
Short-Term Vegetation Response to Bulldozed Fire Lines in Northern Great Plains Grasslands

Abstract
:1. Introduction
2. Materials and Methods
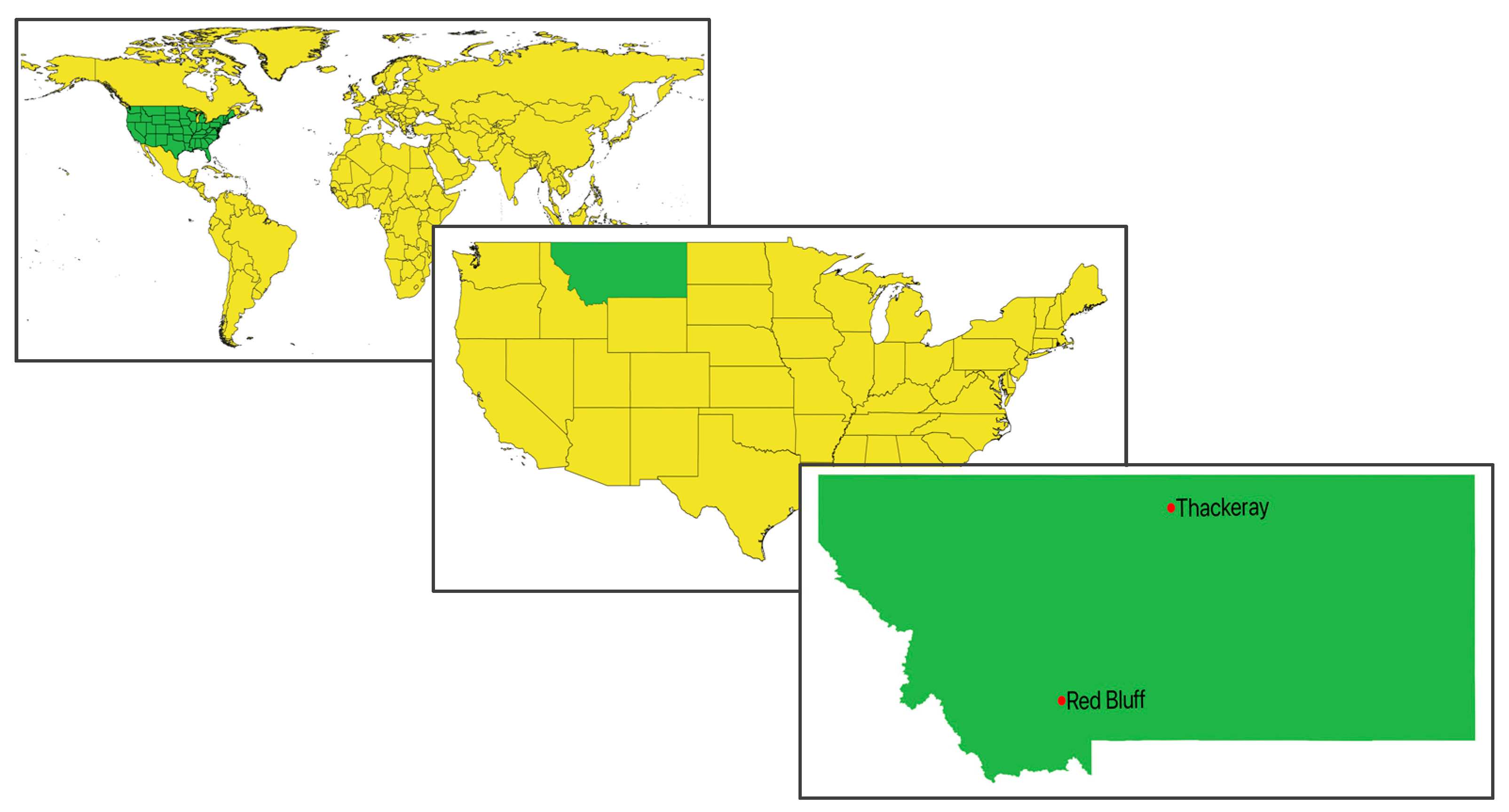
2.1. Study Area
2.2. Sampling Layout
2.3. Data Collection and Analyses
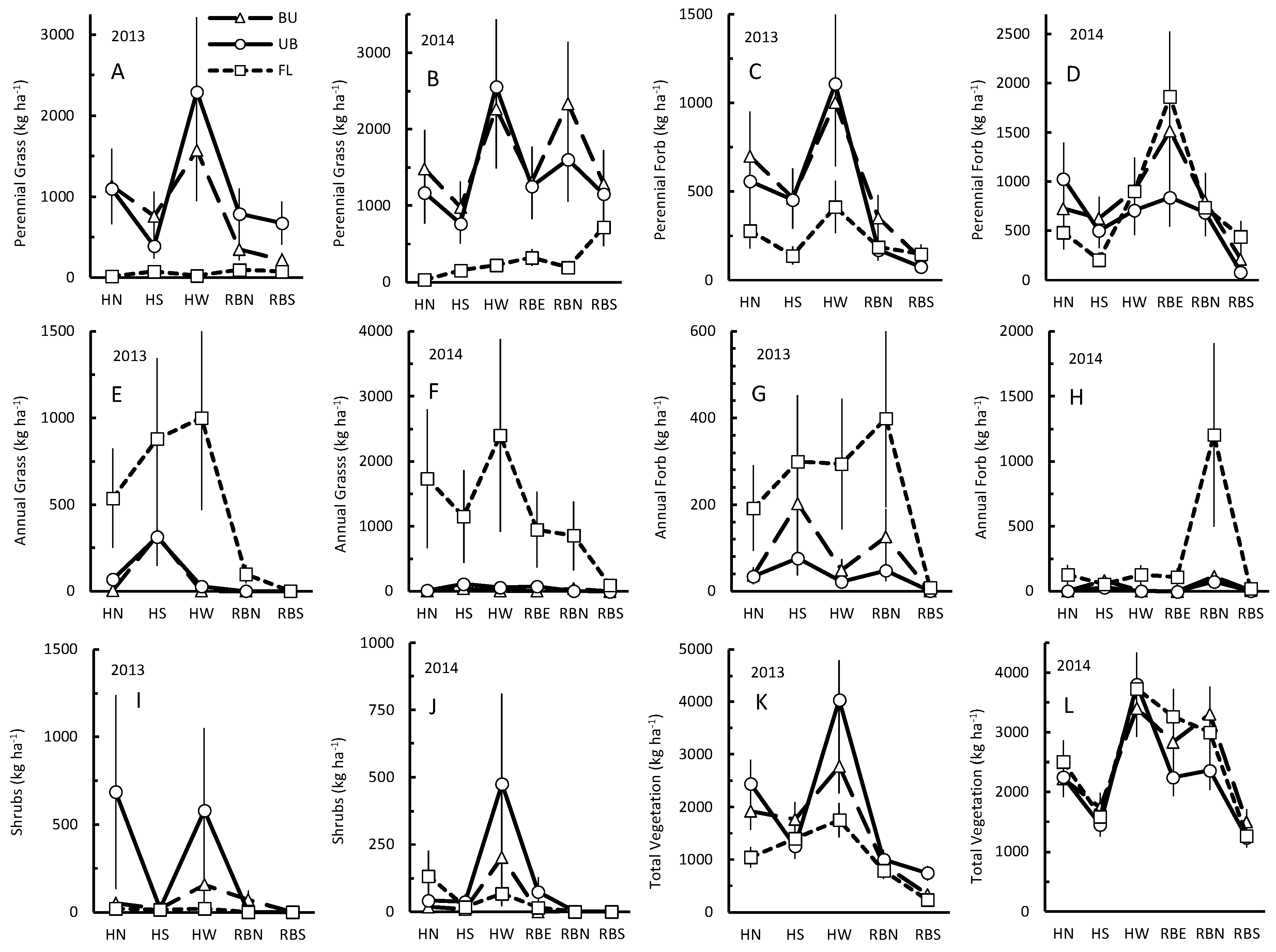
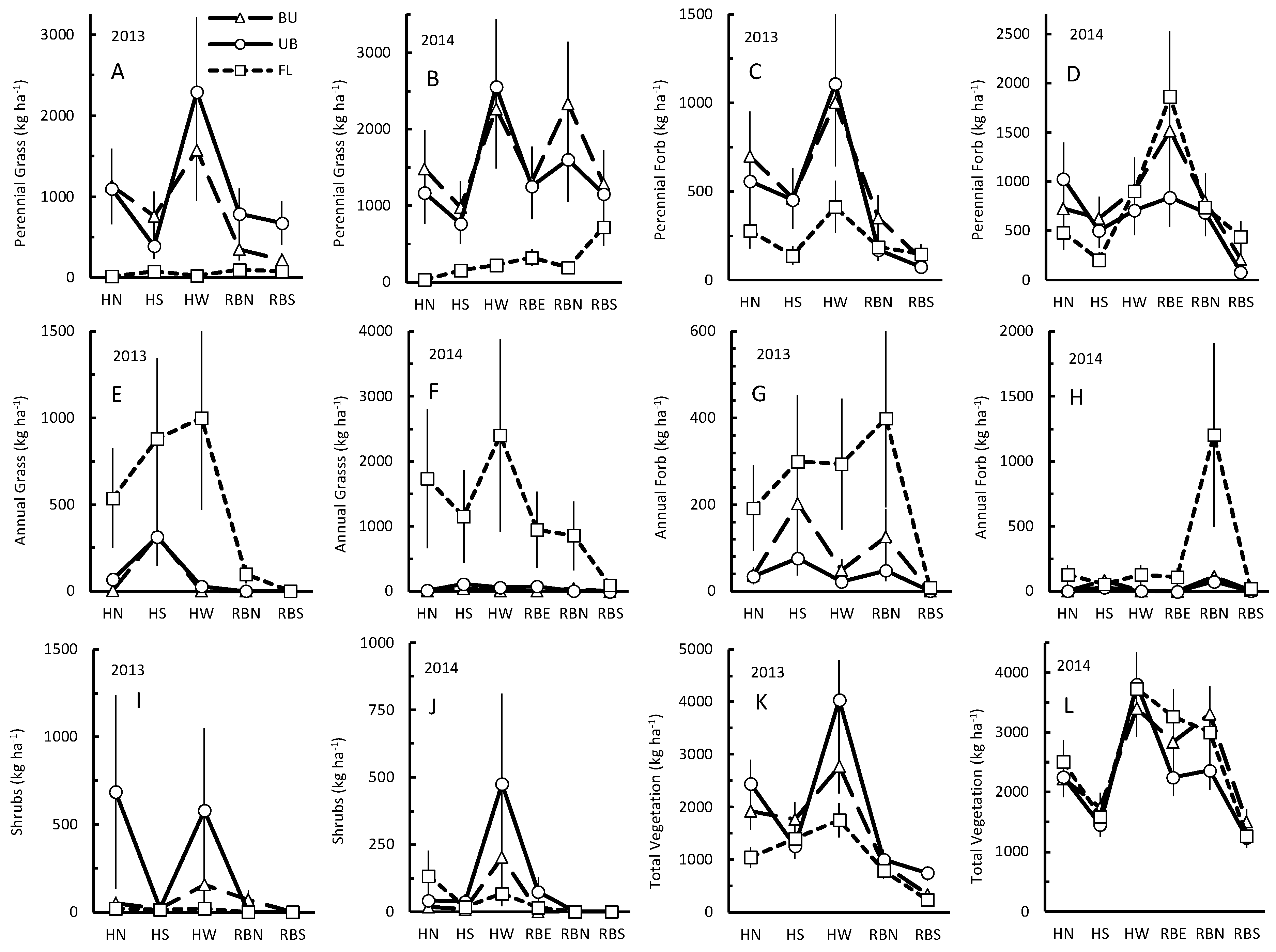
3. Results
3.1. Perennial Grass
3.2. Perennial Forbs
3.3. Annual Grasses
3.4. Annual Forbs

{kind=link}
{kind=link}
| 2013 | 2014 | ||||||
|---|---|---|---|---|---|---|---|
| BU–FL | BU–UB | FL–UB | BU–FL | BU–UB | FL–UB | ||
| Perennial Grass | HN | <0.0001 | 0.9978 | <0.0001 | <0.0001 | 0.8780 | <0.0001 |
| HS | 0.0001 | 0.4666 | 0.0105 | 0.0005 | 0.8688 | 0.0033 | |
| HW | <0.0001 | 0.7817 | <0.0001 | <0.0001 | 0.9679 | <0.0001 | |
| RBE | - | - | - | 0.0108 | 0.9946 | 0.0145 | |
| RBN | 0.0659 | 0.3267 | 0.0007 | <0.0001 | 0.7199 | 0.0001 | |
| RBS | 0.1603 | 0.1147 | 0.0004 | 0.4587 | 0.9750 | 0.5917 | |
| Perennial Forb | HN | 0.1683 | 0.8982 | 0.3607 | 0.6957 | 0.7674 | 0.2885 |
| HS | 0.0491 | 0.9994 | 0.0535 | 0.0727 | 0.9044 | 0.1817 | |
| HW | 0.1907 | 0.9786 | 0.1283 | 0.9996 | 0.8637 | 0.1817 | |
| RBE | - | - | - | 0.9110 | 0.4689 | 0.2522 | |
| RBN | 0.4369 | 0.3211 | 0.9757 | 0.9863 | 0.9484 | 0.9873 | |
| RBS | 0.9240 | 0.6125 | 0.3828 | 0.3061 | 0.1438 | 0.0024 | |
| Annual Grass | HN | <0.0001 | 0.0013 | 0.0177 | <0.0001 | 0.2885 | <0.0001 |
| HS | 0.3764 | 0.9993 | 0.3568 | 0.0006 | 0.5273 | 0.0232 | |
| HW | <0.0001 | 0.0007 | <0.0001 | <0.0001 | 0.0488 | 0.0001 | |
| RBE | - | - | - | <0.0001 | 0.0001 | 0.0095 | |
| RBN | <0.0001 | 0.8797 | <0.0001 | 0.0017 | 0.0011 | <0.0001 | |
| RBS | 1.0000 | 1.0000 | 1.0000 | 0.0001 | 0.8896 | <0.0001 | |
| Annual Forb | HN | 0.0611 | 0.9942 | 0.0474 | <0.0001 | 0.9422 | <0.0001 |
| HS | 0.8552 | 0.3753 | 0.1481 | 0.8840 | 0.4785 | 0.7720 | |
| HW | 0.0378 | 0.5376 | 0.0013 | <0.0001 | 0.8320 | <0.0001 | |
| RBE | - | - | - | <0.0001 | 1.0000 | <0.0001 | |
| RBN | 0.2490 | 0.3884 | 0.0104 | 0.0133 | 0.8706 | 0.0026 | |
| RBS | 0.0383 | 0.9355 | 0.0652 | 0.4420 | 0.8208 | 0.1693 | |
| Shrub | HN | 0.6817 | 0.0565 | 0.0051 | 0.1179 | 0.6791 | 0.4880 |
| HS | 0.9582 | 1.0000 | 0.9582 | 0.8169 | 0.3463 | 0.7110 | |
| HW | 0.1775 | 0.4841 | 0.0098 | 0.5236 | 0.6680 | 0.1276 | |
| RBE | - | - | - | 0.0174 | 0.0004 | 0.2730 | |
| RBN | 0.0016 | 0.0013 | 0.9098 | 1.0000 | 0.7511 | 0.7511 | |
| RBS | 1.000 | 0.9098 | 0.9098 | 0.9515 | 0.7511 | 0.8991 | |
| Total Vegetation | HN | 0.0547 | 0.6427 | 0.0039 | 0.8233 | 0.9980 | 0.8544 |
| HS | 0.6654 | 0.3987 | 0.9016 | 0.8901 | 0.6390 | 0.8978 | |
| HW | 0.1910 | 0.3357 | 0.0047 | 0.8874 | 0.8404 | 0.9949 | |
| RBE | - | - | - | 0.7587 | 0.4706 | 0.1448 | |
| RBN | 0.8736 | 0.9067 | 0.6297 | 0.8785 | 0.2190 | 0.4654 | |
| RBS | 0.3408 | 0.0086 | <0.0001 | 0.6828 | 0.6073 | 0.9922 | |
3.5. Shrubs
3.6. Total Vegetation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyne, S.J.; Andrews, P.L.; Laven, R.D. Introduction to Wildland Fire, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1996; p. 769. [Google Scholar]
- Mohr, F. Light-hand suppression tactics: A fire management challenge. Fire Manag. Notes 1989, 50, 21–23. [Google Scholar]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Merriam, K.E.; Keeley, J.E.; Beyers, J.L. Fuel breaks affect nonnative species abundance in Californian plant communities. Ecol. Appl. 2006, 16, 515–527. [Google Scholar] [CrossRef]
- Young, J.A.; Allen, F.L. Cheatgrass and range science: 1930–1950. J. Range Manag. 1997, 50, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Moroney, J.R.; Rundel, P.W. Effects of fire and fireline disturbance on the plant community in a southern California ecological reserve. Madroño 2013, 60, 173–178. [Google Scholar] [CrossRef]
- National Wildfire Coordinating Group (NWCG). NWCG Glossary of Wildland Fire; National Wildfire Coordinating Group: Washington, DC, USA, 2021. [Google Scholar]
- QGIS.org. QGIS Geographic Information System; QGIS Association: 2022. Available online: http://www.qgis.org (accessed on 29 March 2022).
- Natural Resources Conservation Service [NRCS]. Web Soil Survey. Available online: http://websoilsurvey.sc.egove.usda.gov/App?HomePage.htm (accessed on 30 January 2015).
- PRISM Climate Group. Data Explorer: Time Series Values for Individual Locations. Available online: http://www.prism.oregonstate.edu/explorer/ (accessed on 25 February 2016).
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- O’connor, T. Local extinction in perennial grasslands: A life-history approach. Am. Nat. 1991, 137, 753–773. [Google Scholar] [CrossRef]
- Marlette, G.M.; Anderson, J.E. Seed banks and propagule dispersal in crested-wheatgrass stands. J. Appl. Ecol. 1986, 23, 161–175. [Google Scholar] [CrossRef]
- Iverson, L.R.; Wali, M.K. Buried, viable seeds and their relation to revegetation after surface mining. J. Range Manag. 1982, 35, 648–652. [Google Scholar] [CrossRef]
- Strickler, G.S.; Edgerton, P.J. Emergent seedlings from coniferous litter and soil in eastern Oregon. Ecology 1976, 57, 801–807. [Google Scholar] [CrossRef]
- Baskin, J.; Baskin, C. Ecology of germination and flowering in the weedy winter annual grass Bromus japonicus. J. Range Manag. 1981, 34, 369–372. [Google Scholar] [CrossRef]
- Mack, R.N.; Pyke, D.A. The demography of Bromus tectorum: Variation in time and space. J. Ecol. 1983, 71, 69–93. [Google Scholar] [CrossRef]
- Karl, M.G.; Heitschmidt, R.; Haferkamp, M.R. Vegetation biomass dynamics and patterns of sexual reproduction in a northern mixed-grass prairie. Am. Midl. Nat. 1999, 141, 227–237. [Google Scholar] [CrossRef]
- Harris, G.A. Some competitive relationships between Agropyron spicatum and Bromus tectorum. Ecol. Monogr. 1967, 37, 89–111. [Google Scholar] [CrossRef]
- Whisenant, S.G. Postfire population dynamics of Bromus japonicus. Am. Midl. Nat. 1990, 123, 301–308. [Google Scholar] [CrossRef]
- Cheplick, G.P. Seed dispersal and seedling establishment in grass populations. In Population Biology of Grasses; Cheplick, G.P., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 84–105. [Google Scholar]
- Ozeran, R.; Carr, C.A.; Maxwell, B.D.; Olson, B.E. Bromus tectorum abundance on northern great plains foothills rangelands is related to disturbance, vegetation diversity, and site characteristics. In Proceedings of the future management of grazing and Wild Lands in a High-Tech World: Proceedings 10th International Rangeland Congress, Saskatoon, SK, Canada, 16–22 July 2016. [Google Scholar]
- Ozeran, R. Disturbance and Site Characteristics Relate to Cheatgrass (Bromus tectorum) Abundance on Ranches in Montana Foothills Ecosystems. Master’s Thesis, Montana State University, Bozeman, MT, USA, 2016. [Google Scholar]
- Benson, N.C.; Kurth, L.L. Vegetation establishment on rehabilitated bulldozer lines after the 1988 Red Bench Fire in Glacier National Park. In Symposium on Fire in Wilderness and Park Management; Missoula, M.T., Brown, J.K., Mutch, R.W., Spoon, C.W., Wakimoto, R.H., Eds.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1995; INT-GTR-320; pp. 164–167. [Google Scholar]
- Reisner, M.D.; Grace, J.B.; Pyke, D.A.; Doescher, P.S. Conditions favoruing Bromus tectorum dominance of endangered sagebrush steppe ecosystems. J. Appl. Ecol. 2013, 50, 1039–1049. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, L.D.; Schupp, E.W. Competition as a barrier to establishment of a native perennial grass (Elymus elymoides) in alien annual grass (Bromus tectorum) communities. J. Arid Environ. 2004, 58, 405–422. [Google Scholar] [CrossRef]
- Whisenant, S.G. Changing fire frequencies on Idaho’s Snake River plains: Ecological and management implications. In Symposium on Cheatgrass Invasion, Shrub Die-Off, and Other Aspects of Shrub Biology and Management; McArthur, E.D., Romney, E.M., Smith, S.D., Tueller, P.T., Eds.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1990; INT-GTR-276; pp. 4–10. [Google Scholar]
- Gates, E.; Vermeire, L.T.; Marlow, C.; Waterman, R.C. Fire and season of postfire defoliation effects on biomass, composition, and cover in mixed-grass prairie. Rangel. Ecol. Manag. 2017, 70, 430–436. [Google Scholar] [CrossRef]
- Gates, E.A.; Vermeire, L.T.; Marlow, C.B.; Waterman, R.C. Reconsidering rest following fire: Northern mixed-grass prairie is resilient to grazing following spring wildfire. Agric. Ecosyst. Environ. 2017, 237, 258–264. [Google Scholar]
- Wollstein, K.; O’Connor, C.; Gear, J.; Hoagland, R. Minimize the bad days: Wildland fire response and suppression success. Rangelands 2022. [Google Scholar] [CrossRef]
- Pellant, M. History and applications of the intermountain greenstripping program. In Proceedings Ecology and Management of Annual Rangelands; Monsen, S.B., Kitchen, S.G., Eds.; United States Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1994; INT-GTR-313; pp. 63–68. [Google Scholar]
- Shinneman, D.J.; Germino, M.J.; Pilliod, D.S.; Aldridge, C.L.; Vaillant, N.M.; Coates, P.S. The ecological uncertainty of wildfire fuel breaks: Examples from the sagebrush steppe. Front. Ecol. Environ. 2019, 17, 279–288. [Google Scholar] [CrossRef]
| Perennial Grass | Perennial Forb | Annual Grass | Annual Forb | Shrub | Total Vegetation | |
|---|---|---|---|---|---|---|
| Treatment | ||||||
| 2013 | <0.0001 | 0.1981 | <0.0001 | 0.0209 | 0.0075 | 0.0062 |
| 2014 | <0.0001 | 0.3292 | <0.0001 | <0.0001 | 0.1472 | 0.8053 |
| Site | ||||||
| 2013 | 0.0047 | 0.0023 | <0.0001 | <0.0001 | 0.0099 | <0.0001 |
| 2014 | 0.4152 | 0.0147 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatment × Site | ||||||
| 2013 | 0.0002 | 0.2369 | <0.0001 | 0.8000 | 0.0012 | 0.0248 |
| 2014 | 0.0009 | 0.0416 | 0.0001 | 0.0001 | 0.0264 | 0.8517 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tulganyam, S.; Carr, C.A. Short-Term Vegetation Response to Bulldozed Fire Lines in Northern Great Plains Grasslands. Fire 2022, 5, 79. https://doi.org/10.3390/fire5030079
Tulganyam S, Carr CA. Short-Term Vegetation Response to Bulldozed Fire Lines in Northern Great Plains Grasslands. Fire. 2022; 5(3):79. https://doi.org/10.3390/fire5030079
Chicago/Turabian StyleTulganyam, Samdanjigmed, and Craig A. Carr. 2022. "Short-Term Vegetation Response to Bulldozed Fire Lines in Northern Great Plains Grasslands" Fire 5, no. 3: 79. https://doi.org/10.3390/fire5030079
APA StyleTulganyam, S., & Carr, C. A. (2022). Short-Term Vegetation Response to Bulldozed Fire Lines in Northern Great Plains Grasslands. Fire, 5(3), 79. https://doi.org/10.3390/fire5030079