
Fire Has a Positive Effect on the Abundance of Sun Spiders (Arachnida: Solifugae) in the Cerrado-Pantanal Ecotone


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
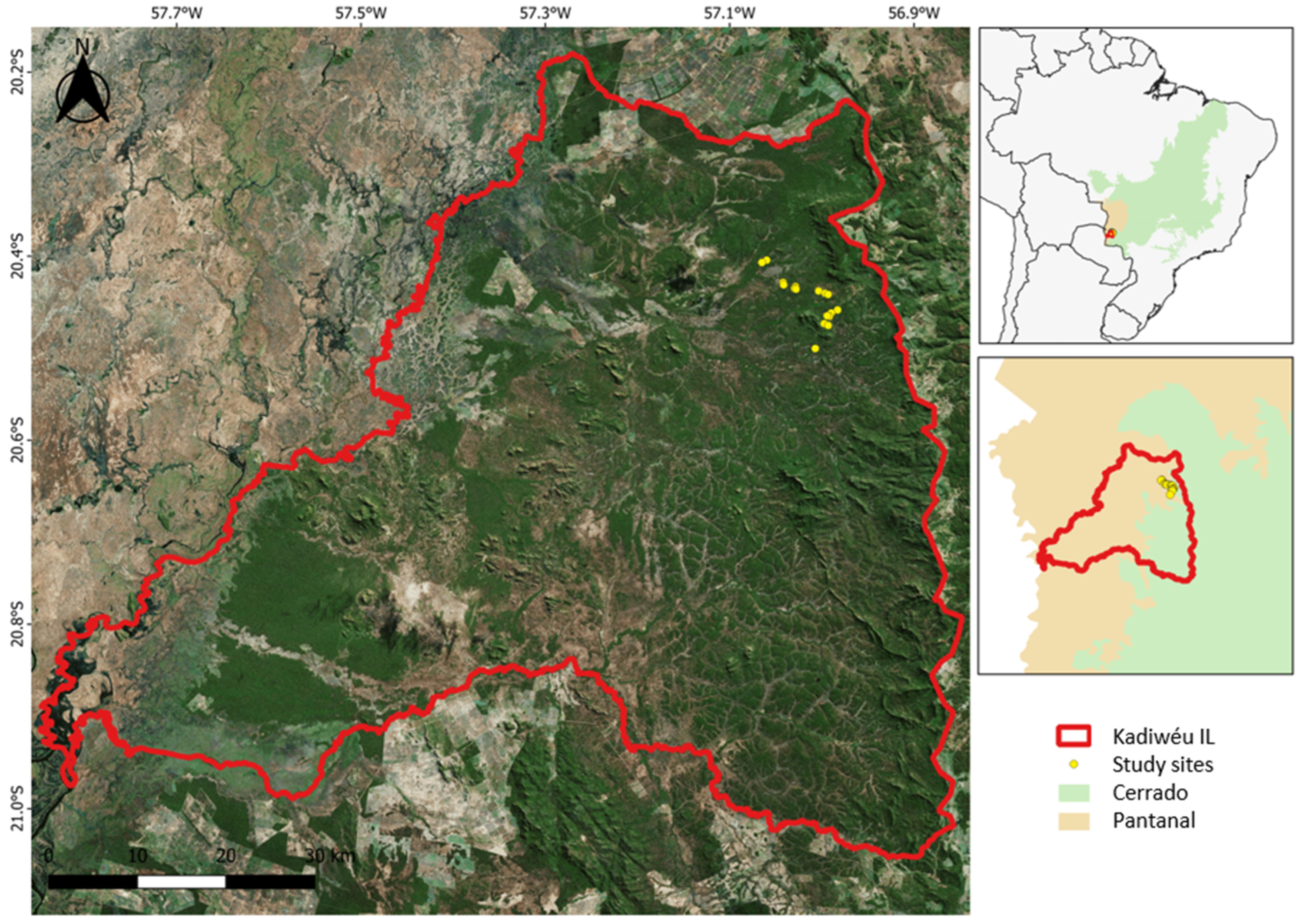
2.1. Study Area
2.2. Experimental Design
2.3. Sun Spiders Sampling
2.4. Data Analysis
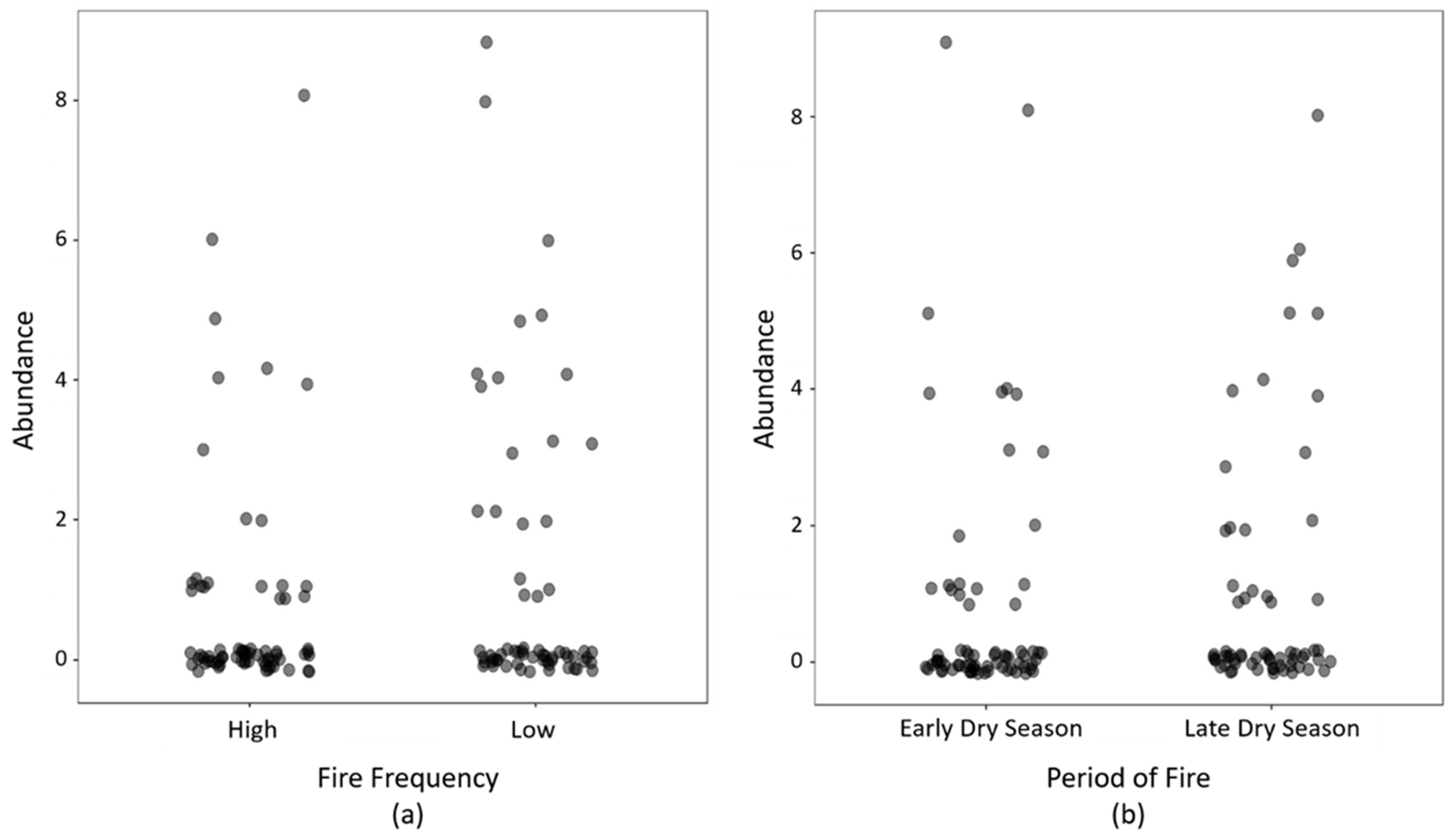
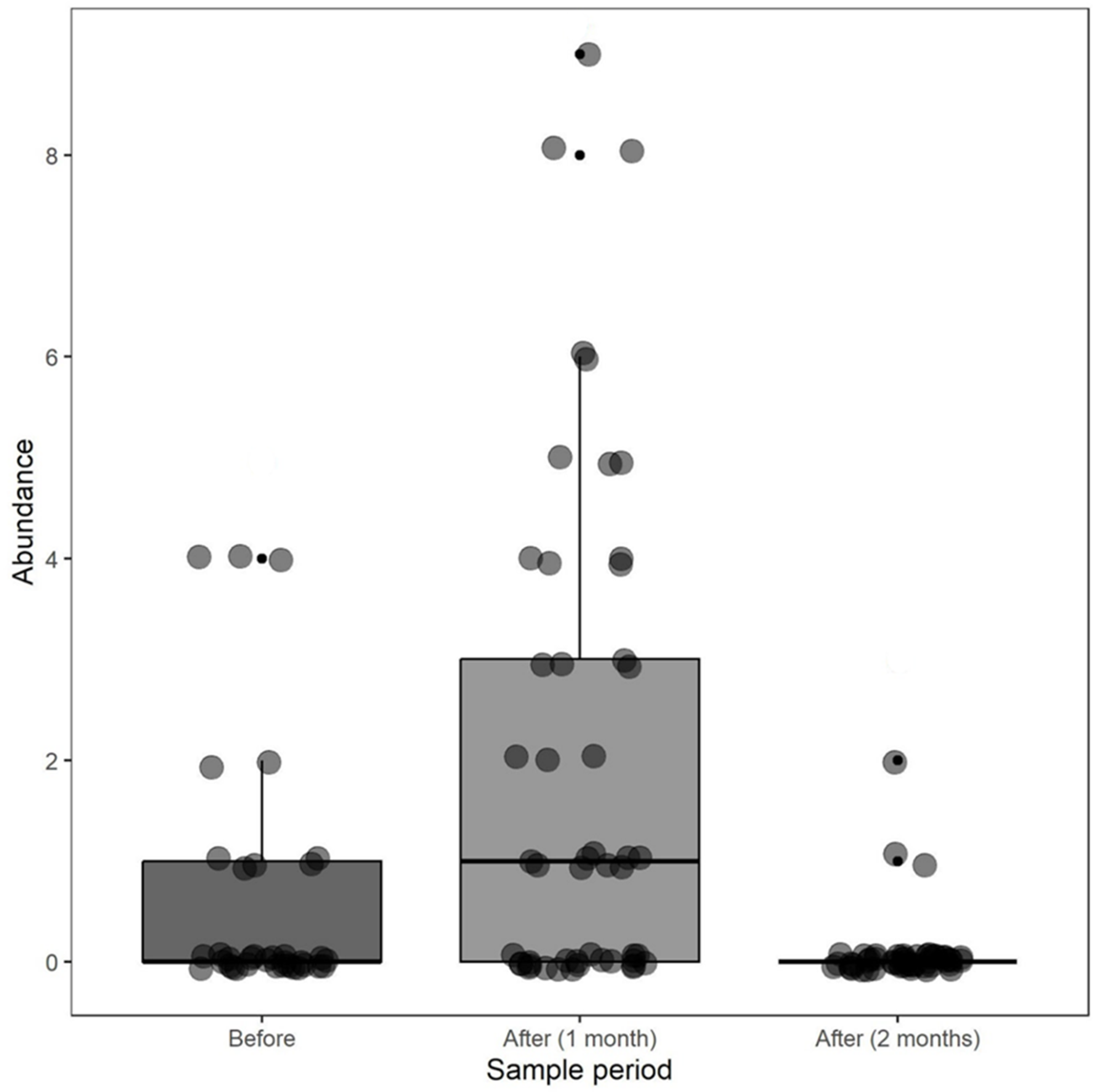
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science (80-) 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, C.N.; Batista, E.K.L. Good Fire, Bad Fire: It Depends on Who Burns. Flora Morphol. Distrib. Funct. Ecol. Plants 2020, 268, 151610. [Google Scholar] [CrossRef]
- Balch, J.K.; Bradley, B.A.; Abatzoglou, J.T.; Chelsea Nagy, R.; Fusco, E.J.; Mahood, A.L. Human-Started Wildfires Expand the Fire Niche across the United States. Proc. Natl. Acad. Sci. USA 2017, 114, 2946–2951. [Google Scholar] [CrossRef]
- Hantson, S.; Andela, N.; Goulden, M.L.; Randerson, J.T. Human-Ignited Fires Result in More Extreme Fire Behavior and Ecosystem Impacts. Nat. Commun. 2022, 13, 2717. [Google Scholar] [CrossRef] [PubMed]
- Hardesty, J.; Myers, R.; Fulks, W. Fire, Ecosystems and People: A Preliminary Assessment of Fire as a Global Conservation Issue. Fire Manag. 2005, 22, 78–87. [Google Scholar]
- Pivello, V.R.; Vieira, I.; Christianini, A.V.; Ribeiro, D.B.; Menezes, L.S.; Berlinck, C.N.; Melo, F.P.L.; Marengo, J.A.; Tornquist, C.G.; Tomas, W.M.; et al. Understanding Brazil’s Catastrophic Fires: Causes, Consequences and Policy Needed to Prevent Future Tragedies. Perspect. Ecol. Conserv. 2021, 19, 233–255. [Google Scholar] [CrossRef]
- Ferreira, B.H.d.S.; Guerra, A.; Oliveira, M.d.R.; Reis, L.K.; Aptroot, A.; Ribeiro, D.B.; Garcia, L.C. Fire Damage on Seeds of Calliandra Parviflora Benth. (Fabaceae), a Facultative Seeder in a Brazilian Flooding Savanna. Plant Species Biol. 2021, 36, 523–534. [Google Scholar] [CrossRef]
- Oliveira, M.R.; Ferreira, B.H.S.; Souza, E.B.; Lopes, A.A.; Bolzan, F.P.; Roque, F.O.; Pott, A.; Pereira, A.M.M.; Garcia, L.C.; Damasceno, G.A.; et al. Indigenous Brigades Change the Spatial Patterns of Wildfires, and the Influence of Climate on Fire Regimes. J. Appl. Ecol. 2022, 59, 1279–1290. [Google Scholar] [CrossRef]
- Damasceno-Junior, G.A.; Roque, F.O.; Garcia, L.C.; Ribeiro, D.B.; Tomas, W.M.; Scremin-Dias, E.; Dias, F.A.; Libonati, R.; Rodrigues, J.A.; Santos, F.L.M.; et al. Lessons to Be Learned from the Wildfire Catastrophe of 2020 in the Pantanal Wetland. Wetl. Sci. Pract. 2021, 38, 107–115. [Google Scholar]
- Buckingham, S.; Murphy, N.; Gibb, H. Effects of Fire Severity on the Composition and Functional Traits of Litter-Dwelling Macroinvertebrates in a Temperate Forest. For. Ecol. Manag. 2019, 434, 279–288. [Google Scholar] [CrossRef]
- Kelly, L.T.; Giljohann, K.M.; Duane, A.; Aquilué, N.; Archibald, S.; Batllori, E.; Bennett, A.F.; Buckland, S.T.; Canelles, Q.; Clarke, M.F.; et al. Fire and Biodiversity in the Anthropocene. Science (80-) 2020, 370, eabb0355. [Google Scholar] [CrossRef]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The Impact of Fire on Soil-Dwelling Biota: A Review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Duarte, M.H.L.; Sousa-Lima, R.S.S.; Young, R.J.; Vasconcelos, M.F.; Bittencourt, E.; Scarpelli, M.D.A.; Farina, A.; Pieretti, N. Changes on Soundscapes Reveal Impacts of Wildfires in the Fauna of a Brazilian Savanna. Sci. Total Environ. 2021, 769, 144988. [Google Scholar] [CrossRef] [PubMed]
- Word Solifugae Catalog. Available online: https://wac.nmbe.ch/order/solifugae/6 (accessed on 21 November 2022).
- Cloudsley-Thompson, J.L. Adaptational Biology of Solifugae (Solpugida). Bull. Br. Arachnol. Soc. 1977, 4, 61–71. [Google Scholar]
- Punzo, F. Dispersion, Temporal Patterns of Activity, and the Phenology of Feeding and Mating Behaviour in Eremobates Palpisetulosus Fichter (Solifugae, Eremobatidae). Bull. Br. Arachnol. Soc. 1997, 10, 303–307. [Google Scholar]
- Harvey, M.S. The Neglected Cousins: What Do We Know about the Smaller Arachnid Orders? J. Arachnol. 2002, 30, 357–372. [Google Scholar] [CrossRef]
- Carvalho, L.S.; Candiani, D.F.; Bonaldo, A.B.; Suesdek, L.; Silva, P.R.R. A New Species of the Sun-Spider Genus Mummucia(Arachnida: Solifugae: Mummucidae) from Piauí, Northeastern Brazil. Zootaxa 2010, 2690, 19–31. [Google Scholar] [CrossRef]
- Iuri, H.A.; Iglesias, M.S.; Ojanguren Affilastro, A.A. A New Species of Chileotrecha Maury, 1987 (Solifugae: Ammotrechidae) from Argentina with Notes on the Genus. Zootaxa 2014, 3827, 20–30. [Google Scholar] [CrossRef]
- Botero-Trujillo, R.; Iuri, H.A. Chileotrecha Romero (Kraus, 1966) Comb. Nov. And Pseudocleobis Patagonicus (Roewer, 1934) Comb. Nov. Transferral from Mummuciidae to Ammotrechidae (Arachnida, Solifugae). Zootaxa 2015, 3990, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Botero-Trujillo, R. The Smallest Known Solifuge: Vempironiella Aguilari, New Genus and Species of Sun-Spider (Solifugae: Mummuciidae) from the Coastal Desert of Peru. J. Arachnol. 2016, 44, 218–226. [Google Scholar] [CrossRef]
- Botero-Trujillo, R.; Ott, R.; Carvalho, L.S. Systematic Revision and Phylogeny of the South American Sun-Spider Genus Gaucha Mello-Leitão (Solifugae: Mummuciidae), with Description of Four New Species and Two New Generic Synonymies. Arthropod Syst. Phylogeny 2017, 75, 3–44. [Google Scholar]
- Botero-Trujillo, R.; Ott, R.; Mattoni, C.I.; Nime, M.F.; Ojanguren-Affilastro, A.A. Two New Species of the Sun-Spider Genus Gaucha from Argentina and Brazil (Solifugae, Mummuciidae). Zootaxa 2019, 4551, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.S.; Botero-Trujillo, R. On the Sun-Spiders of the Ibirapemussu Species-Group of the Genus Gaucha Mello-Leitão, 1924 (Solifugae, Mummuciidae), with Description of a New Species. Zootaxa 2019, 4700, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.F.V.R.; Ferreira, R.L.; Carvalho, L.S. A New Species of the Genus Gaucha Mello-Leito, 1924 from Minas Gerais, Brazil (Solifugae, Mummuciidae). Zootaxa 2021, 5061, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Iuri, H.A.; Ramírez, M.J.; Mattoni, C.I.; Ojanguren-Affilastro, A.A. Revision and Cladistic Analysis of Subfamily Nothopuginae (Solifugae, Ammotrechidae). Zool. Anz. 2021, 295, 126–155. [Google Scholar] [CrossRef]
- Martins, E.G.; Bonato, V.; Machado, G.; Pinto-Da-Rocha, R.; Rocha, L.S. Description and Ecology of a New Species of Sun Spider (Arachnida: Solifugae) from the Brazilian Cerrado. J. Nat. Hist. 2004, 38, 2361–2375. [Google Scholar] [CrossRef]
- Peretti, A.V.; Willemart, R.H. Sexual Coercion Does Not Exclude Luring Behavior in the Climbing Camel-Spider Oltacola Chacoensis (Arachnida, Solifugae, Ammotrechidae). J. Ethol. 2007, 25, 29–39. [Google Scholar] [CrossRef]
- Peretti, A.V.; Vrech, D.E.; Hebets, E.A. Solifuge (Camel Spider) Reproductive Biology: An Untapped Taxon for Exploring Sexual Selection. J. Arachnol. 2021, 49, 299–316. [Google Scholar] [CrossRef]
- Diniz, I.R.; Morais, H.C. Efeitos Do Fogo Sobre Os Insetos Do Cerrado: Consensos e Controvérsias. In Efeitos do Regime do Fogo Sobre a Estrutura de Comunidades de Cerrado: Resultados do Projeto Fogo; Ibama: Brasília, Brazil, 2010; pp. 121–131. [Google Scholar]
- Diniz, I.R.; Higgins, B.; Morais, H.C. How Do Frequent Fires in the Cerrado Alter the Lepidopteran Community? Biodivers. Conserv. 2011, 20, 1415–1426. [Google Scholar] [CrossRef]
- Seyffarth, J.A.; Calouro, A.M.; Price, P.W. Leaf Rollers in Ouratea Hexasperma (Ochnaceae): Fire Effect and the Plant Vigor Hypothesis. Rev. Bras. Biol. 1996, 56, 135–137. [Google Scholar]
- Marquis, R.J.; Diniz, I.R.; Morais, H.C. Patterns and Correlates of Interspecific Variation in Foliar Insect Herbivory and Pathogen Attack in Brazilian Cerrado. J. Trop. Ecol. 2001, 17, 127–148. [Google Scholar] [CrossRef]
- Pott, A.; Oliveira, A.K.M.; Damasceno-Junior, G.A.; Silva, J.S.V. Plant Diversity of the Pantanal Wetland. Braz. J. Biol. 2011, 71, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.S.; Gamarra, R.M.; Mioto, C.L.; Silva, N.M.; Conceição Filho, A.P.; Pott, A. Analysis of the Landscape Complexity and Heterogeneity of the Pantanal Wetland. Braz. J. Biol. 2018, 78, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Chinder, G.B.; Hattas, D.; Massad, T.J. Growth and Functional Traits of Julbernardia Globiflora (Benth) Resprouts and Seedlings in Response to Fire Frequency and Herbivory in Miombo Woodlands. S. Afr. J. Bot. 2020, 135, 476–483. [Google Scholar] [CrossRef]
- Manrique-Pineda, D.A.; de Souza, E.B.; Paranhos Filho, A.C.; Cáceres Encina, C.C.; Damasceno-Junior, G.A. Fire, Flood and Monodominance of Tabebuia Aurea in Pantanal. For. Ecol. Manag. 2021, 479, 118599. [Google Scholar] [CrossRef]
- USGS. Available online: https://lpdaac.usgs.gov/products/mod13q1v006/ (accessed on 26 August 2022).
- Câmara, G.; Souza, R.C.M.; Freitas, U.M.; Garrido, J. Spring: Integrating Remote Sensing and Gis by Object-Oriented Data Modelling. Comput. Graph. 1996, 20, 395–403. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. Available online: https://earthengine.google.com/faq/ (accessed on 27 December 2022). [CrossRef]
- Gamarra, R.M.; Teixeira-Gamarra, M.C.; Carrijo, M.G.G.; Filho, A.C.P. Use of NDVI in Analysis of Structure Vegetation and Protection Effectiveness of Conservation Unit in the Cerrado. RA’E GA-O Espac. Geogr. Anal. 2016, 37, 307–332. [Google Scholar]
- Muma, M.H. Comparison of Three Methods for Estimating Solpugid (Arachnida) Populations. J. Arachnol. 1980, 8, 267–270. [Google Scholar]
- R Core Team. R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 20 November 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Mangiafico, S. Package ‘Rcompanion’. Functions to Support Extension Education Program Evaluation. R package Version 2.4.18 2022; 136p. Available online: https://cran.r-project.org/package=rcompanion (accessed on 23 November 2022).
- Zar, J.H. Bioestatistical Analyses; Pearson Prentice Hall: Hoboken, NJ, USA, 2010. [Google Scholar]
- Bakeman, R. Recommended Effect Size Statistic. Behav. Res. Methods 2005, 37, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Kral, K.C.; Limb, R.F.; Harmon, J.P.; Hovick, T.J. Arthropods and Fire: Previous Research Shaping Future Conservation. Rangel. Ecol. Manag. 2017, 70, 589–598. [Google Scholar] [CrossRef]
- Antunes, S.C.; Curado, N.; Castro, B.B.; Gonçalves, F. Short-Term Recovery of Soil Functional Parameters and Edaphic Macro-Arthropod Community after a Forest Fire. J. Soils Sediments 2009, 9, 267–278. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Forrest, T.G.; Waldrop, T. Short-Term Response of Ground-Dwelling Arthropods to Prescribed Hardwood Forest. For. Sci. 2010, 56, 112–121. [Google Scholar]
- Jansen, R.; Makaka, L.; Little, I.T.; Dippenaar-Schoeman, A. Response of Ground-Dwelling Spider Assemblages (Arachnida, Araneae) to Montane Grassland Management Practices in South Africa. Insect Conserv. Divers. 2013, 6, 572–589. [Google Scholar] [CrossRef]
- Parr, C.L.; Robertson, H.G.; Biggs, H.C.; Chown, S.L. Response of African Savanna Ants to Long-Term Fire Regimes. J. Appl. Ecol. 2004, 41, 630–642. [Google Scholar] [CrossRef]
- van Mantgem, E.F.; Keeley, J.E.; Witter, M. Faunal Responses to Fire in Chaparral and Sage Scrub in California, USA. Fire Ecol. 2015, 11, 128–148. [Google Scholar] [CrossRef]
- Podgaiski, L.R.; Joner, F.; Lavorel, S.; Moretti, M.; Ibanez, S.; Mendonça, M.d.S.; Pillar, V.D. Spider Trait Assembly Patterns and Resilience under Fire-Induced Vegetation Change in South Brazilian Grasslands. PLoS ONE 2013, 8, e60207. [Google Scholar] [CrossRef]
- Bengtsson, J. Disturbance and Resilience in Soil Animal Communities. Eur. J. Soil Biol. 2002, 38, 119–125. [Google Scholar] [CrossRef]
- Moretti, M.; Conedera, M.; Duelli, P.; Edwards, P.J. The Effects of Wildfire on Ground-Active Spiders in Deciduous Forests on the Swiss Southern Slope of the Alps. J. Appl. Ecol. 2002, 39, 321–336. [Google Scholar] [CrossRef]
- Uehara-Prado, M.; Bello, A.D.M.; Fernandes, J.D.O.; Santos, A.J.; Silva, I.A.; Cianciaruso, M.V. Abundance of Epigaeic Arthropods in a Brazilian Savanna under Different Fire Frequencies. Zoologia 2010, 27, 718–724. [Google Scholar] [CrossRef]
- Pressler, Y.; Moore, J.C.; Cotrufo, M.F. Belowground Community Responses to Fire: Meta-Analysis Reveals Contrasting Responses of Soil Microorganisms and Mesofauna. Oikos 2019, 128, 309–327. [Google Scholar] [CrossRef]
- Taiton, N.M.; Mentis, M.T. Fire in Grassland. In Ecological Effects of Fire in South African Ecosystems; Booysen, P.V., Taiton, N.M., Eds.; Springer: Berlin, Germany, 1984; pp. 115–147. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Malmström, A.; Zaitsev, A.S.; Shakhab, S.V.; Bengtsson, J.; Persson, T. Do Burned Areas Recover from inside? An Experiment with Soil Fauna in a Heterogeneous Landscape. Appl. Soil Ecol. 2012, 59, 73–86. [Google Scholar] [CrossRef]
- Howard, D.R.; Hill, P.S.M. The Effect of Fire on Spatial Distributions of Male Mating Aggregations in Gryllotalpa Major Saussure (Orthoptera: Gryllotalpidae) at the Nature Conservancy’s Tallgrass Prairie Preserve in Oklahoma: Evidence of a Fire-Dependent Species. J. Kansas Entomol. Soc. 2007, 80, 51–64. [Google Scholar] [CrossRef]
- Parmenter, R.R.; Kreutzian, M.; Moore, D.I.; Lightfoot, D.C. Short-Term Effects of a Summer Wildfire on a Desert Grassland Arthropod Community in New Mexico. Environ. Entomol. 2011, 40, 1051–1066. [Google Scholar] [CrossRef]
- Anjos, A.G.; Alvarado, S.T.; Sol, M. Patch and Landscape Features Drive Fire Regime in a Brazilian Flammable Ecosystem. J. Nat. Conserv. 2022, 69, 126261. [Google Scholar] [CrossRef]
- Wikars, L.O. Dependence on Fire in Wood-Living Insects: An Experiment with Burned and Unburned Spruce and Birch Logs. J. Insect Conserv. 2002, 6, 1–12. [Google Scholar] [CrossRef]
- Zanzarini, F.V.; Pissarra, T.C.T.; Brandão, F.J.C.; Teixeira, D.D.B. Correlação Espacial Do Índice de Vegetação (NDVI) de Imagem Landsat/ETM+ Com Atributos Do Solo. Rev. Bras. Eng. Agric. Ambient. 2013, 17, 608–614. [Google Scholar] [CrossRef]
- Ribeiro, M.N.; Sanchez, M.; Pedroni, F.; Peixoto, K.d.S. Fogo e Dinâmica Da Comunidade Lenhosa Em Cerrado Sentido Restrito, Barra Do Garças, Mato Grosso. Acta Bot. Bras. 2012, 26, 203–217. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Persson, T. Recovery of Soil Macrofauna after Wildfires in Boreal Forests. Soil Biol. Biochem. 2013, 57, 182–191. [Google Scholar] [CrossRef]
- Zaitsev, A.S.; Gongalsky, K.B.; Persson, T.; Bengtsson, J. Connectivity of Litter Islands Remaining after a Fire and Unburnt Forest Determines the Recovery of Soil Fauna. Appl. Soil Ecol. 2014, 83, 101–108. [Google Scholar] [CrossRef]
- Yekwayo, I.; Pryke, J.S.; Gaigher, R.; Samways, M.J. Wandering Spiders Recover More Slowly than Web-Building Spiders after Fire. Oecologia 2019, 191, 231–240. [Google Scholar] [CrossRef]
- Martínez, F.J.; Cheli, G.H.; Grismado, C.J.; Bisigato, A.J. Ground-Dwelling Arachnids and Fire Disturbance: A Case Study in Northeastern Patagonia (Argentina). Fire 2022, 5, 91. [Google Scholar] [CrossRef]
- Bell, J.R.; Philip Wheater, C.; Rod Cullen, W. The Implications of Grassland and Heathland Management for the Conservation of Spider Communities: A Review. J. Zool. 2001, 255, 377–387. [Google Scholar] [CrossRef]
- Yang, X.; Liu, R.T.; Shao, M.A.; Wei, X.R.; Li, T.C.; Chen, M.Y.; Li, Z.Y.; Dai, Y.C.; Gan, M. Short-Term Effects of Wildfire on Soil Arthropods in a Semi-Arid Grassland on the Loess Plateau. Front. Microbiol. 2022, 13, 989351. [Google Scholar] [CrossRef]
- Koltz, A.M.; Burkle, L.A.; Pressler, Y.; Dell, J.E.; Vidal, M.C.; Richards, L.A.; Murphy, S.M. Global Change and the Importance of Fire for the Ecology and Evolution of Insects. Curr. Opin. Insect Sci. 2018, 29, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Xavier, E.; Rocha, L.S. Autoecology and Description of Mummucia Mauryi (Solifugae, Mummuciidae), a New Solifugae from Brazilian Semi-Arid Caatinga. J. Arachnol. 2001, 29, 127–134. [Google Scholar] [CrossRef]
- Belozerov, V.N. Seasonal Aspects of the Life Cycle of Solifuges (Arachnida, Solifugae) as Compared with Pseudoscorpions (Arachnida, Pseudoscorpiones). Entomol. Rev. 2013, 93, 1050–1072. [Google Scholar] [CrossRef]
- Pausas, J.G. Generalized Fire Response Strategies in Plants and Animals. Oikos 2019, 128, 147–153. [Google Scholar] [CrossRef]
- Warren, S.D.; Scifres, C.J.; Teel, P.D. Response of Grassland Arthropods to Burning: A Review. Agric. Ecosyst. Environ. 1987, 19, 105–130. [Google Scholar] [CrossRef]
- De Araujo Lira, A.F.; Lorenzo, E.P.; Desouza, A.M. The First Record of Gaucha Mauryi (Solifugae: Mummucidae) in the State of Pernambuco, Brazil, and Notes on Brazilian Solifuge Species Distributiont. Entomol. News 2020, 129, 406–413. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site ID | Longitude | Latitude | NDVI | ||
|---|---|---|---|---|---|
| During Fire Season | 10–25 Days after | 30 or More Days after | |||
| 1 | 56°59′36″ W | 20°26′30″ S | 0.330 | 0.204 | 0.400 |
| 2 | 56°59′47″ W | 20°26′24″ S | 0.414 | 0.219 | 0.440 |
| 3 | 57°00′12″ W | 20°26′16″ S | 0.431 | 0.360 | 0.493 |
| 4 | 57°01′47″ W | 20°26′07″ S | 0.376 | 0.392 | 0.330 |
| 5 | 57°01′41″ W | 20°26′09″ S | 0.390 | 0.411 | 0.363 |
| 6 | 57°01′43″ W | 20°25′59″ S | 0.398 | 0.395 | 0.353 |
| 7 | 57°02′31″ W | 20°25′43″ S | 0.328 | 0.312 | 0.255 |
| 8 | 57°02′29″ W | 20°25′52″ S | 0.360 | 0.382 | 0.328 |
| 9 | 56°58′58″ W | 20°27′29″ S | 0.314 | 0.256 | 0.509 |
| 10 | 56°59′23″ W | 20°27′41″ S | 0.336 | 0.250 | 0.550 |
| 11 | 56°59′40″ W | 20°27′53″ S | 0.284 | 0.282 | 0.470 |
| 12 | 56°59′32″ W | 20°27′55″ S | 0.333 | 0.251 | 0.524 |
| 13 | 56°59′36″ W | 20°28′30″ S | 0.273 | 0.273 | 0.477 |
| 14 | 56°59′51″ W | 20°28′24″ S | 0.397 | 0.285 | 0.418 |
| 15 | 57°03′36″ W | 20°24′15″ S | 0.265 | 0.300 | 0.523 |
| 16 | 57°03′54″ W | 20°24′25″ S | 0.256 | 0.237 | 0.434 |
| 17 | 57°00′25″ W | 20°30′01″ S | 0.365 | 0.278 | 0.411 |
| Mean (±SD) | 0.344 (±0.054) | 0.299 (±0.065) | 0.428 (±0.082) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrua, B.A.; Carvalho, L.S.; Teles, T.S.; Oliveira, M.d.R.; Ribeiro, D.B. Fire Has a Positive Effect on the Abundance of Sun Spiders (Arachnida: Solifugae) in the Cerrado-Pantanal Ecotone. Fire 2023, 6, 69. https://doi.org/10.3390/fire6020069
Arrua BA, Carvalho LS, Teles TS, Oliveira MdR, Ribeiro DB. Fire Has a Positive Effect on the Abundance of Sun Spiders (Arachnida: Solifugae) in the Cerrado-Pantanal Ecotone. Fire. 2023; 6(2):69. https://doi.org/10.3390/fire6020069
Chicago/Turabian StyleArrua, Bruno Arguelho, Leonardo Sousa Carvalho, Thiago Silva Teles, Maxwell da Rosa Oliveira, and Danilo Bandini Ribeiro. 2023. "Fire Has a Positive Effect on the Abundance of Sun Spiders (Arachnida: Solifugae) in the Cerrado-Pantanal Ecotone" Fire 6, no. 2: 69. https://doi.org/10.3390/fire6020069
APA StyleArrua, B. A., Carvalho, L. S., Teles, T. S., Oliveira, M. d. R., & Ribeiro, D. B. (2023). Fire Has a Positive Effect on the Abundance of Sun Spiders (Arachnida: Solifugae) in the Cerrado-Pantanal Ecotone. Fire, 6(2), 69. https://doi.org/10.3390/fire6020069