A Tale of Maize, Palm, and Pine: Changing Socio-Ecological Interactions from Pre-Classic Maya to the Present Day in Belize


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Modern Climate and Vegetation at Lamanai, Northern Belize
2.3. Site History in the Regional Context
2.4. Sediment Collection
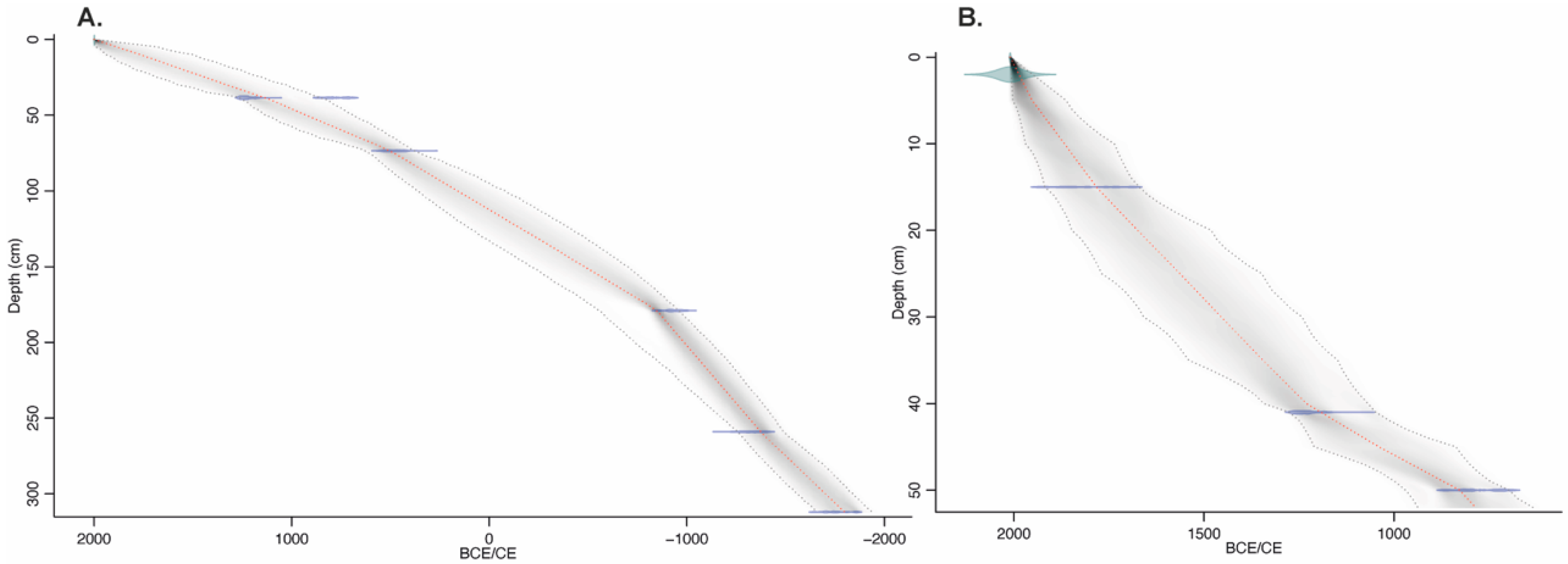
2.5. Chronology
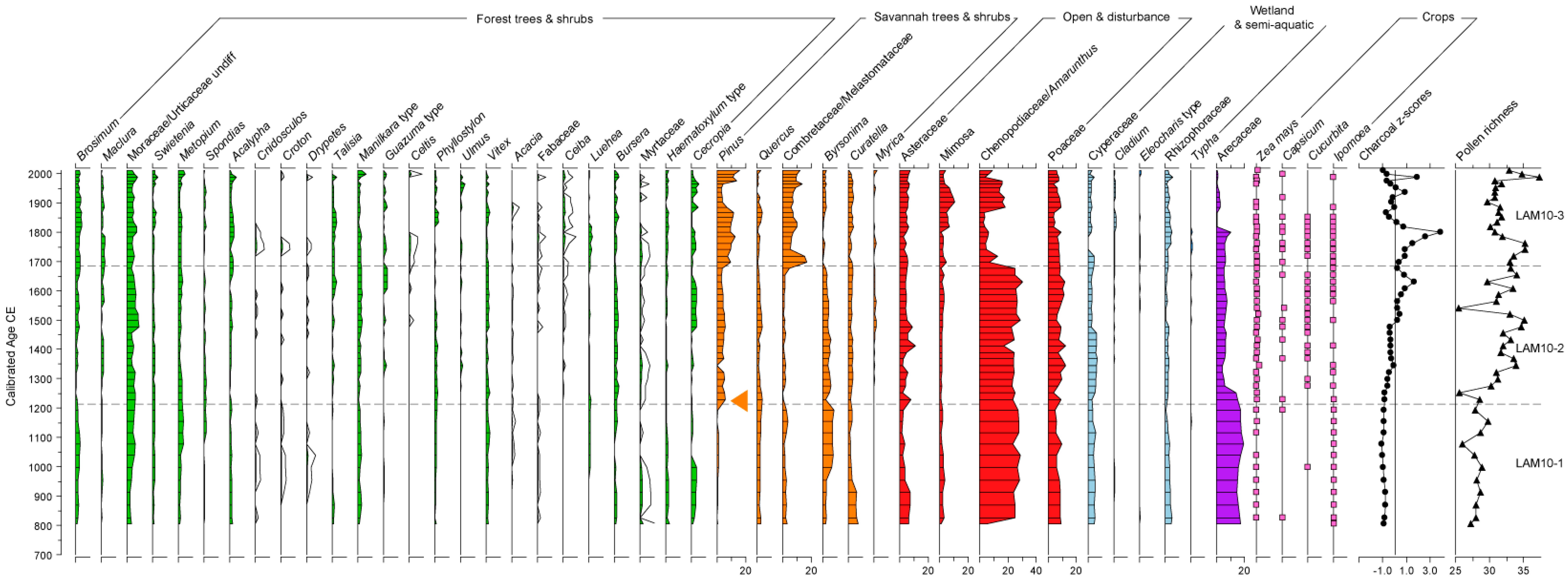
2.6. Pollen and Charcoal Analysis
2.7. Numerical Analyses
3. Results
3.1. Modern Pollen and Charcoal Assemblages
3.2. Long Core Record (Lamanai-99)
3.3. Short Core Record (Lamanai-10)
4. Discussion
4.1. Pollen and Charcoal Signals in Lamanai-99 and Lamanai-10
4.2. Maize and Palm—A Record of Maya Cultivation and Spanish Construction
4.3. Exploitation of Pine by the Maya, Spanish and British
4.4. The Impact of Land Management and Crop Production on Biodiversity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adolf, C.; Tovar, C.; Kühn, N.; Behling, H.; Berrío, J.C.; Dominguez-Vázquez, G.; Figueroa-Rangel, B.; Gonzalez-Carranza, Z.; Islebe, G.A.; Hooghiemstra, H.; et al. Identifying drivers of forest resilience in long-term records from the Neotropics. Biol. Lett. 2020, 16, 20200005. [Google Scholar] [CrossRef]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Loughlin, N.J.D.; Gosling, W.D.; Mothes, P.; Montoya, E. Ecological consequences of post-Columbian indigenous depopulation in the Andean–Amazonian corridor. Nat. Ecol. Evol. 2018, 2, 1233–1236. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Bailey, R.M.; Bhagwat, S.A.; Birks, H.J.B. Biodiversity baselines, thresholds and resilience: Testing predictions and assumptions using palaeoecological data. Trends Ecol. Evol. 2010, 25, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Montoya, E.; Lombardo, U.; Levis, C.; Aymard, G.A.; Mayle, F.E. Human Contribution to Amazonian Plant Diversity: Legacy of Pre-Columbian Land Use in Modern Plant Communities. In Neotropical Diversification: Patterns and Processes (Fascinating Life Sciences); Rull, V., Carnaval, A.C., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 495–520. [Google Scholar]
- De Oliveira, E.A.; Marimon-Junior, B.H.; Marimon, B.S.; Iriarte, J.; Morandi, P.S.; Maezumi, S.Y.; Nogueira, D.S.; Aragão, L.E.; da Silva, I.B.; Feldpausch, T.R. Legacy of Amazonian Dark Earth soils on forest structure and species composition. Glob. Ecol. Biogeogr. 2019. [Google Scholar] [CrossRef]
- White, D.A.; Hood, C.S. Vegetation patterns and environmental gradients in tropical dry forests of the northern Yucatan Peninsula. J. Veg. Sci. 2004, 15, 151–160. [Google Scholar] [CrossRef]
- Perring, M.P.; De Frenne, P.; Baeten, L.; Maes, S.L.; Depauw, L.; Blondeel, H.; Carón, M.M.; Verheyen, K. Global environmental change effects on ecosystems: The importance of land-use legacies. Glob. Chang. Biol. 2016, 22, 1361–1371. [Google Scholar] [CrossRef]
- Islebe, G.A.; Torrescano-Valle, N.; Aragón-Moreno, A.A.; Vela-Peláez, A.A.; Valdez-Hernández, M. The Paleoanthropocene of the Yucatán Peninsula: Palynological evidence of environmental change. Boletín Soc. Geol. Mex. 2018, 70, 49–60. [Google Scholar] [CrossRef]
- Haug, G.H.; Günther, D.; Peterson, L.C.; Sigman, D.M.; Hughen, K.A.; Aeschlimann, B. Climate and the collapse of Maya civilization. Science 2003, 299, 1731–1735. [Google Scholar] [CrossRef]
- Hodell, D.A.; Curtis, J.H.; Brenner, M. Possible role of climate in the collapse of Classic Maya civilization. Nature 1995, 375, 391–394. [Google Scholar] [CrossRef]
- Kennett, D.J.; Breitenbach, S.F.M.; Aquino, V.V.; Asmerom, Y.; Awe, J.; Baldini, J.U.L.; Bartlein, P.; Culleton, B.J.; Ebert, C.; Jazwa, C.; et al. Development and Disintegration of Maya Political Systems in Response to Climate Change. Science 2012, 338, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Kennett, D.J.; Beach, T.P. Archeological and environmental lessons for the Anthropocene from the Classic Maya collapse. Anthropocene 2013, 4, 88–100. [Google Scholar] [CrossRef]
- Franco-Gaviria, F.; Correa-Metrio, A.; Cordero-Oviedo, C.; López-Pérez, M.; Cárdenes-Sandí, G.M.; Romero, F.M. Effects of late Holocene climate variability and anthropogenic stressors on the vegetation of the Maya highlands. Quat. Sci. Rev. 2018, 189, 76–90. [Google Scholar] [CrossRef]
- Hansen, R.D.; Bozarth, S.; Jacob, J.; Wahl, D.; Schreiner, T. Climatic and environmental variability in the rise of Maya civilization: A preliminary perspective from northern Petén. Anc. Mesoam. 2002, 13, 273–295. [Google Scholar] [CrossRef]
- Leyden, B.W. Pollen evidence for climatic variability and cultural disturbance in the Maya Lowlands. Anc. Mesoam. 2002, 13, 85–101. [Google Scholar] [CrossRef]
- Wahl, D.; Byrne, R.; Schreiner, T.; Hansen, R. Holocene vegetation change in the northern Peten and its implications for Maya prehistory. Quat. Res. 2006, 65, 380–389. [Google Scholar] [CrossRef]
- McNeil, C.L.; Burney, D.A.; Burney, L.P. Evidence disputing deforestation as the cause for the collapse of the ancient Maya polity of Copan, Honduras. Proc. Natl. Acad. Sci. USA 2010, 107, 1017–1022. [Google Scholar] [CrossRef]
- Rushton, E.A.; Metcalfe, S.E.; Whitney, B.S. A late-Holocene vegetation history from the Maya lowlands, Lamanai, Northern Belize. Holocene 2013, 23, 485–493. [Google Scholar] [CrossRef]
- Ford, A.; Nigh, R. Origins of the Maya Forest Garden: Maya Resource Management. J. Ethnobiol. 2009, 29, 213–236. [Google Scholar] [CrossRef]
- Ford, A.; Nigh, R. The Maya Forest Garden: Eight Millennia of Sustainable Cultivation of the Tropical Woodlands; Routledge: Abingdon, UK, 2016. [Google Scholar]
- Ross, N.J. Modern tree species composition reflects ancient Maya “forest gardens” in northwest Belize. Ecol. Appl. 2011, 21, 75–84. [Google Scholar] [CrossRef]
- Graham, E. Collapse, conquest and Maya survival at Lamanai, Belize. Archaeol. Int. 2000, 4, 52–56. [Google Scholar] [CrossRef]
- Graham, E.A. Lamanai Reloaded: Alive and well in the early post-Classic. In Archaeological Investigations in the Eastern Maya Lowlands: Papers of the 2003 Belize Archaeology Symposium; Institute of Archaeology National Institute of Culture and History: Belmopan, Belize, 2004; Volume 1, p. 223. [Google Scholar]
- Rushton, E.A. “Under the Shade I Flourish”: An Environmental History of Northern Belize over the Last Three Thousand Five Hundred Years. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2014. [Google Scholar]
- Metcalfe, S.; Breen, A.; Murray, M.; Furley, P.; Fallick, A.; McKenzie, A. Environmental change in northern Belize since the latest Pleistocene. J. Quat. Sci. 2009, 24, 627–641. [Google Scholar] [CrossRef]
- Bridgewater, S. A Natural History of Belize: Insights from the Chiquibul Forest and Las Cuevas Research Station, 1st ed.; University of Texas Press: Austin, TX, USA, 2012. [Google Scholar]
- Setzerkorn, W.D. Formerly British Honduras: A Profile of The New Nation of Belize; Ohio University Press: Athens, OH, USA, 1981. [Google Scholar]
- Esselman, P.; Boles, E. Status and future needs of limnological research in Belize. Limnol. Dev. Ctries. 2001, 3, 35–68. [Google Scholar]
- Walker, S.H. Summary of Climatic Records for Belize; Land Resources Division, Overseas Development Administration: Surrey, UK, 1973. [Google Scholar]
- Meerman, J.; Sabido, W. Central American Ecosystems Map: Belize. In Ecosystem Map and Descriptions; The World Bank: Washington, DC, USA, 2001; Volume II. [Google Scholar]
- Beach, T.; Luzzadder-Beach, S.; Dunning, N.; Jones, J.; Lohse, J.; Guderjan, T.; Bozarth, S.; Millspaugh, S.; Bhattacharya, T. A review of human and natural changes in Maya Lowland wetlands over the Holocene. Quat. Sci. Rev. 2009, 28, 1710–1724. [Google Scholar] [CrossRef]
- Hansen, B.; Pohl, M.D. Pollen stratigraphy of Laguna de Cocos. In Ancient Maya Wetland Agriculture: Excavations on Albion Island, Northern Belize; Pohl, M., Ed.; Westview Press: Boulder, CO, USA, 1990; pp. 155–186. [Google Scholar]
- Jones, J.G. Pollen evidence for early settlement and agriculture in northern Belize. Palynology 1994, 18, 205–211. [Google Scholar] [CrossRef]
- Luzzadder-Beach, S.; Beach, T.P.; Dunning, N.P. Wetland fields as mirrors of drought and the Maya abandonment. Proc. Natl. Acad. Sci. USA 2012, 109, 3646–3651. [Google Scholar] [CrossRef]
- Morse, M.L. Pollen from Laguna Verde, Blue Creek, Belize: Implications for Paleoecology, Paleoethnobotany, Agriculture, and Human Settlement. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2009. [Google Scholar]
- Pohl, M.D.; Pope, K.O.; Jones, J.G.; Jacob, J.S.; Piperno, D.R.; DeFrance, S.D.; Lentz, D.L.; Gifford, J.A.; Danforth, M.E.; Josserand, J.K. Early Agriculture in the Maya Lowlands. Lat. Am. Antiq. 1996, 7, 355–372. [Google Scholar] [CrossRef]
- Adams, R.E.; Robichaux, H.R.; Valdez, F., Jr.; Houk, B.A.; Mathews, R. Transformations, periodicity, and urban development in the Three Rivers region. In The Terminal Classic in the Maya Lowlands: Collapse, Transition, and Transformation; Demarest, A.A., Rice, P.M., Rice, D.S., Eds.; University Press of Colorado: Boulder, CO, USA, 2004; pp. 324–341. [Google Scholar]
- Anselmetti, F.S.; Hodell, D.A.; Ariztegui, D.; Brenner, M.; Rosenmeier, M.F. Quantification of soil erosion rates related to ancient Maya deforestation. Geology 2007, 35, 915–918. [Google Scholar] [CrossRef]
- Beach, T.; Luzzadder-Beach, S.; Dunning, N.; Cook, D. Human and natural impacts on fluvial and karst depressions of the Maya Lowlands. Geomorphology 2008, 101, 308–331. [Google Scholar] [CrossRef]
- Deevey, E.S.; Rice, D.S.; Rice, P.M.; Vaughan, H.H.; Brenner, M.; Flannery, M.S. Mayan Urbanism: Impact on a Tropical Karst Environment. Science 1979, 206, 298–306. [Google Scholar] [CrossRef]
- Rice, D.S. Paleolimnological analysis in the central Peten, Guatemala. In The Managed Mosaic: Ancient Maya Agricultural and Resource Use; Fedick, S.L., Ed.; University of Utah Press: Salt Lake City, UT, USA, 1996; pp. 193–206. [Google Scholar]
- Rosenmeier, M.F.; Hodell, D.A.; Brenner, M.; Curtis, J.H.; Guilderson, T.P. A 4000-Year Lacustrine Record of Environmental Change in the Southern Maya Lowlands, Petén, Guatemala. Quat. Res. 2002, 57, 183–190. [Google Scholar] [CrossRef]
- Berry, K.A.; McAnany, P.A. Reckoning with the wetlands and their role in ancient Maya society. In The Political Economy of Ancient Mesoamerica: Transformations during the Formative and Classic Periods; Scarborough, V.L., Clark, J.E., Eds.; University of New Mexico Press: Albuquerque, NM, USA, 2007; pp. 149–162. [Google Scholar]
- Pendergast, D.M. The church in the jungle: The ROM’s first season at Lamanai. Rotunda 1975, 8, 32–40. [Google Scholar]
- Pendergast, D.M. Lamanai, Belize: Summary of excavation results, 1974–1980. J. Field Archaeol. 1981, 8, 29–53. [Google Scholar]
- Pendergast, D.M. The 19th-century sugar mill at Indian Church, Belize. J. Field Archaeol. 1982, 8, 1–5. [Google Scholar]
- Pendergast, D.M. Ancient Maya mercury. Science 1982, 217, 533–535. [Google Scholar] [CrossRef]
- Pendergast, D.M. Intercessions with the Gods: Caches and their significance at Altun Ha and Lamanai, Belize. In The Sowing and the Dawning: Termination, Dedication, and Transformation in the Archaeological and Ethnographic Record of Mesoamerica; Boteler-Mock, S., Ed.; University of New Mexico Press: Albuquerque, NM, USA, 2002; pp. 55–63. [Google Scholar]
- Pendergast, D.M.; Sabloff, J.; Andrews, E.W. Stability through Change: Lamanai, Belize from the Ninth to the Seventeenth Century. In Late Lowland Maya Civilization: Classic to Postclassic; Sabloff, A., Andrews, E.W., Eds.; University of New Mexico Press: Albuquerque, NM, USA, 1986; pp. 223–250. [Google Scholar]
- Graham, E. Maya Christians and Their Churches in Sixteenth-Century Belize; University Press of Florida: Gainesville, FL, USA, 2011. [Google Scholar]
- Pendergast, D.M. The southern Maya lowlands contact experience: The view from Lamanai, Belize. In Columbian Consequences: The Spanish Borderlands in Pan-American Perspective; Thomas, D.E., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1991; pp. 336–354. [Google Scholar]
- Pendergast, D.M.; Jones, G.D.; Graham, E. Locating Maya lowlands Spanish colonial towns: A case study from Belize. Lat. Am. Antiq. 1993, 4, 59–73. [Google Scholar] [CrossRef]
- Scarborough, V.L.; Gallopin, G.G. A water storage adaptation in the Maya lowlands. Science 1991, 251, 658–662. [Google Scholar] [CrossRef]
- Jones, G.D. Maya Resistance to Spanish Rule: Time and History on a Colonial Frontier; University of New Mexico Press: Albuquerque, NM, USA, 1989. [Google Scholar]
- Mayfield, T. Ceramics, Landscape, and Colonialism: Archaeological Analysis of the British Settlement at Lamanai, Belize. Master’s Thesis, Illinois State University, Normal, IL, USA, 2009. [Google Scholar]
- Reimer, P.J.; William, E.N.; Austin, E.B.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J. rbacon: Age-Depth Modelling Using Bayesian Statistics; R Package Version 2; R Package: R Development Core Team: Vienna, Austria, 2018. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bennett, K.D.; Willis, K.J. Pollen. In Tracking Environmental Change Using Lake Sediments: Vol 3. Terrestrial, Algal, and Siliceous Indicators; Smol, J.P., Birks, J.B., Last, W.M., Eds.; Springer: Berlin, Germany, 2002; pp. 5–32. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; John Wiley: Chichester, UK, 1989. [Google Scholar]
- Whitney, B.S.; Rushton, E.A.; Carson, J.F.; Iriarte, J.; Mayle, F.E. An improved methodology for the recovery of Zea mays and other large crop pollen, with implications for environmental archaeology in the Neotropics. Holocene 2012, 22, 1087–1096. [Google Scholar] [CrossRef]
- Holst, I.; Moreno, J.E.; Piperno, D.R. Identification of teosinte, maize, and Tripsacum in Mesoamerica by using pollen, starch grains, and phytoliths. Proc. Natl. Acad. Sci. USA 2007, 104, 17608–17613. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [PubMed]
- Van Heerwaarden, J.; Doebley, J.; Briggs, W.H.; Glaubitz, J.C.; Goodman, M.M.; Gonzalez, J.D.; Ross-Ibarra, J. Genetic signals of origin, spread, and introgression in a large sample of maize landraces. Proc. Natl. Acad. Sci. USA 2011, 108, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Juggins, S. Rioja: Analysis of Quaternary Science Data, R package version (0.9-21). 2017. Available online: http://cran.r-project.org/package=rioja (accessed on 12 October 2020).
- Bennett, K. Psimpoll and Pscomb. Software, Queen’s University Belfast. 2008. Available online: http://chrono.qub.ac.uk/psimpoll/psimpoll.html (accessed on 12 October 2020).
- Birks, H.J.B.; Line, J. The use of rarefaction analysis for estimating palynological richness from Quaternary pollen-analytical data. Holocene 1992, 2, 1–10. [Google Scholar] [CrossRef]
- Bhattacharya, T.; Beach, T.; Wahl, D. An analysis of modern pollen rain from the Maya lowlands of northern Belize. Rev. Palaeobot. Palynol. 2011, 164, 109–120. [Google Scholar] [CrossRef]
- Sugita, S. Pollen Representation of Vegetation in Quaternary Sediments: Theory and Method in Patchy Vegetation. J. Ecol. 1994, 82, 881–897. [Google Scholar] [CrossRef]
- Jarosz, N.; Loubet, B.; Durand, B.; McCartney, A.; Foueillassar, X.; Huber, L. Field measurements of airborne concentration and deposition rate of maize pollen. Agric. For. Meteorol. 2003, 119, 37–51. [Google Scholar] [CrossRef]
- Raynor, G.S.; Ogden, E.C.; Hayes, J.V. Dispersion and Deposition of Corn Pollen from Experimental Sources 1. Agron. J. 1972, 64, 420–427. [Google Scholar] [CrossRef]
- Twiddle, C.L.; Bunting, M.J. Experimental investigations into the preservation of pollen grains: A pilot study of four pollen types. Rev. Palaeobot. Palynol. 2010, 162, 621–630. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a fire proxy. In Tracking Environmental Change Using Lake Sediments: Vol 3. Terrestrial, Algal, and Siliceous Indicators; Smol, J.P., Birks, J.B., Last, W.M., Eds.; Springer: Berlin, Germany, 2002; pp. 75–97. [Google Scholar]
- Krause, S.; Beach, T.; Luzzadder-Beach, S.; Guderjan, T.H.; Valdez, F.; Eshleman, S.; Doyle, C.; Bozarth, S.R. Ancient Maya wetland management in two watersheds in Belize: Soils, water, and paleoenvironmental change. Quat. Int. 2019, 502, 280–295. [Google Scholar] [CrossRef]
- La Torre, M.D.L.A.; Islebe, G.A. Others Traditional ecological knowledge and use of vegetation in southeastern Mexico: A case study from Solferino, Quintana Roo. Biodivers. Conserv. 2003, 12, 2455–2476. [Google Scholar]
- Schlesinger, V. Animals and Plants of the Ancient Maya: A Guide; University of Texas Press: Austin, TX, USA, 2001. [Google Scholar]
- Pendergast, D.M. Lamanai, Belize: An updated view. In Lowland Maya Postclassic; University of Texas Press: Austin, TX, USA, 1985; pp. 91–103. [Google Scholar]
- Rue, D.J. Early agriculture and early Postclassic Maya occupation in western Honduras. Nature 1987, 326, 285–286. [Google Scholar] [CrossRef]
- Lentz, D.L.; Graham, E.; Vinaja, X.; Slotten, V.; Jain, R. Agroforestry and ritual at the ancient Maya center of Lamanai. J. Archaeol. Sci: Rep. 2016, 8, 284–294. [Google Scholar] [CrossRef][Green Version]
- Morehart, C.T.; Lentz, D.L.; Prufer, K.M. Wood of the gods: The ritual use of pine (Pinus spp.) by the ancient lowland Maya. Lat. Am. Antiq. 2005, 16, 255–274. [Google Scholar] [CrossRef]
- Brady, J.E. An Investigation of Maya Ritual Cave Use with Special Reference to Naj Tunich, Guatemala. Ph.D. Thesis, Department of Anthropology, University of California, Los Angeles, CA, USA, 1989. [Google Scholar]
- Graham, E.; MeNatt, L.; Gutehen, M.A. Excavations in Footprint Cave, Caves Branch, Belize. J. Field Archaeol. 1980, 7, 153–172. [Google Scholar]
- Reents-Budet, D.; MacLeod, B. The Archaeology of Petroglyph Cave, Cayo District, Belize; Royal Ontario Museum: Toronto, ON, Canada, 1997. [Google Scholar]
- Stone, A.J. Images from the Underworld: Naj Tunich and the Tradition of Maya Cave Painting; University of Texas Press: Austin, TX, USA, 2010. [Google Scholar]
- Martin, S.; Grube, N. Chronicle of the Maya Kings and Queens: Deciphering the Dynasties of the Ancient Maya; Thames and Hudson: London, UK, 2008. [Google Scholar]
- Schele, L.; Miller, J.H. The Mirror, the Rabbit, and the Bundle: “Accession” Expressions from the Classic Maya Inscriptions. In Studies in Pre-Columbian Art and Archaeology No. 25; Dumbarton Oaks: Washington, DC, USA, 1983; pp. 1–99. [Google Scholar]
- Stuart, D. “The fire enters his house” Architecture and ritual in Classic Maya texts. In Function and Meaning in Classic Maya Architecture; Houston, S.D., Ed.; Dumbarton Oaks: Washington, DC, USA, 1998; pp. 373–425. [Google Scholar]
- Curtis, J.H.; Hodell, D.A.; Brenner, M. Climate variability on the Yucatan Peninsula (Mexico) during the past 3500 years, and implications for Maya cultural evolution. Quat. Res. 1996, 46, 37–47. [Google Scholar] [CrossRef]
- Hodell, D.A.; Brenner, M.; Curtis, J.H. Terminal Classic drought in the northern Maya lowlands inferred from multiple sediment cores in Lake Chichancanab (Mexico). Quat. Sci. Rev. 2005, 24, 1413–1427. [Google Scholar] [CrossRef]
- Hodell, D.A.; Brenner, M.; Curtis, J.H. Climate and cultural history of the northeastern Yucatan Peninsula, Quintana Roo, Mexico. Clim. Chang. 2007, 83, 215–240. [Google Scholar] [CrossRef]
- Neff, H.; Pearsall, D.M.; Jones, J.G.; de Pieters, B.A.; Freidel, D.E. Climate change and population history in the Pacific lowlands of southern Mesoamerica. Quat. Res. 2006, 65, 390–400. [Google Scholar] [CrossRef]
- Mayfield, T.; Graham, E.; Pendergast, D. Cane and Consumerism. In Technology and Tradition in Mesoamerica after the Spanish Invasion: Archaeological Perspectives; Alexander, R.T., Ed.; University of New Mexico Press: Albuquerque, NM, USA, 2019; pp. 147–165. [Google Scholar]
- Morris, D. The Colony of British Honduras: Its Resources and Prospects; with Particular Reference to Its Indigenous Plants and Economic Productions; E. Stanford: London, UK, 1883. [Google Scholar]
- Chazdon, R.L.; Peres, C.A.; Dent, D.; Sheil, D.; Lugo, A.E.; Lamb, D.; Stork, N.E.; Miller, S.E. The potential for species conservation in tropical secondary forests. Conserv. Biol. 2009, 23, 1406–1417. [Google Scholar] [CrossRef]
- Akers, P.D.; Brook, G.A.; Railsback, L.B.; Cherkinksy, A.; Liang, F.; Ebert, C.E.; Hoggarth, J.A.; Awe, J.J.; Cheng, H.; Edwards, R.L. Integrating U-Th, 14C, and 210Pb methods to produce a chronologically reliable isotope record for the Belize River Valley Maya from a low-uranium stalagmite. Holocene 2019, 29, 1234–1248. [Google Scholar] [CrossRef]
- Ford, A. Dominant plants of the Maya forest and gardens of El Pilar: Implications for paleoenvironmental reconstructions. J. Ethnobiol. 2008, 28, 179–199. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Event |
|---|---|
| 1500 BCE | Archaeological, palaeolimnological and palaeoecological records indicate settlement at Lamanai. |
| 600 BCE | Period of substantial settlement at Lamanai. |
| 100 BCE | Archaeological record indicates that Main Temple (Str. N10-43) reaches 33 m in height. |
| 100–400 CE | Early Classic building activity across the site. |
| 400–600 CE | Construction activity continues during the Middle Classic, including at the Mask Temple (Str. N9-56). |
| 600–800 CE | Throughout the Late Classic, building continues around the main plaza, including the carving of stela. |
| 800–1000 CE | Regional disintegration of Maya sites at La Milpa and Altun Ha whilst at Lamanai continues to thrive during the Terminal Classic, with construction of masonry platforms and a ball court. |
| 1000–1200 CE | Early Postclassic period appears to have a reduction in population at Lamanai and an increased focus of activity at the lagoon-side. |
| 1200–1500 CE | Continued occupation of the site, with distinctive pottery and evidence of effigy censers. |
| 1500–1544 CE | Terminal Postclassic/Early Colonial Period, likely last pre-Colonial ruler buried ca. 1525 CE. |
| 1544–1800 CE | Spanish Colonial Period. Two stone churches built at Lamanai (YDL 1 and 2). |
| 1800–1964 CE | 1862 CE Belize is designated a British Crown Colony, with a British Governor. Widespread archaeological evidence of British ceramic imports at Lamanai and British sugar mill constructed at Lamanai, in operation until 1875 CE. |
| 1964–1981 CE | 1964 CE Belize, self-governing British Colony. Belize became an independent nation in 1981 CE. |
| Lab ID | Core | Depth | Material | 14C yr BP | Corrected 14C age |
| Lamanai-10 dates; age-model based on five dates from the core and one using a correlated pollen tie point | |||||
| SUERC-43153 | 2010 | 2 cm | Terrestrial macros | Modern | ---- |
| SUERC-43156 | 2010 | 15 cm | Terrestrial macros | 143 ± 36 | ---- |
| ** SUERC-43157 | 2010 | 30 cm | Terrestrial macros | Modern | ---- |
| ** SUERC-45706 | 2010 | 40 cm | Terrestrial macros | 176 ± 37 | ---- |
| * AA-35787 | 1999 | 41 cm (=39.5 cm in Lamanai-99) | Terrestrial macros | 810 ± 40 | ---- |
| BETA-309657 | 2010 | 50 cm | Terrestrial macros | 1240 ± 30 | ---- |
| Lamanai-99 dates; age-model based on three macrofossil dates and two carbonate dates corrected with a 1660-yr offset | |||||
| † AA-35787 | 1999 | 38.5 cm | Organic Material | 810 ± 40 | ---- |
| † CAMS-77196 | 1999 | 38.5 cm | Gastropods | 2470 ± 40 | ---- |
| SUERC-4104 | 1999 | 72–75 cm | Gastropods | 3260 ± 50 | 1600 ± 50 |
| SUERC-4108 | 1999 | 178–180 cm | Gastropods | 4445 ± 35 | 2785 ± 35 |
| AA-35786 | 1999 | 259 cm | Organic Material | 3070 ± 50 | ---- |
| CAMS-77195 | 1999 | 312 cm | Organic Material | 3440 ± 40 | ---- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rushton, E.A.C.; Whitney, B.S.; Metcalfe, S.E. A Tale of Maize, Palm, and Pine: Changing Socio-Ecological Interactions from Pre-Classic Maya to the Present Day in Belize. Quaternary 2020, 3, 30. https://doi.org/10.3390/quat3040030
Rushton EAC, Whitney BS, Metcalfe SE. A Tale of Maize, Palm, and Pine: Changing Socio-Ecological Interactions from Pre-Classic Maya to the Present Day in Belize. Quaternary. 2020; 3(4):30. https://doi.org/10.3390/quat3040030
Chicago/Turabian StyleRushton, Elizabeth A. C., Bronwen S. Whitney, and Sarah E. Metcalfe. 2020. "A Tale of Maize, Palm, and Pine: Changing Socio-Ecological Interactions from Pre-Classic Maya to the Present Day in Belize" Quaternary 3, no. 4: 30. https://doi.org/10.3390/quat3040030
APA StyleRushton, E. A. C., Whitney, B. S., & Metcalfe, S. E. (2020). A Tale of Maize, Palm, and Pine: Changing Socio-Ecological Interactions from Pre-Classic Maya to the Present Day in Belize. Quaternary, 3(4), 30. https://doi.org/10.3390/quat3040030