

Middle Pleistocene Hippopotamuses from the Italian Peninsula: An Overview


 , ,
, , 
Abstract
1. Introduction
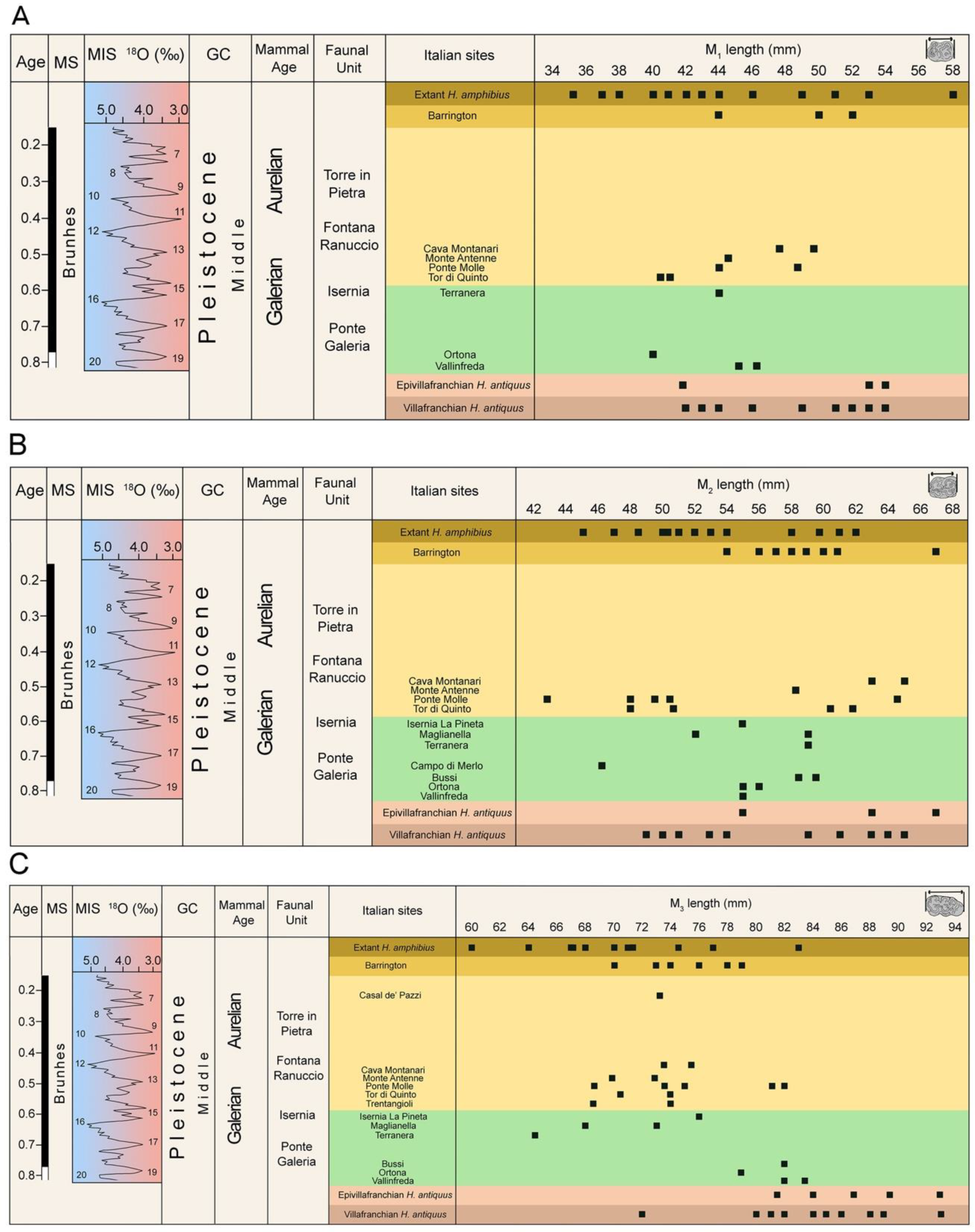
2. An Overview of the Late Early to Late Middle Pleistocene Hippopotamus Record

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Previous Attribution | References for Previous Attribution | Geochronology | Age (Ma) | MIS | References for Chronological Assigment | Analyzed in This Work | Current Attribution |
|---|---|---|---|---|---|---|---|---|
| Cese di Preturo | Hippopotamus antiquus | [26] | late Early Pleistocene | 1.3–1.1 | [26,45] | |||
| Cava Santarelli | Hippopotamus antiquus | [45] | late Early Pleistocene | 1.3–1.1 | [26,45] | |||
| Colle Curti | Hippopotamus antiquus | [26,43] | late Early Pleistocene | 1.1 | [26,43] | |||
| Saticula | Hippopotamus antiquus | [48] | late Early Pleistocene | 1.1 | [48] | |||
| Cesi | Hippopotamus sp. | [49] | late Early Pleistocene | ca 1.0 | [49] | |||
| Colle Lepre | Hippopotamus ex gr. H. antiquus | [34] | late Early Pleistocene | 1.0 | [34] | |||
| Frantoio | Hippopotamus sp. | [50] | late Early Pleistocene | 0.99 | [50] | |||
| Slivia | Hippopotamus sp. | [51,52] | late Early Pleistocene | ca. 0.9 | [51,52] | |||
| Imola | Hippopotamus amphibius (?) | [53] | late Early Pleistocene | 0.85–0.8 | 21/20 | [54] | ||
| Cava Redicicoli | Hippopotamus cf. antiquus | [40] | late Early Pleistocene | ca. 0.8 | 20/19 | [40,55] | ||
| Monte delle Piche | Hippopotamus sp. | [56] | late Early Pleistocene | ca. 0.8 | 20/19 | [56] | ||
| Bussi | Hippopotamus antiquus | [57] | early Middle Pleistocene | 0.78–0.6 | 19–15 | [57] | X | Hippopotamus antiquus |
| Ortona | Hippopotamus antiquus | [34,58] | early Middle Pleistocene | 0.78–0.7 | 19–17 | [34] | ||
| Vallinfreda | Hippopotamus antiquus | [24,59] | early Middle Pleistocene | 0.78–0.7 | 19–17 | [59] | X | Hippopotamus antiquus |
| Campo di Merlo | Hippopotamus antiquus | [36] | early Middle Pleistocene | ca. 0.7 | 18–17 | [60] | X | Hippopotamus antiquus |
| Cava Alibrandi | Hippopotamus sp. | [61] | early Middle Pleistocene | ca. 0.7 | 18–17 | [55] | X | Hippopotamus sp. |
| Cava Arnolfi | Hippopotamus antiquus | [36] | early Middle Pleistocene | ca. 0.7 | 18–17 | [55] | X | Hippopotamus antiquus |
| Monte Oliveto | Hippopotamus sp. cf. H. antiquus | [62] | early Middle Pleistocene | 0.8–0.6 | 19–15 | [62] | ||
| Notarchirico | Hippopotamus antiquus | [15] | early Middle Pleistocene | 0.7–0.65 | 17 | [15] | X | Hippopotamus sp. |
| Pagliare di Sassa | Hippopotamus sp. | [46] | early Middle Pleistocene | ca. 0.7–0.6 | 18–17 | [45,46] | ||
| Borgonuovo | Hippopotamus sp. | [63] | early Middle Pleistocene | ca. 600 | 15 | [64] | ||
| Cava di Breccia—Casal Selce 2 | Hippopotamus antiquus | [2] | early Middle Pleistocene | 0.615–0.611 | 15 | [2,55] | X | Hippopotamus antiquus |
| Isernia La Pineta | Hippopotamus antiquus | [26] | early Middle Pleistocene | ca. 600 | 15 | [65] | X | Hippopotamus antiquus |
| Maglianella | Hippopotamus antiquus | [36] | early Middle Pleistocene | ca. 600 | 15 | [55] | ||
| Terranera | early Middle Pleistocene | ca. 600 | 15 | [66] | X | Hippopotamus antiquus | ||
| Cava Montanari | Hippopotamus amphibius | [22] | Mid Middle Pleistocene | ca. 540–460 | 13 | [22] | ||
| Cava Nera Molinario | Hippopotamus sp. | [55] | mid Middle Pleistocene | ca. 500 | 13 | [55] | X | Hippopotamus sp. |
| Ficoncella | Hippopotamus sp. | [67] | mid Middle Pleistocene | 499–441 | 13 | [67] | ||
| Fosse della Bufala | Hippopotamus cf. antiquus | [68] | mid Middle Pleistocene | ca. 600–500 | 15 | [68] | ||
| Monte Antenne | Hippopotamus amphibius | [69] | Mid Middle Pleistocene | ca. 500 | 13 | [55] | X | Hippopotamus amphibius |
| Osteria Moricone | Hippopotamus cf. antiquus | [68] | mid Middle Pleistocene | ca. 600–500 | 15 | [68] | ||
| Ponte Molle | Hippopotamus ex gr. antiquus | [70] | Mid Middle Pleistocene | ca. 540–460 | 13 | [70] | X | Hippopotamus cf. amphibius |
| Rignano Flaminio | Hippopotamus cf. amphibius | [71] | mid Middle Pleistocene | ca. 500 | 13 | [71] | X | Hippopotamus sp. |
| Tor di Quinto | Hippopotamus sp. and Hippooptamus antiquus | [72] | mid Middle Pleistocene | ca. 500 | 13 | [41] | X | Hippopotamus amphibius |
| Trentangioli | early Middle Pleistocene | ca. 500 | 13 | This work | X | Hippopotamus amphibius | ||
| Castel di Guido | Hippopotamus sp. | [73] | mid Middle Pleistocene | ca. 400 | 11 | [73,74,75] | ||
| Collina Barbattini | Hippopotamus sp. and Hippooptamus antiquus | [60,76,77] | mid Middle Pleistocene | ca. 400 | 11 | [60,75,76] | X | Hippopotamus sp. |
| Fontana Ranuccio | Hippopotamus amphibius | [78] | mid Middle Pleistocene | ca. 400 | 11 | [79] | ||
| Malagrotta | Hippopotamus sp. | [80] | mid Middle Pleistocene | ca. 400 | 11 | [60,75,80] | ||
| Serbaro di Romagnano | Hippopotamus amphibius | [81] | mid Middle Pleistocene | ca. 400 | 11 | [82,83] | X | Hippopotamus sp. |
| Batteria Nomentana | Hippopotamus (Tetraprotodon) major | [84] | late Middle Pleistocene | 0.370–0.290 | 9–8 | [41] | X | Hippopotamus amphibius |
| Sedia del Diavolo | Hippopotamus cf. amphibius | [24] | late Middle Pleistocene | 0.370–0.290 | 9–8 | [41] | X | Hippopotamus amphibius |
| Vigne Torte | Hippopotamus sp. | [72] | Late Middle Pleistocene | 0.370–0.290 | 9–8 | [41] | X | Hippopotamus sp. |
| Villa Chigi | Hippopotamus amphibius | [72] | Late Middle Pleistocene | 0.370–0.290 | 9–8 | This work | X | Hippopotamus sp. |
| Grotta Romanelli—ISU1-2 | Hippopotamus antiquus | [26] | late Middle Pleistocene | 0.320–0.200 | 9–7 | [85] | X | Hippopotamus sp. |
| Casal de’ Pazzi | Hippopotamus cf. amphibius | [86] | late Middle Pleistocene | ca. 240 | 7 | [86] | ||
| Torre del Pagliaccetto—upper level | Hippopotamus sp. | [26,87] | late Middle Pleistocene | 0.270–0.240 | 7 | [88] |
3. Materials and Methods
4. Newly Studied Hippopotamus Material in Italy
4.1. Vallinfreda
4.2. Venosa Basin
4.3. Isernia La Pineta
4.4. Bussi
4.5. Rignano Flaminio
4.6. Serbaro di Romagnano
4.7. Grotta Romanelli
4.8. Rome Area
4.9. Cava Alibrandi, Cava Arnolfi, Campo di Merlo, and Cava di Breccia di Casal Selce
4.10. Cava Nera Molinario, Monte Antenne, Ponte Molle, and Tor di Quinto
4.11. Collina Barbattini—Via Aurelia Km 18.0–19.3
4.12. Batteria Nomentana, Sedia del Diavolo, Vigne Torte, and Villa Chigi
5. Discussion
5.1. Taxonomic Attribution
5.2. Size Variability and Climatic Impact on Hippopotamuses
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gliozzi, E.; Abbazzi, L.; Argenti, P.; Azzaroli, A.; Caloi, L.; Capasso Barbato, L.; Di Stefano, G.; Esu, D.; Ficcarelli, G.; Girotti, O.; et al. Biochronology of selected mammals, molluscs and ostracods from the Middle Pliocene to the Late Pleistocene in Italy. The state of the art. Riv. Ital. Paleontol. Stratig. 1997, 103, 369–388. [Google Scholar]
- Strani, F.; Bellucci, L.; Iannucci, A.; Iurino, D.A.; Mecozzi, B.; Sardella, R. Palaeoenvironments of the MIS 15 site of Cava di Breccia-Casal Selce 2 (central Italian Peninsula) and niche occupation of fossil ungulates during Middle Pleistocene interglacials. Hist. Biol. 2022, 34, 555–565. [Google Scholar] [CrossRef]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene–Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanogr. Paleoclimatol. 2005, 20, PA1003. [Google Scholar]
- Lisiecki, L.E.; Raymo, M.E. Plio-Pleistocene climate evolution: Trends and transition in glacial cycle dynamics. Quat. Sci. Rev. 2007, 26, 56–69. [Google Scholar] [CrossRef]
- Candy, I.; Rose, J.; Lee, J. A seasonally ‘dry’ interglacial climate in eastern England during the early Middle Pleistocene: Palaeopedological and stable isotopic evidence from Pakefield, UK. Boreas 2006, 35, 255–265. [Google Scholar] [CrossRef]
- Candy, I.; Coope, G.R.; Lee, J.R.; Parfitt, S.A.; Preece, R.C.; Rose, J.; Schreve, D.C. Pronounced warmth during early Middle Pleistocene interglacials: Investigating the Mid-Brunhes Event in the British terrestrial sequence. Earth-Sci. Rev. 2010, 103, 183–196. [Google Scholar] [CrossRef]
- Clark, P.; Archer, D.; Pollard, D.; Blum, J.D.; Rial, J.A.; Brovkin, V.; Mix, A.C.; Pisias, N.G.; Roy, M. The Middle Pleistocene transition: Characteristics, mechanisms, and implications for long-term changes in atmospheri pCO2. Quat. Sci. Rev. 2006, 25, 3150–3184. [Google Scholar] [CrossRef]
- Head, M.J.; Gibbard, P.L. Early-Middle Pleistocene transitions: Linking terrestrial and marine realms. Quat. Int. 2015, 389, 7–46. [Google Scholar] [CrossRef]
- Maslin, M.A.; Brierley, C.M. The role of orbital forcing in the early middle Pleistocene transition. Quat. Int. 2015, 389, 47–55. [Google Scholar] [CrossRef]
- Kahlke, R.-D. Late Early Pleistocene European large mammals and the concept of an Epivillafranchian biochron. In Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations, Volume II; Kahlke, R.-D., Maul, L.C., Mazza, P.P.A., Eds.; Cour. Forschungsinst. Senckenberg 259; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 2007; pp. 265–278. [Google Scholar]
- Bellucci, L.; Sardella, R.; Rook, L. Large mammal biochronology framework in Europe at Jaramillo: The Epivillafranchian as a formal biochron. Quat. Int. 2015, 389, 84–89. [Google Scholar] [CrossRef]
- van der Made, J.; Rosell, J.; Blasco, R. Faunas from Atapuerca at the Early-Middle Pleistocene limit: The ungulates from level TD8 in the context of climatic change. Quat. Int. 2017, 433, 296–346. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Kostopoulos, D.S.; Koufos, G.D. Mammuthus meridionalis (Nesti, 1825) from Apollonia-1 (Mygdonia Basin, Northern Greece) and its importance within the Early Pleistocene mammoth evolution in Europe. Geodiversitas 2020, 42, 69–91. [Google Scholar] [CrossRef]
- Laws, R.M. Dentition and ageing of the Hippopotamus. East Afr. Wildl. J. 1968, 6, 19–52. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Santagata, C.; Pereira, A.; Nomade, S.; Voinchet, P.; Bahain, J.-J.; Daujeard, C.; Curci, A.; Lemorini, C.; Hardy, B.; et al. The origin of early Acheulean expansion in Europe 700 ka ago: New findings at Notarchirico (Italy). Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kahlke, R.-D. The Pleistocene of Untermassfeld—A synopsis of site origin, palaeobiodiversity, taphonomic characteristics, palaeoanvironment, chronostratigraphy, and significance in western Palaearctic faunal history. In The Pleistocene of Untermassfeld near Meiningen (Thu¨ringen, Germany). Part 5; Kahlke, R.-D., Ed.; Monographien des Romisch-Germanischen Zentralmuseums: Köln, Germany, 2022; pp. 1635–1734. [Google Scholar]
- Kahlke, R.-D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Ollé, A.; Lombao, D.; Asryan, L.; Garcìa-Medrano, P.; Arroyo, A.; Fernàndez-Marchena, J.L.; Yeşilova, G.C.; Cáceres, I.; Huguet, R.; López-Polín, L.; et al. The earliest European Acheulean: New insights into the large shaped tools from the late Early Pleistocene site of Barranc de la Boella (Tarragona, Spain). Front. Earth Sci. 2023, 11, 1188663. [Google Scholar] [CrossRef]
- Breda, M.; Lister, A.M. Dama roberti, a new species of deer from the early Middle Pleistocene of Europe, and the origins of modern fallow deer. Quat. Sci. Rev. 2013, 69, 155–167. [Google Scholar] [CrossRef]
- Boulbes, N.; Van Asperen, E.N. Biostratigraphy and palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7, 301. [Google Scholar] [CrossRef]
- Iannucci, A.; Mecozzi, B.; Sardella, R. Large mammals from the Middle Pleistocene (MIS 11) site of Fontignano 2 (Rome, central Italy), with an overview of “San Cosimato” assemblages. Alp. Mediterr. Quat. 2021, 34, 155–164. [Google Scholar]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Tentori, D.; Cavasinni, C.; Conti, J.; Messina, M.Y.; Sarra, A.; Sardella, R. Reinforcing the idea of an early dispersal of Hippopotamus amphibius in Europe: Restoration and multidisciplinary study of the skull from the Middle Pleistocene of Cava Montanari (Rome, central Italy). PLoS ONE 2023, 18, e0293405. [Google Scholar] [CrossRef]
- Iannucci, A.; Mecozzi, B.; Sardella, R. Beware of the “Wolf Event”—Remarks on large mammal dispersals in Europe and the late Villafranchian faunal turnover. Alp. Mediterr. Quat. 2023, 36, 1–16. [Google Scholar]
- Caloi, L.; Palombo, M.R.; Petronio, C. Resti cranici di Hippopotamus antiquus (=H. major) e Hippopotamus amphibius conservati nel Museo di Paleontologia dell’Università di Roma. Geol. Rom. 1980, 19, 91–119. [Google Scholar]
- Mazza, P. Interrelations between Pleistocene hippopotami of Europe and Africa. Boll. Soc. Paleont. Ital. 1991, 30, 153–186. [Google Scholar]
- Mazza, P. New evidence on the Pleistocene hippopotami of western Europe. Geol. Rom. 1995, 31, 24–61. [Google Scholar]
- Galobart, A.; Ros, X.; Maroto, J.; Vila, B. Descripción del material de hipopótamo (Hippopotamus antiquus Desmarest, 1822) de los yacimientos del Pleistoceno inferior de Incarcal (Girona, NE de la Península Ibérica). Paleontol. Evol. 2003, 34, 153–173. [Google Scholar]
- Fidalgo, D.; Galli, E.; Madurell-Malapeira, J.; Rosas, A. Earliest Pleistocene European hippos: A review. Comun. Geológicas 2021, 108, 65–69. [Google Scholar]
- Adams, N.F.; Candy, I.; Schreve, D.C. An Early Pleistocene hippopotamus from Westbury Cave, Somerset, England: Support for a previously unrecognized temperate interval in the British Quaternary record. J. Quat. Sci. 2022, 37, 28–41. [Google Scholar] [CrossRef]
- Kahlke, R.-D. Schädelreste von Hippopotamus aus dem Unterpleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen): Teil 2; Kahlke, R.-D., Ed.; Dr Rudolf Habelt GmbH: Bonn, Germany, 2001; pp. 483–500. [Google Scholar]
- Kierdorf, U.; Kahlke, R.-D. Pathological findings on remains of hippopotamids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thüringen, Germany): Part 4.; Kahlke, R.-D., Ed.; Verlag des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; pp. 1251–1272. [Google Scholar]
- Faure, M. Hippopotamus incognitus nov. sp., un hippopotame (Mammalia, Artiodactyla) du Pléistocène d’Europe occidentale. Geobios 1984, 17, 427–437. [Google Scholar] [CrossRef]
- Faure, M. Les hippopotames quaternaires non-insulaires d’Europe occidentale. Nouv. Arch. Mus. Hist. Nat. Lyon 1985, 23, 13–79. [Google Scholar] [CrossRef]
- Mazza, P.P.A.; Bertini, A. Were Pleistocene hippopotamuses exposed to climate-driven body size changes? Boreas 2013, 42, 194–209. [Google Scholar] [CrossRef]
- van der Made, J.; Sahnouni, M.; Boulaghraief, K. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In Proceedings of the II Meeting of African Prehistory, Burgos, Spain, 15–16 April 2015; Sahnouni, M., Semaw, S., Rios Garaizar, J., Eds.; CENIEH: Burgos, Spain, 2017; pp. 135–169. [Google Scholar]
- Petronio, C. Note on the taxonomy of Pleistocene hippopotamuses. Ibex 1995, 3, 53–55. [Google Scholar]
- Martino, R.; Pandolfi, L. The Quaternary Hippopotamus records from Italy. Hist. Biol. 2022, 34, 1146–1156. [Google Scholar] [CrossRef]
- Martino, R.; Rìoz, M.I.; Mateus, O.; Pandolfi, L. Taxonomy chronology and dispersal pattern of Western European Quaternary hippopotamuses Portuguese. Quat. Int. 2023, 674-675, 121–137. [Google Scholar] [CrossRef]
- Fidalgo, D.; Rosas, A.; Madurell-Malapeira, J.; Pineda, A.; Huguet, R.; García-Tabernero, A.; Cáceres, I.; Ollé, A.; Vallverdú, J.; Saladie, P. A review on the Pleistocene occurrences and palaeobiology of Hippopotamus antiquus based on the record from the Barranc de la Boella Section (Francoli Basin NE Iberia). Quat. Sci. Rev. 2023, 307, 108304. [Google Scholar] [CrossRef]
- Mecozzi, B. The Hippopotamus remains from the latest Early Pleistocene site of Cava Redicicoli (Rome, central Italy). Boll. Soc. Paleont. Ital. 2023, 62, 263–279. [Google Scholar]
- Pandolfi, L.; Petronio, C. A brief review of the occurrences of Pleistocene Hippopotamus (Mammalia, Hippopotamidae) in Italy. Geol. Croat. 2015, 68, 313–319. [Google Scholar] [CrossRef]
- Blandamura, F.; Azzaroli, A. L’Ippopotamo Maggiore di Filippo Nesti; serie 8; Atti dell’Accademia Nazionale dei Lincei: Rome, Italy, 1977; Volume 14, pp. 169–188. [Google Scholar]
- Mazza, P.P.A.; Ventra, D. Pleistocene debris-flow deposition of the hippopotamus-bearing Collecurti bonebed (Macerata, Central Italy): Taphonomic and paleoenvironmental analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 296–314. [Google Scholar] [CrossRef]
- Marini, G. Il Lago Pleistocenico Della Conca de L’Aquila; CET: Lanciano, Italy, 1967; 64p. [Google Scholar]
- Mancini, M.; Cavuoto, G.; Pandolfi, L.; Petronio, C.; Salari, L.; Sardella, R. Coupling basin infill history and mammal biochronology in a Pleistocene intramontane basin: The case of western L’Aquila Basin (central Apennines, Italy). Quat. Int. 2012, 267, 62–67. [Google Scholar] [CrossRef]
- Palombo, M.R.; Mussi, M.; Agostini, S.; Barbieri, M.; Di Canzio, E.; Di Rita, F.; Fiore, I.; Iacumin, P.; Magri, D.; Speranza, F.; et al. Human peopling of Italian intramontane basins: The early Middle Pleistocene site of Pagliare di Sassa (L’Aquila, central Italy). Quat. Int. 2010, 223, 170–178. [Google Scholar] [CrossRef]
- Agostini, S.; Rossi, M.A.; Tallini, M. Geologia e paleontologia del Quaternario nel territorio aquilano. In Atti del Convegno “Nei Dintorni di L’Aquila. Ricerche Archeologiche nel Territorio dei Vestini Cismontani Prima e Dopo il Terremoto del 6 Aprile 2009”, Roma, 12–13 Febbraio 2010; Collection de l’Ecole Francaise de Rome: Rome, Italy, 2014; Volume 494, pp. 7–19. [Google Scholar]
- Russo Ermolli, E.; Sardella, R.; Di Maio, G.; Petronio, C.; Santangelo, N. Pollen and mammals from the late Early Pleistocene site of Saticula (Sant’Agata de’ Goti, Benevento, Italy). Quat. Int. 2010, 225, 128–137. [Google Scholar] [CrossRef]
- Ficcarelli, G.; Abbazzi, L.; Albianelli, A.; Bertini, A.; Coltorti, M.; Magnatti, M.; Masini, F.; Mazza, P.; Mezzabotta, C.; Napoleone, G.; et al. Cesi, and early Middle Pleistocene site in Colfiorito Basin (Umbro-Marchean Apennine), central Italy. J. Quat. Sci. 1997, 12, 507–518. [Google Scholar] [CrossRef]
- Bona, F.; Sala, B. Villafranchian-Galerian mammal faunas transition in South-western Europe. The case of the late early Pleistocene mammal fauna of the Frantoio locality, Arda River (Castell’Arquato, Piacenza, northern Italy). Geobios 2016, 49, 329–347. [Google Scholar] [CrossRef]
- Bon, M.; Piccoli, G.; Sala, B. La Fauna Pleistocenica Della Breccia di Slivia (Carso triestino) Nella Collezione del Museo Civico di Storia Naturale di Trieste; Atti del Museo Civico Storia Naturale: Trieste, Italy, 1992; Volume 44, pp. 33–51. [Google Scholar]
- Benussi, B.; Melato, M. Considerazioni preliminari su reperti di una fauna fossile a pachidermi in una breccia ossifera a Slivia-Visogliano. Att. E Mem. Della Comm. Grotte E. Boegan 1970, 10, 77–95. [Google Scholar]
- Azzaroli, A.; Berzi, A. On an upper Villafranchian fauna at Imola, northern Italy, and its correlation with the marine Pleistocene sequence of the Po plain. Paleontogr. Ital. 1970, 66, 1–12. [Google Scholar]
- Muttoni, G.; Scardia, G.; Kent, D.V.; Morsiani, E.; Tremolada, F.; Cremaschi, M.; Peretto, C. First dated human occupation of Italy at~ 0.85 Ma during the late Early Pleistocene climate transition. Earth Planet. Sci. Lett. 2011, 307, 241–252. [Google Scholar] [CrossRef]
- Marra, F.; Pandolfi, L.; Petronio, C.; Di Stefano, G.; Gaeta, M.; Salari, L. Reassessing the sedimentary deposits and vertebrate assemblages from Ponte Galeria area (Rome, central Italy): An archive for the Middle Pleistocene faunas of Europe. Earth-Sci. Rev. 2014, 139, 104–122. [Google Scholar] [CrossRef]
- Pandolfi, L.; Marra, F. Rhinocerotidae (Mammalia, Perissodactyla) from the chrono-stratigraphically constrained Pleistocene deposits of the urban area of Rome (Central Italy). Geobios 2015, 48, 147–167. [Google Scholar] [CrossRef]
- Bosi, C.; Galadini, F.; Giacico, B.; Messina, P.; Sposato, A. Plio-Quaternary continental deposits in the Latium-Abruzzi apennines: The correlation of geological events across different intermontane basins. Il Quat. Ital. J. Quat. Sci. 2003, 16, 55–76. [Google Scholar]
- Agostini, S.; Bertini, A.; Caramiello, S.; De Flaviis, A.G.; Mazza, P.; Rossi, M.A.; Satolli, S. A new mammalian bone bed from the lower Middle Pleistocene of Ortona (Chieti, Abruzzo, central Italy). In Proceedings of the Giornate di Paleontologia; Coccioni, R., Marsili, A., Eds.; Grzybowski Foundation: Kraków, Poland, 2007; Volume 12, pp. 1–5. [Google Scholar]
- Fabbi, S.; Romano, M.; Strani, F.; Sardella, R.; Bellucci, L. The Pleistocene vertebrate fauna of the Oricola-Carsoli intermontane Basin (Latium-Abruzzi, Italy): State of the art and historical review. Boll. Soc. Paleont. Ital. 2021, 60, 255–268. [Google Scholar]
- Ceruleo, P.; Rolfo, M.F.; Petronio, C.; Salari, L. Review of Middle Pleistocene archaeological and biochronological data in Malagrotta—Ponte Galeria area (Rome, Central Italy). Atti Della Soc. Toscana Di Sci. Nat. Mem. Ser. A 2021, 128, 85–103. [Google Scholar]
- Capasso Barbato, L.; Petronio, C. Nuovi resti di mammiferi del Pleistocene medio-inferiore di Ponte Galeria (Roma). Boll. Serv. Geol. d’Ital. 1983, 104, 157–176. [Google Scholar]
- Berzi, A. An early Middle Pleistocene fauna at Monte Oliveto (S. Giaminiano, Siena, Italy). Palaeontogr. Ital. 1972, 68, 28–33. [Google Scholar]
- Azzaroli, A. A skull of the Giant deer Megaceros verticornis from Eastern Tuscany. Att. Della Accad. Naz. Dei Lincei. Cl. Sci. Fis. Mat. E Naturali. Rend. Ser. 8 1976, 61, 485–487. [Google Scholar]
- Palombo, M.R.; Azanza, B.; Alberdi, M.T. Italian Mammal Biochronology from the latest Miocene to the Middle Pleistocene: A multivariate approach. Geol. Romana 2003, 36, 355–368. [Google Scholar]
- Coltorti, M.; Feraud, G.; Marzoli, A.; Peretto, C.; Ton-That, Y.; Voinchet, P.; Bahain, J.J.; Minelli, A.; Thun Hohenstein, U. New 40Ar/39 Ar, stratigraphic and palaeoclimatic data on the Isernia La Pineta Lower Palaeolithic site, Molise, Italy. Quat. Int. 2005, 131, 11–22. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, M.R. Megaceros solilhacus Robert, da Terranera (Bacino di Venosa, Potenza). Quaternaria 1979, 21, 129–2138. [Google Scholar]
- Aureli, D.; Rocca, R.; Lemorini, C.; Modesti, V.; Scaramucci, S.; Milli, S.; Giaccio, B.; Marano, F.; Palombo, M.R.; Contardi, A. Mode 1 or mode 2? “Small tools” in the technical variability of the European Lower Palaeolithic: The site of Ficoncella (Tarquinia, Lazio, central Italy). Quat. Int. 2016, 393, 169–184. [Google Scholar] [CrossRef]
- Marra, F.; Ceruleo, P.; Jichac, B.; Pandolfi, L.; Petronio, C.; Salari, L.; Giaccio, B.; Sottili, G. Glacio-eustatic and tectonic forcing on the lacustrine succession of the Cretone basin: Chronostratigraphic constraints to Acheulian industry and Middle Pleistocene faunal assemblages of Latium (central Italy). J. Quat. Sci. 2016, 31, 641–658. [Google Scholar] [CrossRef]
- Clerici, E. Resti di ippopotamo al Monte Antene. Boll. Soc. Geol. Ital. 1926, 45, 118–122. [Google Scholar]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Sardella, R. Redefining Ponte Molle (Rome, central Italy): An important locality for Middle Pleistocene mammal assemblages of Europe. Alp. Mediterr. Quat. 2021, 34, 131–154. [Google Scholar]
- Petronio, C.; Di Stefano, G.; Kotsakis, T.; Salari, L.; Marra, F.; Jicha, B.R. Biochronological framework for the late Galerian and early-middle Aurelian Mammal Ages o peninsular Italy. Geobios 2019, 53, 35–50. [Google Scholar] [CrossRef]
- Di Stefano, G.; Petronio, C.; Sardella, R. Biochronology of Pleistocene mammal faunas from Rome urban area. Il Quat. 1998, 11, 191–199. [Google Scholar]
- Sala, B.; Barbi, G. Descrizione della fauna. In Gli Scavi a Castel di Guido. Il Più Antico Giacimento di Cacciatori Nell’agro Romano; Radmilli, A.M., Boschian, G., Eds.; ETS: Pisa, Italy, 1996; pp. 55–91. [Google Scholar]
- Villa, P.; Boschian, G.; Pollarolo, L.; Saccà, D.; Marra, F.; Nomade, S.; Pereira, A. Elephant bones for the Middle Pleistocene toolmaker. PloS ONE 2021, 16(8), e0256090. [Google Scholar] [CrossRef] [PubMed]
- Marra, F.; Pereira, A.; Boschian, G.; Nomade, S. MIS 13 and MIS 11 aggradational successions of the Paleo-Tiber delta: Geochronological constraints to sea-level fluctuations and to the Acheulean sites of Castel di Guido and Malagrotta (Rome, Italy). Quat. Int. 2022, 616, 1–11. [Google Scholar] [CrossRef]
- Anzidei, A.P.; Caloi, L.; Giacopini, L.; Mantero, D.; Palombo, M.R.; Sebastiani, R.; Segre, A.G. Saggi di scavo nei depositi pleistocenici del Km 18,900 della Via Aurelia e di Collina Barbattini (Castel di Guido-Roma). Quad. Di Archeol. Etrusco-Ital. 1993, 21, 81–90. [Google Scholar]
- Capasso Barbato, L.; Petronio, C. La mammalofauna pleistocenica di Castel di Guido (Roma). Boll. Serv. Geol. d’Ital. 1981, 102, 95–108. [Google Scholar]
- Strani, F.; Demiguel, D.; Bona, F.; Sardella, R.; Biddittu, I.; Bruni, L.; De Castro, A.; Guadagnoli, F.; Bellucci, L. Ungulate dietary adaptations and palaeoecology of the Middle Pleistocene site of Fontana Ranuccio (Anagni, Central Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 496, 238–247. [Google Scholar] [CrossRef]
- Grimaldi, S.; Santaniello, F.; Angelucci, D.E.; Bruni, L.; Parenti, F. A Techno-Functional Interpretation of the Lithic Assemblage from Fontana Ranuccio (Anagni, Central Italy): An Insight into a MIS 11 Human Behaviour. J. Paleolit. Archaeol. 2020, 3, 944–966. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, P. Resti di mammiferi del Pleistocene Medio di Malagrotta (Roma). Boll. Ser. Geol. d’Ital. 1979, 100, 141–188. [Google Scholar]
- Pasa, A. I mammiferi di alcune antiche brecce veronesi. Mem. Museo Civ. St. Nat. Verona 1947, 1, 1–111. [Google Scholar]
- Caloi, L.; Palombo, M.R. Osservazioni sugli equidi italiani del Pleistocene medio inferiore. Geol. Rom. 1990, 26, 187–221. [Google Scholar]
- Caloi, L.; Palombo, M.R. Gli equidi del Pleistocene medio-inferiore di Soave il Castello, Monte Tenda e Romagnano il Serbaro (Verona, Italia). Boll. Mus. Civ. St. Nat. Verona 1991, 15, 447–468. [Google Scholar]
- Meli, R. Sopra alcune ossa fossili rinvenute nelle ghiaie allivionali presso la via Nomentana, al 3° chilom. da Roma. Boll. Reg. C. Geol. d’Ital. 1886, 17, 265–280. [Google Scholar]
- Pieruccini, P.; Forti, L.; Mecozzi, B.; Iannucci, A.; Yu, T.L.; Shen, C.-C.; Bona, F.; Lembo, G.; Muttillo, B.; Sardella, R.; et al. Stratigraphic reassessment of Grotta Romanelli sheds light on Middle-Late Pleistocene palaeoenvironments and human settling in the Mediterranean. Sci. Rep. 2022, 12, 13530. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, L.; Martino, R.; Palombo, M.R. New insights on the fossil mammals from Casal de’Pazzi (Rome). J. Mediterr. Earth Sci. 2023, 15. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, M.R. Anfibi, rettili e mammiferi di Torre del Pagliaccetto (Torre in Pietra, Roma). Quaternaria 1978, 20, 315–428. [Google Scholar]
- Villa, P.; Soriano, S.; Grün, R.; Marra, F.; Nomade, S.; Pereira, A.; Boschian, G.; Pollarolo, L.; Fang, F.; Bahain, J.-J. The Acheulian and Early Middle Paleolithic in Latium (Italy): Stability and Innovation. PLoS ONE 2016, 11, e0160516. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.A.; Tongiorgi, E.; Trevisan, L. Giacimento di fauna ad Ippopotamo nel quaternario antico della Maglianella presso Roma. Att. Della Soc. Ital. Per Il Prog. Delle Sci. 1951, 42, 505–506. [Google Scholar]
- Fabiani, R.; Maxia, C. L’Istituto e i Musei di Geologia e Paleontologia; Pubblicazioni Università degli Studi di Roma Istituto di Geologia e Paleontologia; Istituto grafico tiberino: Roma, Italy, 1953; Volume 9, pp. 1–31. [Google Scholar]
- Radmilli, A.M.; Boschian, G. Gli Scavi a Castel di Guido. Il più Antico Giacimento di Cacciatori Nell’agro Romano; ETS: Pisa, Italy, 1996. [Google Scholar]
- Rubini, M.; Cerroni, V.; Festa, G.; Sardella, R.; Zaio, P. A revision of hominin fossil teeth from Fontana Ranuccio (middle Pleistocene, Anagni, Frosinone, Italy). J. Hum. Evol. 2014, 77, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Biddittu, I.; Cassoli, P.F.; Radicati di Brozolo, F.; Segre, A.G.; Segre Naldini, E.; Villa, I. Anagni a K/Ar dated Lower Middle Pleistocene site, Central Italy: Preliminary report. Quaternaria 1979, 21, 53–71. [Google Scholar]
- Gioia, P.; Silvestri, L.; Zanzi, G.L. Introduction: Casal de’ Pazzi: The Pleistocene on the outskirts between past, present, and future. The reasons for a conference. J. Mediterr. Earth Sci. 2023, 15. [Google Scholar]
- Osborn, H.F. Palaeoloxodon antiquus italicus sp. nov., final stage in the ‘Elephas antiquus’ phylum. Am. Mus. Novit. 1931, 460, 1–24. [Google Scholar]
- De Lorenzo, G.; D’Erasmo, G.D. Avanzi di ippopotamo nell’Italia meridionale. Att. R. Acc. Sci. Fis. Mat. Napoli 1935, 1, 1–11. [Google Scholar]
- Cavinato, G.P.; Petronio, C.; Sardella, R. The mercure river basin (southern Italy): Quaternary stratigraphy and large mammal biochronology. World Elephants Proc. 1st Int. Congr. Rome 2001, 187–190. [Google Scholar]
- Airaghi, C. L’ippopotamo (Hippopotamus amphibius L.) dell’antico lago di Mércure (Calabria). Att. Soc. Ital. Sci. Nat. E Mus. Civ. Stor. Nat. Milano 1922, 9, 408–418. [Google Scholar]
- Leonardi, P. Nuovi resti di Ippopotamo nelle ligniti del Mercure. Studi Trentini Sci. Nat. 1932, 14, 3–6. [Google Scholar]
- Petrosino, P.; Russo Ermolli, E.; Donato, P.; Jicha, B.; Robustelli, G.; Sardella, R. Using Tephrochronology and palynology to date the MIS 13 lacustrine sediments of the Mercure Basin (Southern Apennines–Italy). Ital. J. Geosci. 2014, 133, 169–186. [Google Scholar] [CrossRef]
- Ponzi, G. Le Ossa Fossili Subappennine dei Dintorni di Roma; Serie III; Atti della Reale Accademia dei Lincei: Roma, Italy, 1878; Volume 2, pp. 1–30. [Google Scholar]
- Lefévre, D.; Raynal, J.-P.; Vernet, G.; Kieffer, G.; Piperno, M. Tephrostratigraphy and the age of ancient Southern Italian Acheulean settlements: The sites of Loreto and Notarchirico (Venosa, Basilicata, Italy). Quat. Int. 2010, 223-224, 360–368. [Google Scholar] [CrossRef]
- Piperno, M. Notarchirico, Un sito del Pleistocene Medio Iniziale nel Bacino di Venosa; Edizioni Osanna: Venosa, Italy, 1999. [Google Scholar]
- Pereira, A.; Nomade, S.; Voinchet, P.; Bahain, J.J.; Falguères, C.; Garon, H.; Lefévre, D.; Raynal, J.P.; Scao, V.; Piperno, M. The earliest securely dated hominin fossil in Italy and evidence of Acheulian occupation during glacial MIS 16 at Notarchirico (Venosa, Basilicata, Italy). J. Quat. Sci. 2015, 30, 639–650. [Google Scholar] [CrossRef]
- Moncel, M.H.; Lemorini, C.; Eramo, G.; Fioretti, G.; Daujeard, C.; Curci, A.; Berto, C.; Hardy, B.; Pineda, A.; Rineau, V.; et al. A taphonomic and spatial distribution study of the new levels of the middle Pleistocene site of Notarchirico (670–695 ka, Venosa, Basilicata, Italy). Archaeol. Anthropol. Sci. 2023, 15, 106. [Google Scholar] [CrossRef]
- Ferrara, F.; Piperno, M. Località di interesse preistorico nel bacino di Venosa. Collezioni, ritrovamenti isolati e scavi. In Notarchirico, Un Sito del Pleistocene Medio Iniziale nel Bacino di Venosa; Piperno, M., Ed.; Edizioni Osanna: Venosa, Italy, 1999; pp. 41–66. [Google Scholar]
- A cura di Colella, A.; Schiattarella, M.; Giannandrea, P.; Lazzari, M.; Longhitano, S.G.; Vitale, G. APAT-SERVIZIO GEOLOGICO D’ITALIA-Carta Geologica d’Italia Alla Scala 1:50.000 e Relative Note Illustrative. Foglio 452 Rionero in Vulture. Serv. Geol. d’Ital. 2018, 1–95. The Documents Are Available for Consultation on the Website. Available online: www.apat.it (accessed on 19 February 2024).
- Rellini, U. Sulle Stazioni Quaternarie di Tipo “Chelléen” Del1′agro Venosino; Tipografia della R. Accademia dei Lincei address: Roma, Italy, 1915; Volume 2, pp. 183–210. [Google Scholar]
- Peretto, C.; Arnaud, J.; Moggi-Cecchi, J.; Manzi, G.; Nomade, S.; Pereira, A.; Falguéres, C.; Bahain, J.J.; Grimaud-Hervé, D.; Berto, C.; et al. A Human Deciduous Tooth and New 40Ar/39Ar Dating Results from the Middle Pleistocene Archaeological Site of Isernia La Pineta, Southern Italy. PLoS ONE 2015, 10, e0140091. [Google Scholar] [CrossRef]
- Sala, B. Le nuove specie rinvenute a La Pinet. In Preistoria in Molise, gli Insediamenti del Territorio di Isernia; Peretto, C., Ed.; Centro Europeo di Ricerche Preistoriche, Collana Ricerche 3; Aracne editrice: Pomezia, Italy, 2006; pp. 36–38. [Google Scholar]
- Breda, M.; Thun-Hohenstein, U.; Peretto, C. The deer from the early Middle Pleistocene site of Isernia La Pineta (Molise, Italy): Revised identifications and new remains from the last 10 years of excavation. Geol. J. 2015, 50, 290–305. [Google Scholar] [CrossRef]
- Karner, D.B.; Marra, F.; Renne, P.R. The history of the Monti Sabatini and Alban Hills volcanoes: Groundwork for assessing volcani-tectonic hazards for Rome. J. Volcanol. Geotherm. Res. 2001, 107, 185–219. [Google Scholar] [CrossRef]
- Bon, M.; Piccoli, G.; Sala, B. I giacimenti Quaternari di vertebrati fossili nell’Italia Nord-Orientale. Mem. Di Sci. Geol. 1991, 18, 185–231. [Google Scholar]
- Sardella, R.; Mazzini, I.; Giustini, F.; Mecozzi, B.; Brilli, M.; Iurino, D.A.; Lembo, G.; Muttillo, B.; Massussi, M.; Sigari, D.; et al. Grotta Romanelli (Southern Italy, Apulia): Legacies and issues in excavating a key site for the Pleistocene of the Mediterranean. Riv. Ital. Paleontol. Stratigr. 2018, 124, 247–264. [Google Scholar]
- Sardella, R.; Iurino, D.A.; Mecozzi, B.; Sigari, D.; Bona, F.; Bellucci, L.; Coltorti, M.; Conti, J.; Lembo, G.; Muttillo, B.; et al. Grotta Romanelli (Lecce, Southern Italy) Between Past and Future: New Studies and Perspectives for an Archaeo-geosite Symbol of the Palaeolithic in Europe. Geoheritage 2019, 11, 1413–1432. [Google Scholar] [CrossRef]
- Mecozzi, B.; Iannucci, A.; Bona, F.; Mazzini, I.; Pieruccini, P.; Sardella, R. Rediscovering Lutra lutra from Grotta Romanelli (southern Italy) in the framework of the puzzling evolutionary history of Eurasian otter. PalZ 2022, 96, 161–174. [Google Scholar] [CrossRef]
- Mecozzi, B.; Buzi, C.; Iannucci, A.; Micarelli, I.; Bona, F.; Forti, L.; Lembo, G.; Manzi, G.; Mazzini, I.; Muttillo, B.; et al. New human fossil from the latest Pleistocene Pieruccini of Grotta Romanelli (Apulia, southern Italy). Archaeol. Anthropol. Sci. 2022, 14, 27. [Google Scholar] [CrossRef]
- Blanc, G.A.; Grotta Romanelli, I. Stratigrafia dei depositi e natura e origine di essi. Arch. Per L’antropologia E La Etnol. 1920, 50, 1–39. [Google Scholar]
- Blanc, G.A. Grotta Romanelli II. Dati ecologici e paletnologici. Arch. Per L’antropologia E La Etnol. 1928, 58, 1–49. [Google Scholar]
- Fornaca-Rinaldi, G. Il metodo 230Th/238U per la datazione di stalattiti e stalagmite. Boll. Geofis. Teor. Appl. 1968, 10, 3–14. [Google Scholar]
- Fornaca-Rinaldi, G. 230Th/234Th dating of cave concretions. Earth Planet. Sci. Lett. 1968, 5, 120–122. [Google Scholar] [CrossRef]
- Fornaca-Rinaldi, G.; Radmilli, A.M. Datazione con il metodo 230Th/238U di stalagmiti contenute in depositi musteriani. Att. Della Soc. Toscana Sci. Nat. 1968, 75, 639–646. [Google Scholar]
- Romano, M.; Mecozzi, B.; Sardella, R. The Quaternary paleontological research in the Campagna Romana (Central Italy) at the 19th-20th century transition: Historical overview. Alp. Mediterr. Quat. 2021, 34, 1–21. [Google Scholar]
- Iannucci, A.; Conti, J.; Curcio, F.; Iurino, D.A.; Mancini, M.; Mecozzi, B.; Strani, F.; Sardella, R. Middle Pleistocene mammal faunas of the area of Rome: Recent results and ongoing work on the MUST collection. J. Mediterr. Earth Sci. 2023, 15, 117–135. [Google Scholar]
- Mecozzi, B.; Iannucci, A.; Sardella, R.; Curci, A.; Daujeard, C.; Moncel, M.-H. Macaca ulna from new excavations at the Notarchirico Acheulean site (Middle Pleistocene, Venosa, southern Italy). J. Hum. Evol. 2021, 153, 102946. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.; Di Salvo, C.; Giallini, S.; Marini, M.; Simionato, M.; Caciolli, M.C.; Cavinato, G.P.; Moscatelli, M.M.; Polpetta, F.; Sirianni, P.; et al. The subsoil of the Colosseum and the detection of the ancient Tiber river Paleovalley (MIS 12-11) in Rome. J. Mediterr. Earth Sci. 2023, 15. [Google Scholar] [CrossRef]
- Milli, S.; Palombo, M.R. The high-resolution sequence stratigraphy and the mammal fossil record: A test in the Middle-Upper Pleistocene deposits of the Roman Basin (Latium, Italy). Quat. Int. 2005, 126/128, 251–270. [Google Scholar] [CrossRef]
- Bonadonna, P. Resti di Hippopotamus amphibius Linn. nei sedimenti del Pleistocene medio-inferiore della via Portuense, Roma. Boll. Soc. Geol. Ital. 1965, 84, 29–39. [Google Scholar]
- Petronio, C. Nuovi resti di Ippopotami del Pleistocene medio inferiore dei dintorni di Roma e problemi di tassonomia e filogenesi del gruppo. Geol. Rom. 1986, 25, 63–73. [Google Scholar]
- Ambrosetti, P. Cromerian fauna of the Rome area. Quaternaria 1967, 9, 267–283. [Google Scholar]
- Karner, D.B.; Marra, F. Correlation of fluviodeltaic aggradational sections with 1 glacial climate history: A revision of the classical Pleistocene stratigraphy of Rome. Geol. Soc. Am. Bull. 1998, 110, 748–758. [Google Scholar] [CrossRef]
- Blanc, A.C.; Lona, F.; Settepassi, F. Una torba ad Abies, malacofauna montana e criosedimenti nel Pleistocene inferiore di Roma—Il periodo glaciale Cassio. Ricerche sul Quaternario Laziale 1. Quaternaria 1955, 2, 151–158. [Google Scholar]
- Marra, F.; Nomade, S.; Pereira, A.; Petronio, C.; Salari, L.; Sottili, G.; Bahain, J.-J.; Boschian, G.; Di Stefano, G.; Falguéres, C.; et al. A review of the geologic sections and the faunal assemblages of Aurelian Mammal Age of Latium (Italy) in the light of a new chronostratigraphic framework. Quat. Sci. Rev. 2018, 181, 173–199. [Google Scholar] [CrossRef]
- Portis, A. Contribuzioni Alla Storia Fisica del Bacino di Roma e Studii Sopra L’estensione da Darsi al Pliocene Superiore; L. Roux e C: Roma, Italy, 1893; Volume I. [Google Scholar]
- Terrigi, G. Le formazioni vulcaniche del bacino Romano considerate nella loro fisica costituzione e giacitura. Att. Della Regia Accad. Dei Lincei 1881, 10, 389–419. [Google Scholar]
- Portis, A. Di alcuni avanzi fossili di grandi ruminanti principalmente della provincia di Roma. Palaeontologr. Ital. 1907, 13, 141–198. [Google Scholar]
- Marra, F.; Karner, D.B.; Freda, C.; Gaeta, M.; Renne, P.R. Large mafic eruptions at the Alban Hills Volcanic District (Central Italy): Chronostratigraphy, petrography and eruptive behavior. J. Volcanol. Geotherm. Res. 2009, 179, 217–232. [Google Scholar] [CrossRef]
- Caloi, L.; Palombo, M.R.; Petronio, C. La fauna quaternaria di Sedia del Diavolo (Roma). Quaternaria 1980, 22, 177–209. [Google Scholar]
- Palombo, M.R.; Milli, S.; Rosa, C. Remarks on the biochronology of the late Middle Pleistocene mammalian faunal complexes of the Campagna Romana (Latium, Italy). Geol. Rom. 2004, 37, 135–143. [Google Scholar]
- Fidalgo, D.; Madurell-Malapeira, J.; Martino, R.; Pandolfi, L.; Rosas, A. An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula. Quaternary 2024, 7, 4. [Google Scholar] [CrossRef]
- Klein, R.G.; Scott, K. Glacial/Interglacial Size Variation in Fossil Spotted Hyenas (Crocuta crocuta) from Britain. Quat. Res. 1989, 32, 88–95. [Google Scholar] [CrossRef]
- Cerilli, E.; Petronio, C. Biometrical variations of Bos primigenius Bojanus. In Proceedings of the International Symposium “Ongules/Ungulates 91”, Toulouse, France, 2–6 September 1991; pp. 37–42. [Google Scholar]
- Wright, E. The History of the European Aurochs (Bos primigenius) from the Middle Pleistocene to its Extinction: An Archaeological Investigation of Its Evolution, Morphological Variability and Response to Human Exploitation. Ph.D. Thesis, University of Sheffield, Sheffield, UK, 2013. [Google Scholar]
- Mecozzi, B.; Bartolini Lucenti, S. The Late Pleistocene Canis lupus (Canidae, Mammalia) from Avetrana (Apulia, Italy): Reappraisal and new insights on the European glacial wolves. Ital. J. Geosci. 2018, 137, 138–150. [Google Scholar] [CrossRef]
- Sansalone, G.; Bertè, D.F.; Maiorino, L.; Pandolfi, L. Evolutionary trends and stasis in carnassial teeth of European Pleistocene wolf Canis lupus (Mammalia, Canidae). Quat. Sci. Rev. 2015, 110, 36–48. [Google Scholar] [CrossRef]
- Iannucci, A.; Sardella, R.; Strani, F.; Mecozzi, B. Size shifts in late Middle Pleistocene to Early Holocene Sus scrofa (Suidae, Mammalia) from Apulia (southern Italy): Ecomorphological adaptations? Hystrix 2020, 31, 10–20. [Google Scholar]
- Bergmann, C. Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer Grösse. Güttinger Studien. Gott. Stud. 1847, 3, 595–708. [Google Scholar]
- Bellucci, L.; Mazzini, I.; Scardia, G.; Bruni, L.; Parenti, F.; Segre, A.G.; Segre Naldini, E.; Sardella, R. The site of Coste San Giacomo (Early Pleistocene, central Italy): Palaeoenvironmental analysis and biochronological overview. Quat. Int. 2012, 267, 30–39. [Google Scholar] [CrossRef]
- Palmqvist, P.; Gröcke, D.R.; Arribas, A.; Fariña, R.A. Paleoecological reconstruction of a lower Pleistocene large mammal community using biogeochemical (δ13C, δ15N, δ18O, Sr: Zn) and ecomorphological approaches. Paleobiology 2003, 29, 205–229. [Google Scholar] [CrossRef]
- Palmqvist, P.; Pérez-Claros, J.A.; Janis, C.M.; Figueirido, B.; Torregrosa, V.; Grocke, D.R. Biogeochemical and ecomorphological inferences on prey selection and resource partitioning among mammalian carnivores in an early Pleistocene community. Palaios 2008, 23, 724–737. [Google Scholar] [CrossRef]
- Palmqvist, P.; Rodríguez-Gómez, G.; Figueirido, B.; García-Aguilar, J.M.; Pérez-Claros, J.A. On the ecological scenario of the first hominin dispersal out of Africa. L’Anthropologie 2022, 126, 102998. [Google Scholar] [CrossRef]
- Shackleton, N.J. New data on the evolution of pliocene climatic variability. In Paleoclimate and Evolution, with Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partidge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA, 1995; pp. 242–248. [Google Scholar]








| Grooves on the external surface of canines | |||||
| Parallel | Convergent | ||||
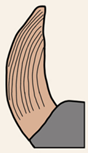 |  | ||||
| Sites | Species | References | Fossil H. antiquus | Fossil H. amphibius | Extant H. amphibius |
| Cava Santarelli | Hippopotamus antiquus | [45] | X | ||
| Colle Curti | Hippopotamus antiquus | [26,43] | X | ||
| Saticula | Hippopotamus antiquus | [48], this work | X | ||
| Cava Redicicoli | Hippopotamus cf. antiquus | [40], this work | X | ||
| Bussi | Hippopotamus antiquus | This work | 1 specimen | ||
| Ortona | Hippopotamus antiquus | [34,58] | X | ||
| Vallinfreda | Hippopotamus antiquus | This work | 1 specimen | ||
| Cava Arnolfi | Hippopotamus antiquus | This work | 1 specimen | ||
| Campo di Merlo | Hippopotamus antiquus | This work | 1 specimen | ||
| Cava di Breccia—Casal Selce 2 | Hippopotamus antiquus | This work | 1 specimen | ||
| Isernia La Pineta | Hippopotamus antiquus | This work | 3 specimens | ||
| Terranera | Hippopotamus antiquus | This work | 5 specimens | ||
| Cava Montanari | Hippopotamus amphibius | [22] | 1 specimen | ||
| Monte Antenne | Hippopotamus amphibius | This work | 2 specimens | ||
| Ponte Molle | Hippopotamus cf. amphibius | This work | 1 specimen | 3 specimens | |
| Tor di Quinto | Hippopotamus amphibius | This work | 3 specimens | ||
| Trentangioli | Hippopotamus amphibius | This work | 2 specimens | ||
| Batteria Nomentana | Hippopotamus amphibius | This work | 3 specimens | ||
| Sedia del Diavolo | Hippopotamus amphibius | This work | 6 specimens | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mecozzi, B.; Iannucci, A.; Arzarello, M.; Carpentieri, M.; Moncel, M.-H.; Peretto, C.; Sala, B.; Sardella, R. Middle Pleistocene Hippopotamuses from the Italian Peninsula: An Overview. Quaternary 2024, 7, 20. https://doi.org/10.3390/quat7020020
Mecozzi B, Iannucci A, Arzarello M, Carpentieri M, Moncel M-H, Peretto C, Sala B, Sardella R. Middle Pleistocene Hippopotamuses from the Italian Peninsula: An Overview. Quaternary. 2024; 7(2):20. https://doi.org/10.3390/quat7020020
Chicago/Turabian StyleMecozzi, Beniamino, Alessio Iannucci, Marta Arzarello, Marco Carpentieri, Marie-Hélène Moncel, Carlo Peretto, Benedetto Sala, and Raffaele Sardella. 2024. "Middle Pleistocene Hippopotamuses from the Italian Peninsula: An Overview" Quaternary 7, no. 2: 20. https://doi.org/10.3390/quat7020020
APA StyleMecozzi, B., Iannucci, A., Arzarello, M., Carpentieri, M., Moncel, M.-H., Peretto, C., Sala, B., & Sardella, R. (2024). Middle Pleistocene Hippopotamuses from the Italian Peninsula: An Overview. Quaternary, 7(2), 20. https://doi.org/10.3390/quat7020020