. Conser. 2018, 27, 2587–2603. [Google Scholar] [CrossRef]
- Memariani, F.; Joharchi, M.R.; Ejtehadi, H.; Emadzadeh, K.H. A cintribution of the flora and vegetation of Binalood mountain range. NE Iran: Floristic and Chrological studies in Fereizi. Ferdowsi Univ. Int. J. Biol. Sci. 2009, 1, 1–17. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Inter. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Bashirzadeh, M.; Shefferson, R.P.; Farzam, M. Plant–plant interactions determine natural restoration of plant biodiversity over time, in a degraded mined land. Ecol. Evol. 2022, 12, e8878. [Google Scholar] [CrossRef] [PubMed]
- Heydari, M.; Omidipour, R.; Abedi, M.; Baskin, C. Effects of fire disturbance on alpha and beta diversity and on beta diversity components of soil seed banks and aboveground vegetation. Plant Ecol. Evol. 2017, 150, 247–256. [Google Scholar] [CrossRef]
- Lienin, P.; Kleyer, M. Plant trait responses to the environment and effects on ecosystem properties. Basic Appl. Ecol. 2012, 13, 301–311. [Google Scholar] [CrossRef]
- Lavorel, S.; Díaz, S.; Cornelissen, J.H.C.; Garnier, E.; Harrison, S.P.; McIntyre, S.; Pausas, J.G.; Pérez-Harguindeguy, N.; Roumet, C.; Urcelay, C. Plant functional types: Are we getting any closer to the Holy Grail? In Terrestrial Ecosystems in a Changing World; Canadell, J., Pitelka, L.F., Pataki, D., Eds.; Springer: Berlin, Germany, 2007; pp. 149–164. [Google Scholar]
- Davies, K.W.; Svejcar, T.J.; Bates, J.D. Interaction of historical and nonhistorical disturbances maintains native plant communities. Ecol. Applic. 2009, 19, 1536–1545. [Google Scholar] [CrossRef]
- Rahmanian, S.; Hejda, M.; Ejtehadi, H.; Farzam, M.; Memariani, F.; Pyšek, P. Effects of livestock grazing on soil, plant functional diversity, and ecological traits vary between regions with different climates in northeastern Iran. Ecol. Evol. 2019, 9, 8225–8237. [Google Scholar] [CrossRef]
- Anacker, B.; Rajakaruna, N.; Ackerly, D.; Harrison, S.; Keeley, J.; Vasey, M. Ecological strategies in California chaparral: Interacting effects of soils, climate, and fire on specific leaf area. Plant Ecol. Div. 2011, 4, 179–188. [Google Scholar] [CrossRef]
- Sakschewski, B.; von Bloh, W.; Boit, A.; Rammig, A.; Kattge, J.; Poorter, L.; Peñuelas, J.; Thonicke, K. Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model. Glob. Change Biol. 2015, 21, 2711–2725. [Google Scholar] [CrossRef]
- Maitner, B.S.; Boyle, B.; Casler, N.; Condit, R.; Enquist, B.J. The BIEN R package: A tool to access the Botanical Information and Ecology Network (BIEN) database. Method Ecol. Evol. 2018, 9, 373–379. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Change Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Sonnenschein, P.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; Klotz, S.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Bocci, G. TR8: An R package for easily retrieving plant species traits. Methods Ecol. Evol. 2015, 6, 347–450. [Google Scholar] [CrossRef]
- Lamanna, C.; Blonder, B.; Violle, C.; Kraft, N.J.B.; Sandel, B.; Imova, I.; Donoghue, J.C.; Svenning, J.C.; McGill, B.J.; Boyle, B.; et al. Functional trait space and the latitudinal diversity gradient. Proc. Natl. Acad. Sci. USA 2014, 111, 13745–13750. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, L.A.; Brooker, R.W.; Butterfield, B.J.; Cook, B.J.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Pugnaire, F.I.; Schöb, C.; Xiao, S.; et al. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 2013, 17, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Cayuela, L.; Granzow-de la Cerda, Í.; Albuquerque, F.S.; Golicher, D.J. taxonstand: An r package for species names standardization in vegetation databases. Methods Ecol. Evol. 2012, 3, 1078–1083. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Davies, T.J.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 2017, 92, 698–715. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Ann. Rev. Ecol. Syst. 2012, 33, 475–505. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Burkle, L.A.; Simanonok, M.P.; Durney, J.S.; Myers, J.A.; Belote, R.T. Wildfires Influence Abundance, Diversity, and Intraspecific and Interspecific Trait Variation of Native Bees and Flowering Plants Across Burned and Unburned Landscapes. Front. Ecol. Evol. 2019, 7, 252. [Google Scholar] [CrossRef]
- Barton, K. Package ‘MuMIn’. Model Selection and Model Averaging Based on Information Criteria. R Package Version 1.15.11. 2013. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 2 January 2023).
- Blomberg, S.P.; Garland, T.J.r.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Herath, D.N.; Lamont, B.B.; Enright, N.J.; Miller, B.P. Impact of fire on plant-species persistence in post-mine restored and natural shrubland communities in southwestern Australia. Biol. Conserv. 2009, 142, 2175–2180. [Google Scholar] [CrossRef]
- Hanes, T.L. Succession after fire in the chaparral of southern California. Ecol. Monogr. 1971, 41, 27–52. [Google Scholar] [CrossRef]
- Enright, N.J.; Mosner, E.; Miller, B.P.; Johnson, N.; Lamont, B.B. Soil versus canopy seed storage and plant species coexistence in species-rich shrublands of southwestern Australia. Ecology 2007, 88, 2292–2304. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Miesel, J.; Rogers, B.M.; Schweitzer, J.; Shuman, J.K.; Tepley, A.J.; Varner, J.M.; Veblen, T.T.; Adalsteinsson, S.A.; et al. Fire as a fundamental ecological process: Research advances and frontiers. J. Ecol. 2020, 108, 2047–2069. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pugnaire, F.I. The role of nurse plants in the restoration of degraded environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- Badano, E.I.; Cavieres, L.A. Impacts of ecosystem engineers on community attributes: Effects of cushion plants at different elevations of the Chilean Andes. Div. Distr. 2006, 12, 388–396. [Google Scholar] [CrossRef]
- Bannister, J.A.; Travieso, N.; Acevedo, M.; Acevedo, M.; Puettmann, K.; Salas-Eljatib, C. Shrub influences on seedling performance when restoring the slow-growing conifer Pilgerodendron uviferum in southern bog forests. Restor. Ecol. 2019, 28, 396–407. [Google Scholar] [CrossRef]
- Gomez-Aparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef]
- Gómez-González, S.; Torres-Díaz, C.; Bustos-Schindler, C.; Gianoli, E. Anthropogenic fire drives the evolution of seed traits. Proc. Natl. Acad. Sci. USA 2011, 108, 18743–18747. [Google Scholar] [CrossRef] [PubMed]
- Le Bagousse-Pinguet, Y.; Soliveres, S.; Gross, N.; Torices, R.; Berdugo, M.; Maestre, F.T. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2019, 116, 8419–8424. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cano, J.A.; Goberna, M.; Verdú, M. Data from: Trait based selection of nurse plants to restore ecosystem functions in mine tailings. J. Appl. Ecol. 2018, 55, 1195–1206. [Google Scholar] [CrossRef]
- Svriz, M.; Damascos, M.A.; Zimmermann, H.; Hensen, I. The exotic shrub Rosa rubiginosa as a nurse plant. Implications for the restoration of disturbed temperate forests in Patagonia, Argentina. For. Ecol. Manag. 2013, 289, 234–242. [Google Scholar] [CrossRef]
- Wilgan, B.W.V.; Govender, V.; Forsyth, G.G.; Krajii, T. Towards adaptive fire management for biodiversity conservation: Experience in South African National Parks. Koedoe-Afr. Prot. Area Conserv. Sci. 2011, 53, 1–9. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef]
- UN. Transforming Our World: The 2030 Agenda for Sustainable Development; Governmental Document A/RES/70/1; General Assembly in September 2015; United Nations: New York, NY, USA, 2015; 35p. [Google Scholar]
- Visser, S.; Keesstra, S.; Maas, G.; De Cleen, M. Soil as a basis to create enabling conditions for transitions towards sustainable land management as a key to achieve the SDGs by 2030. Sustainability 2019, 11, 6792. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).