
A Description of Two New Species of the Genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with Comments on Hominin and South Asian Ruminants Dispersals


Abstract
1. Introduction
2. Materials and Methods
3. Systematic Paleontology
3.1. Taxonomic Context
- Family: Cervidae Goldfuss, 1820.
- Subfamily: Cervinae Goldfuss, 1820.
- Genus: Rucervus Hodgson, 1838.
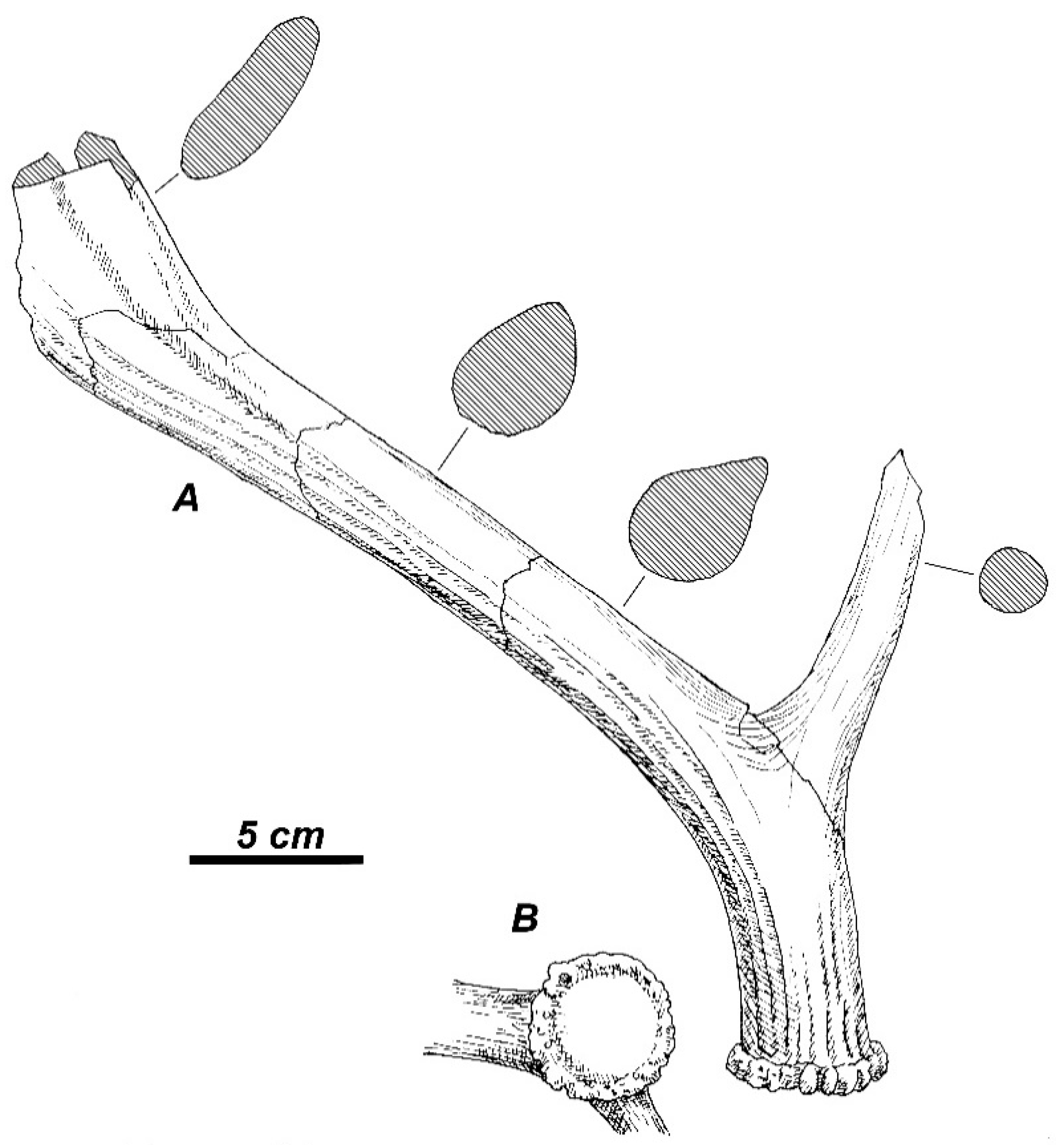
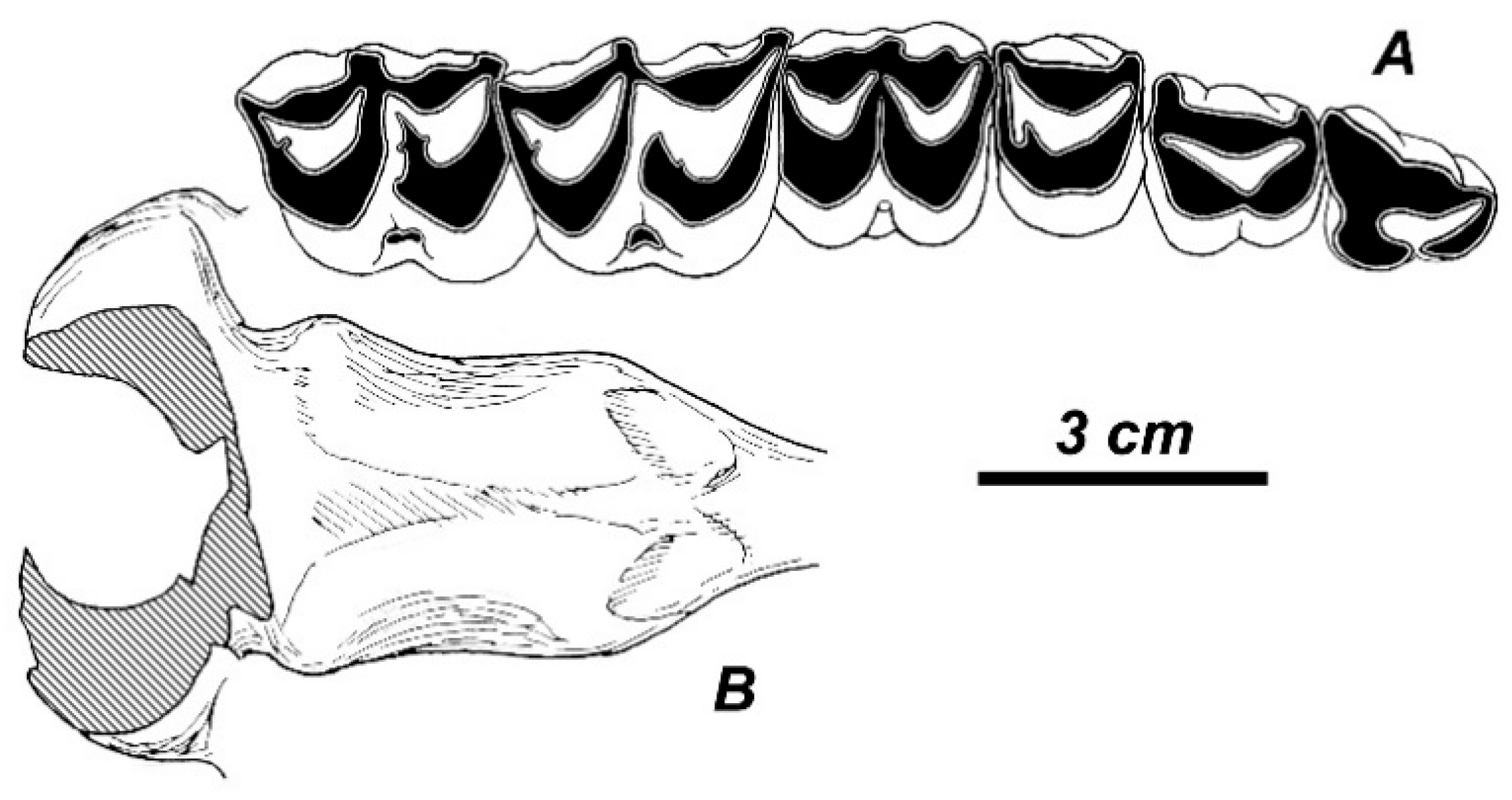
3.2. The New Deer Species from Valea Grăunceanului
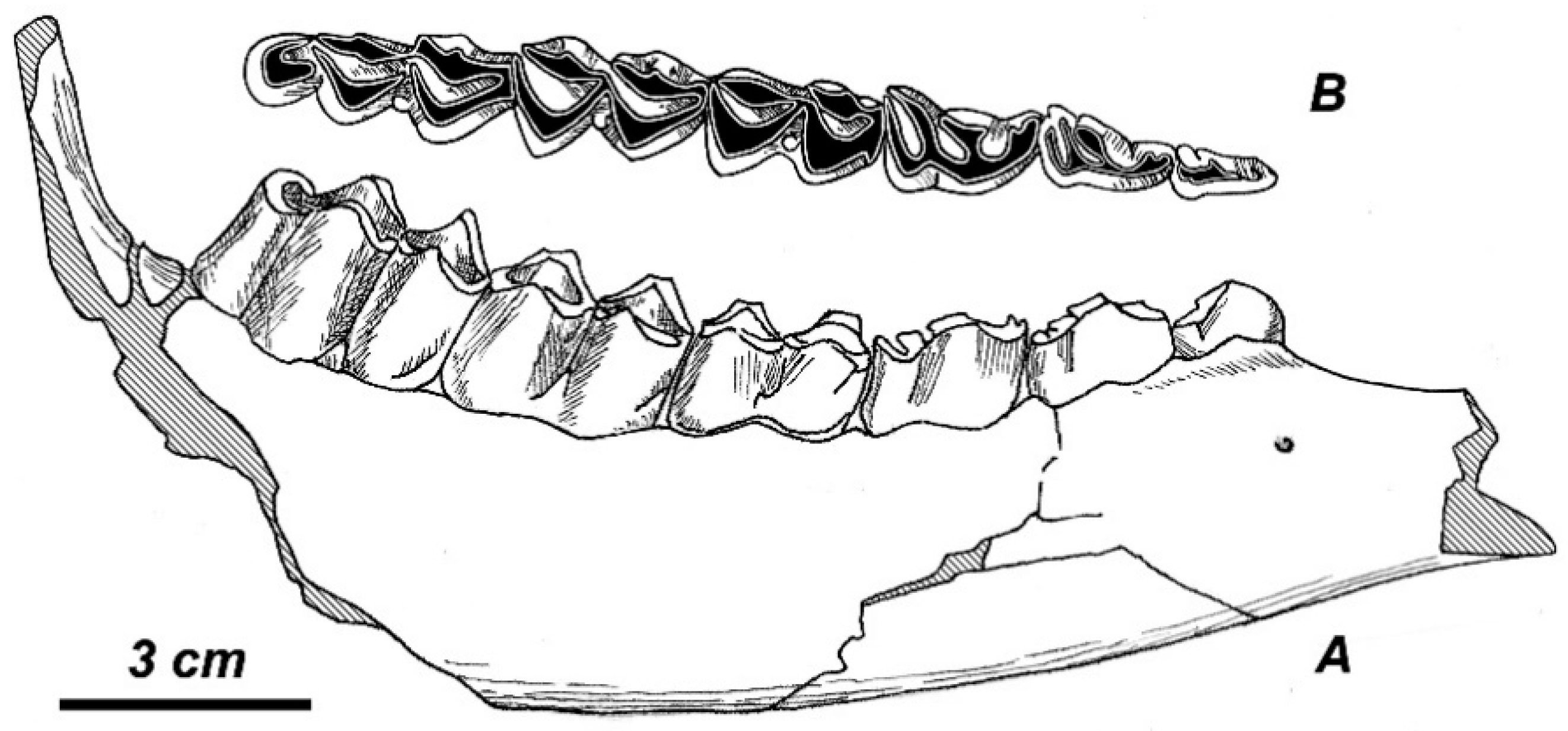
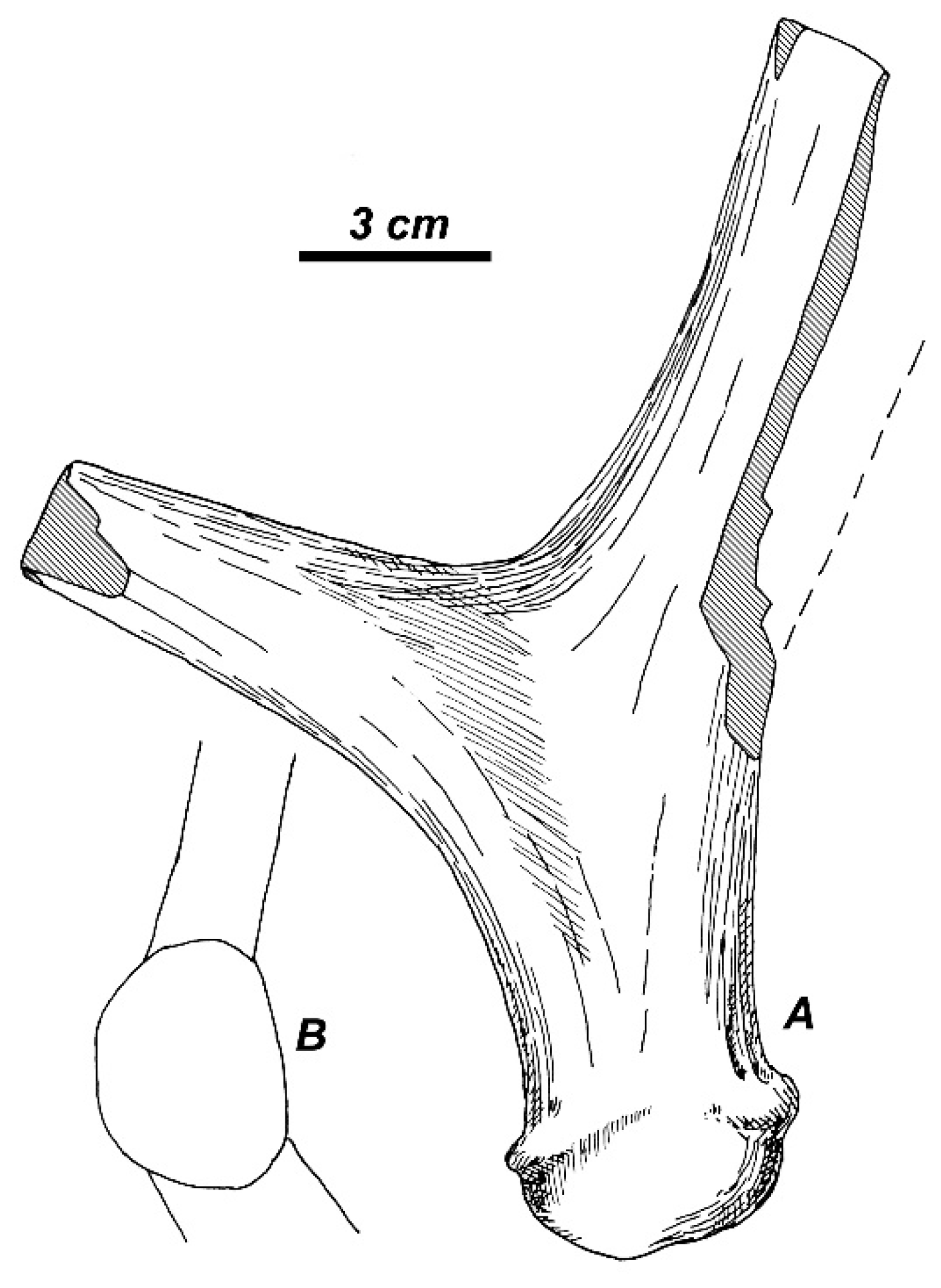
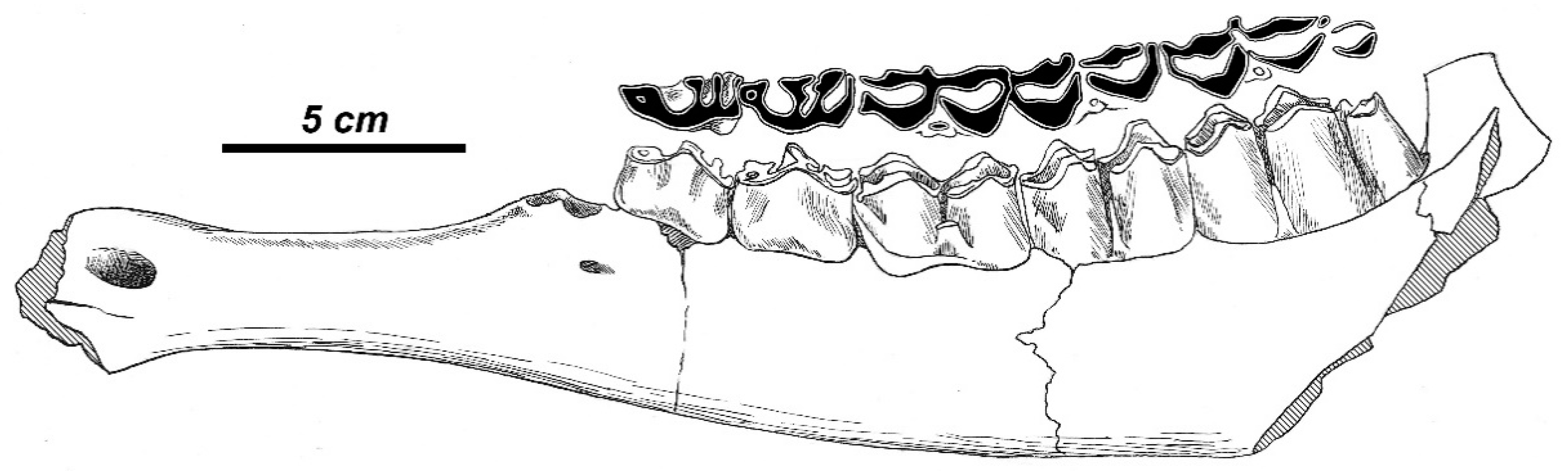
3.2.1. Taxonomic Description
3.2.2. Additional Material from the Type Locality
3.2.3. Other Findings of Rucervus radulescui
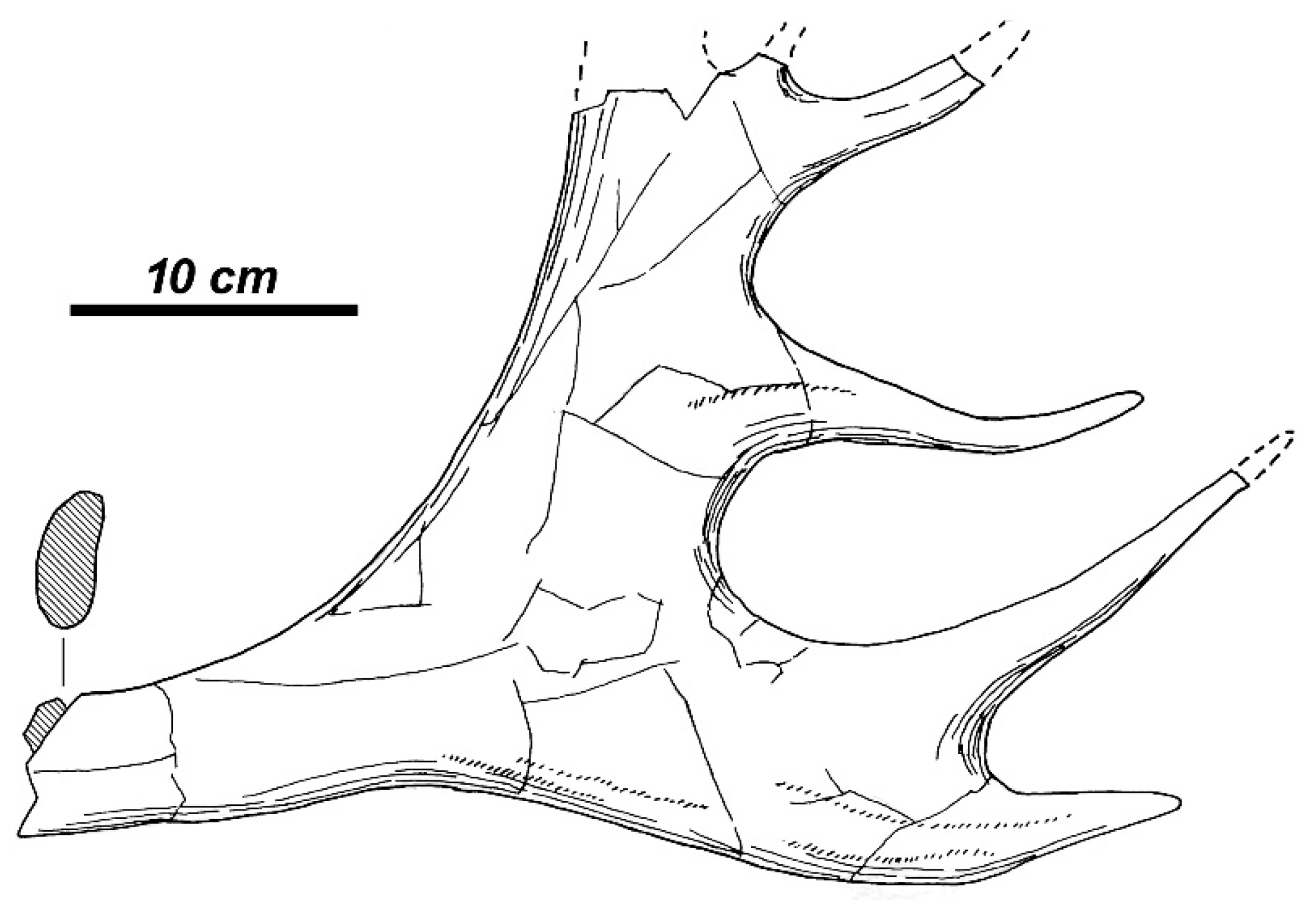
3.3. The New Large-Sized Deer Species from Apollonia-1
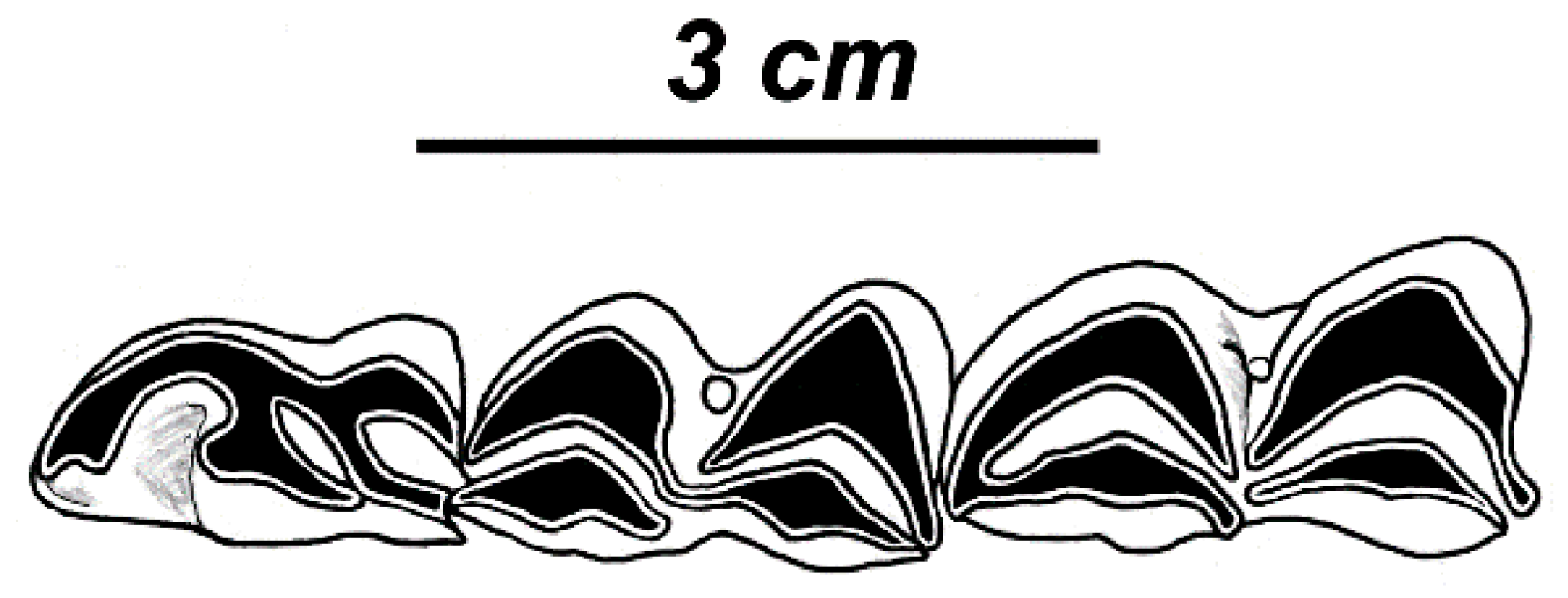
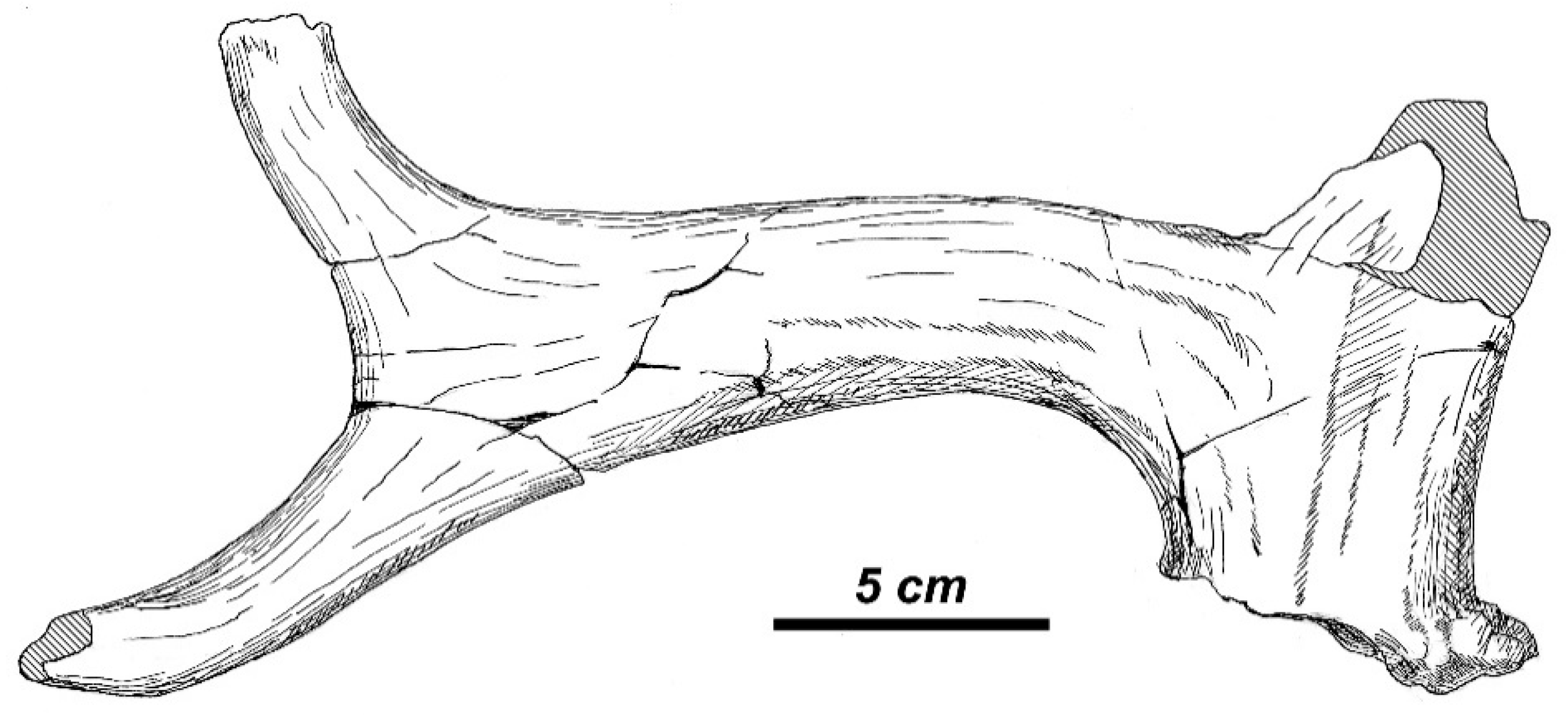
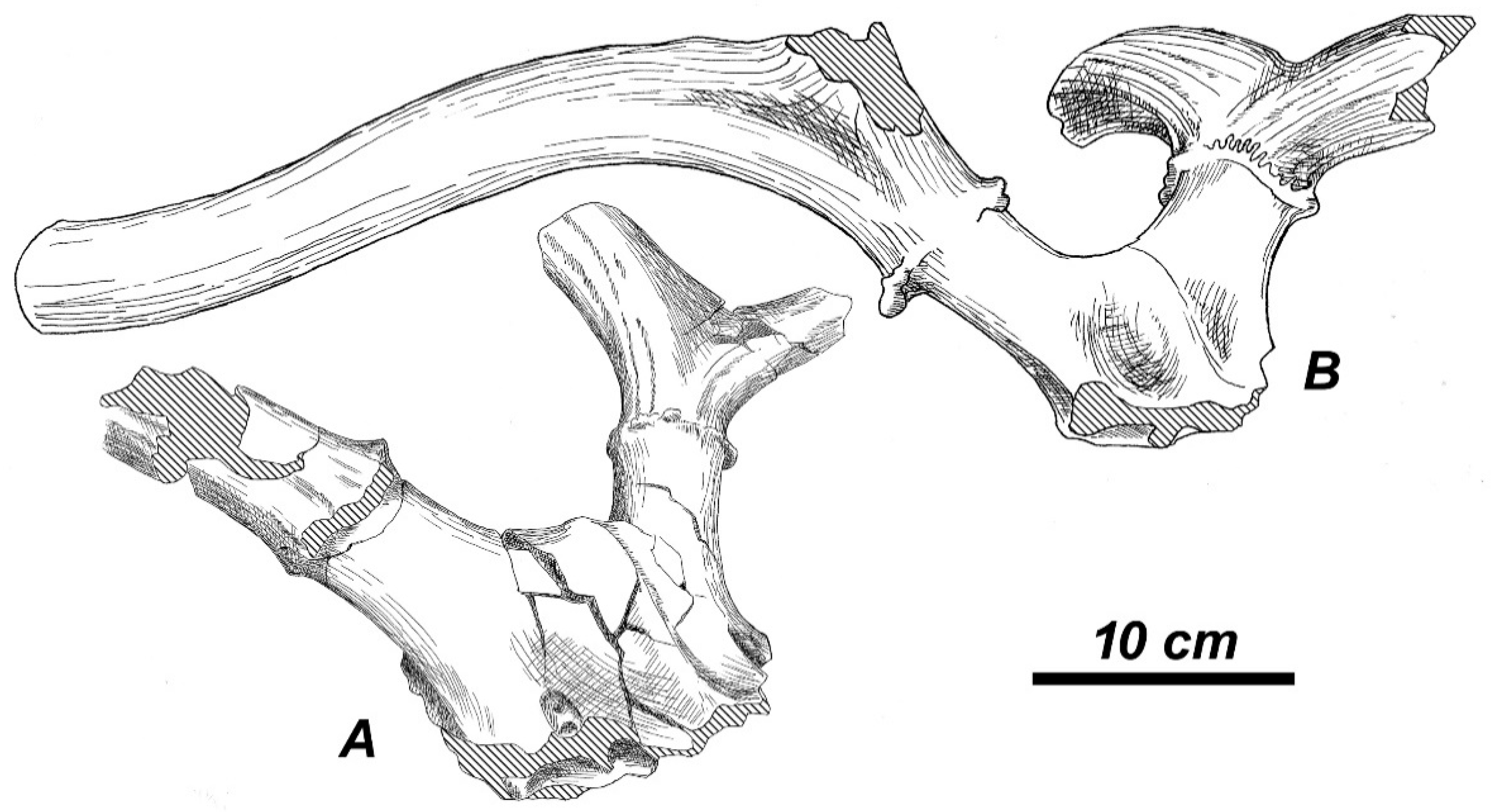
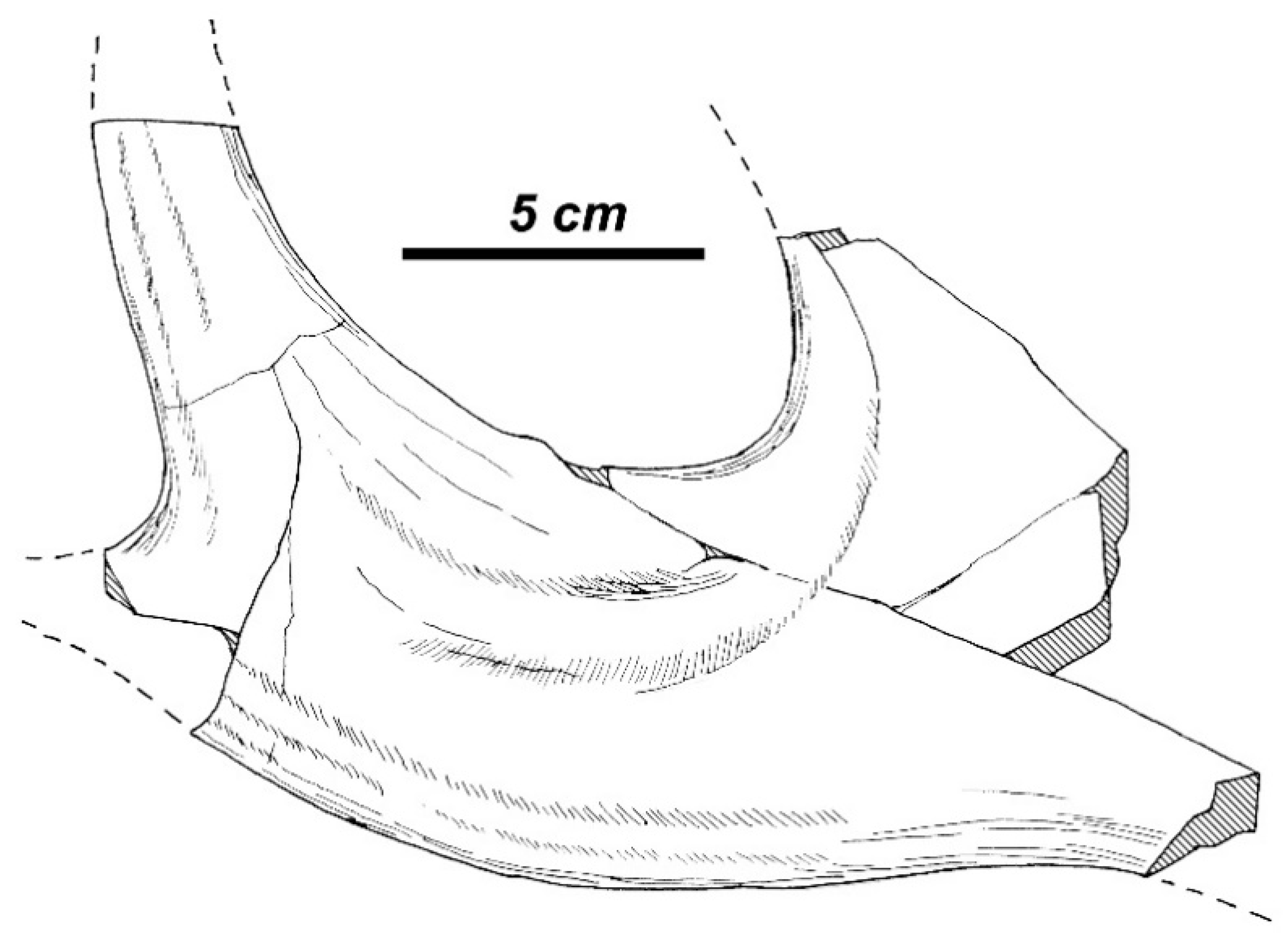
3.3.1. Taxonomic Description
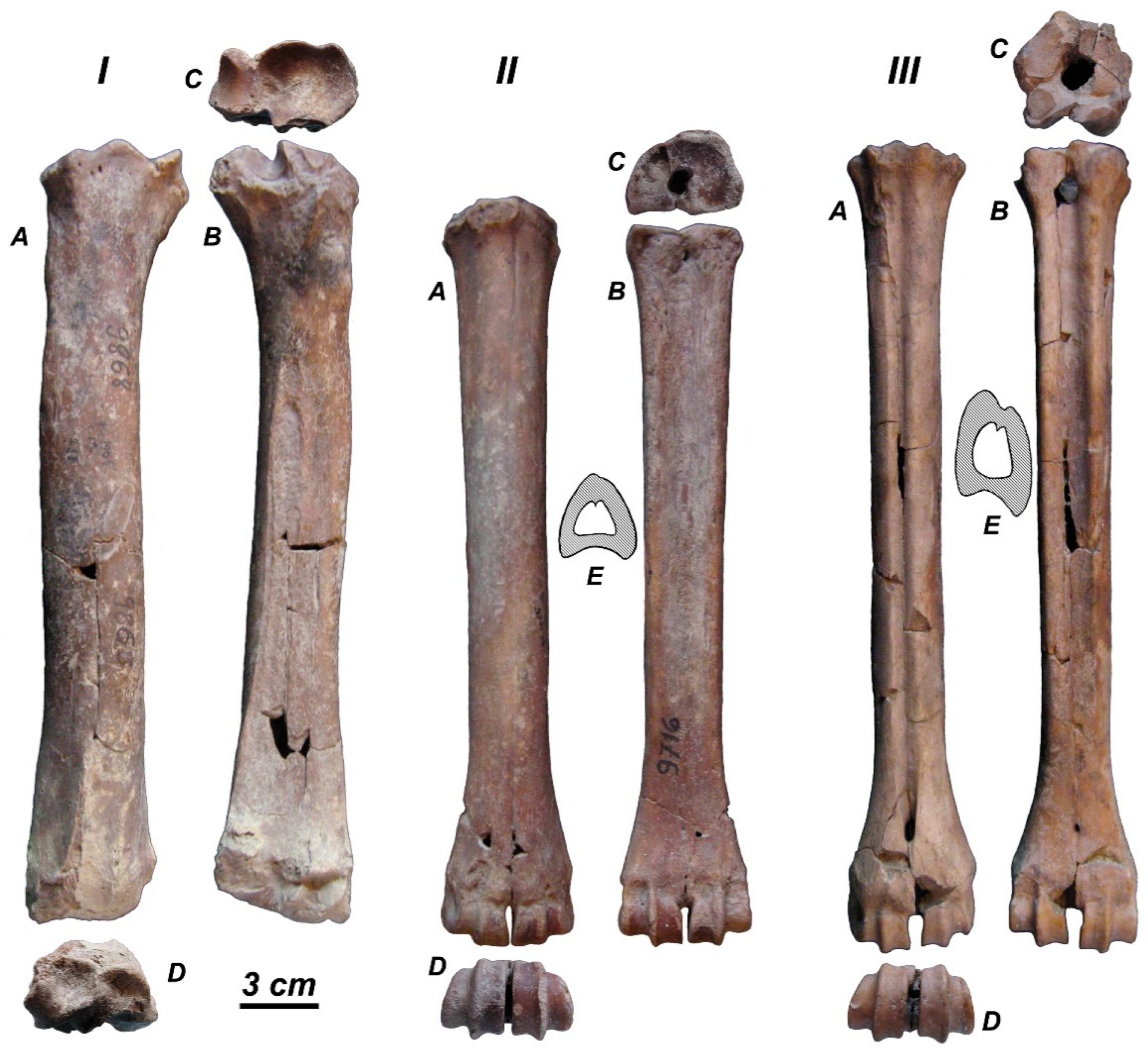
3.3.2. Morphological Description
4. Discussion
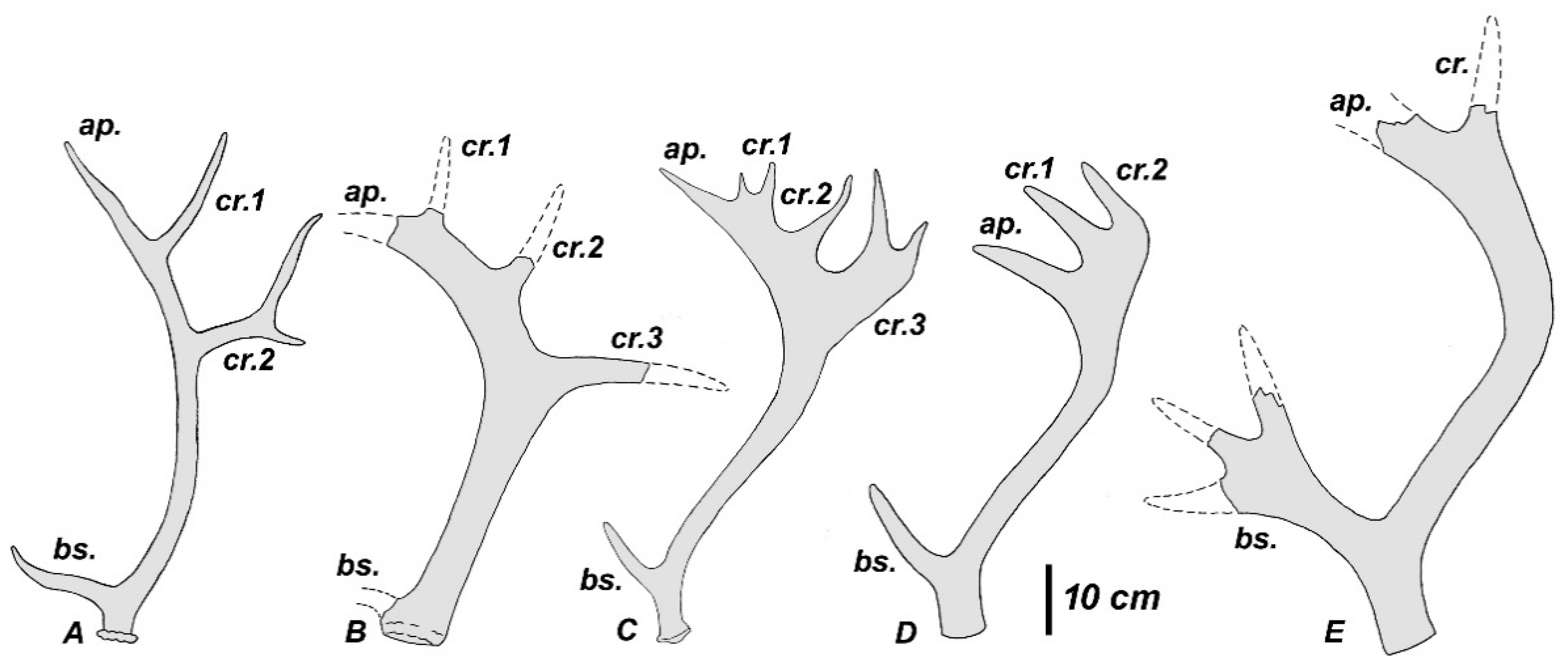
4.1. Plio-Pleistocene Rucervus from Europe
4.2. Plio-Pleistocene Rucervus from the Siwaliks and Caucasus
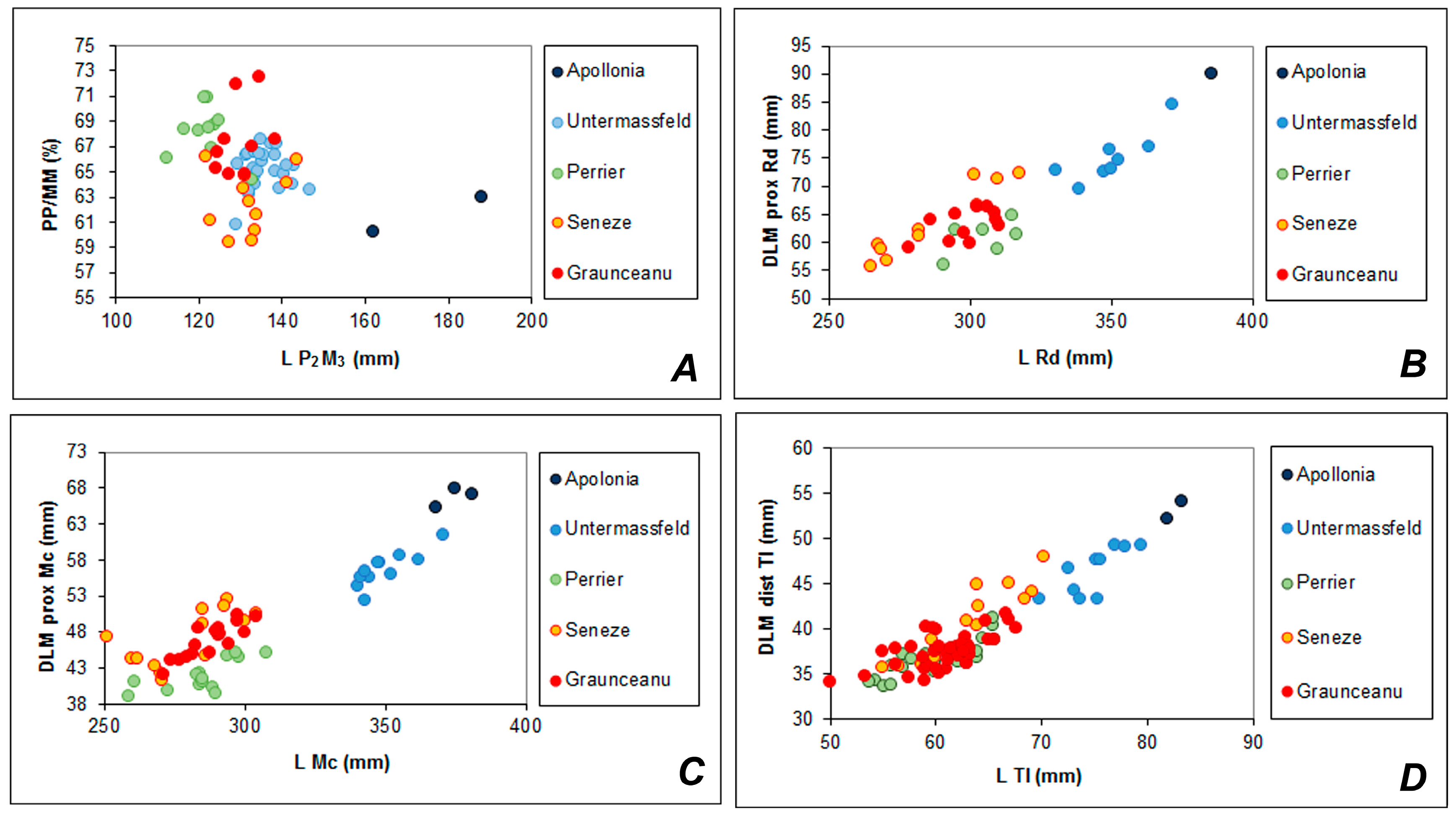
4.3. Morphological Similarities and Geographic Distribution of Rucervus radulescui and Rucervus gigans
4.4. Early Pleistocene Dispersals of Ruminants in Southeastern Europe
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; pp. 1–421. ISBN 0-8117-0496-3. [Google Scholar]
- Ungar, P.S.; Grine, F.E.; Teaford, M.F. Diet in Early Homo: A Review of the Evidence and a New Model of Adaptive Versatility. Annu. Rev. Anthrop. 2006, 35, 209–228. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Croitor, R. Evolution, Ecology, and Biochronology of Herbivore Associations in Europe during the Last 3 Million Years. Quaternaire 2007, 18, 129–152. [Google Scholar] [CrossRef]
- Croitor, R.; Brugal, J.-P. Ecological and evolutionary dynamics of the carnivore community in Europe during the last 3 million years. Quat. Int. 2010, 212, 98–108. [Google Scholar] [CrossRef]
- Curran, S.C. Hominin Paleoecology and Cervid Ecomorphology. Ph.D. Dissertation, University of Minnesota, Minneapolis, MN, USA, 2009. [Google Scholar]
- Curran, S.C. Exploring Eucladoceros ecomorphology using geometric morphometrics. Anatom. Rec. 2015, 298, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe. Rev. Paléobiol. 2003, 22, 197–229. [Google Scholar]
- Spassov, N. Southeastern Europe as a route for the earliest dispersal of Homo toward Europe: Ecological Conditions and the Timing of the First Human Occupation of Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 281–290. ISBN 978-94-024-0873-7. [Google Scholar]
- Strait, D.S.; Orr, C.M.; Hodgkins, J.; Spassov, N.; Gurova, M.; Miller, C.; Tzankov, T. The Human Fossil Record of Bulgaria and the Formulation of Biogeographic Hypotheses. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 69–78. ISBN 978-94-024-0873-7. [Google Scholar]
- Radulescu, C.; Samson, P.M. The Plio-Pleistocene Mammalian Succession of the Olteț Valley, Dacic Basin, Romania. Quartärpaläontologie 1990, 8, 225–232. [Google Scholar]
- Radulescu, C.; Samson, P.M. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Romania. Boll. Soc. Paleont. Ital. 2001, 40, 285–291. [Google Scholar]
- Ghenea, C. The Pliocene-Pleistocene boundary in Romania. In The Pleistocene Boundary and the Beginning of the Quaternary; Van Couvering, J.A., Ed.; Cambridge University Press: Cambridge, UK; New York/Port Chester, NY, USA; Melbourne/Sydney, Australia, 1997; Volume 41, pp. 216–220. ISBN 0521341159. [Google Scholar]
- Radulescu, C.; Samson, P.M.; Petculescu, A.; Stiuca, E. Pliocene large mammals of Romania. Coloq. Paleontol. 2003, 1, 549–558. [Google Scholar]
- Radulescu, C.; Samson, P.M.; Stiuca, E. Cadre biostratigraphique au Paléolithique inférieur en Roumanie. Quaternaire 1998, 9, 283–290. [Google Scholar] [CrossRef]
- Azzaroli, A. Quaternary mammals and the “End-Villafranchian” dispersal event—A turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 44, 113–139. [Google Scholar] [CrossRef]
- Martínez-Navarro, B. Early Pleistocene faunas of Eurasia and hominid dispersals. In Out of Africa I: Who? When? and Where? Vertebrate Paleobiology and Paleoanthropology Series; Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., Leakey, R.E., Eds.; Springer: New York, NY, USA, 2010; pp. 207–224. ISBN 978-90-481-9036-2. [Google Scholar]
- Samson, P.M. Les Equides fossiles de Roumanie. Geol. Rom. 1975, 14, 165–352. [Google Scholar]
- Frost, S.; Ting, N.; Harcourt-Smith, W.; Delson, E. The positional and locomotor behavior of Paradolichopithecus arvernensis as inferred from the functional morphology of the postcrania. Am. J. Primatol. 2005, 66, S134–S135. [Google Scholar]
- Croitor, R. Systematical Position and Evolution of the Genus Arvernoceros (Cervidae, Mammalia) from Plio-Pleistocene of Eurasia. Muzeul Olteniei Craiova Oltenia Studii si communicari S tiintele Naturii 2009, 25, 375–382. [Google Scholar]
- Croitor, R. Deer from Late Miocene to Pleistocene of Western Palearctic: Matching fossil record and molecular phylogeny data. Zitteliana 2014, 32, 115–153. [Google Scholar]
- Kostopoulos, D.S. The Plio-Pleistocene artiodactyls (Vertebrata, Mammalia) of Macedonia 1. The fossiliferous site “Apollonia 1”, Mygdonia basin of Greece. Geodiversitas 1997, 19, 845–875. [Google Scholar]
- Koufos, G.D. New Material and Revision of the Carnivora, Mammalia from the Lower Pleistocene Locality Apollonia 1, Greece. Quaternary 2018, 1, 6. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S. Biochronology and succession of the Plio-Pleistocene macromammalian localities of Greece. Mémoires et travaux de l’Institut de Montpellier 1997, 21, 619–634. [Google Scholar]
- Koufos, G.D.; Syrides, G.E.; Kostopoulos, D.S.; Koliadimou, K.K. Preliminary results about the stratigraphy and the paleoenvironment of Mygdonia basin, Macedonia, Greece. Geobios 1995, 18, 243–249. [Google Scholar] [CrossRef]
- Heintz, E. Les Cervidés Villafranchiens de France et d’Espagne; Mémoires du Muséum national d’Histoire naturelle, Série C; Editions du Museum National d’Histoire Naturelle: Paris, France, 1970; t. I, 330p, t. II, 206p. [Google Scholar]
- Von den Driesch, A. A Guide to the Measurement of Animal Bones from Archaeological Sites; Peabody Museum Press: Salem, MA, USA, 1976; Volume 1, pp. 1–137. ISBN 9780873659505. [Google Scholar]
- Janis, C.M. Correlation of cranial and dental variables with body size in ungulates and macropodoids. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 255–299. ISBN 9780521360999. [Google Scholar]
- Hodgson, B.H. Proceedings of Learned Societies. Linnaean Society, Feb. 20, 1838. Ann. Nat. Hist. 1838, 1, 152–154. [Google Scholar]
- Hodgson, B.H. Specific name and character of a new species of Cervus, discovered by B. H. Hodgson, Esq. in 1825, and indicated in his Catalogue by the local name of Bahraiya. J. Asiatic Soc. Bengal 1835, 4, 648–649. [Google Scholar]
- Hodgson, B.H. Note on Cervus duvaucelii Cuvier, or C. elaphoides and Bahraiya of Hodgson. J. Asiatic Soc. Bengal 1836, 5, 240–242. [Google Scholar]
- Pocock, R.I. The Larger Deer of British India. Part II. J. Bombay Nat. Hist. Soc. 1943, 43, 553–572. [Google Scholar]
- Alekseyeva, L.I. Early Anthropogene Theriofauna of East Europe. Trans. Geol. Inst. 1977, 300, 1–214. (In Russian) [Google Scholar]
- Kostopoulos, D.S.; Athanassiou, A. In the shadow of bovids: Suids, cervids and giraffids from the Plio-Pleistocene of Greece. Quaternaire Hors-Serie 2005, 2, 179–190. [Google Scholar]
- Titov, V.V. Late Pliocene Large Mammals from Northeastern Sea of Azov Region; SSC RAS Publishing: Rostov-upon-Don, Russia, 2008; p. 264. ISBN 978-902982-47-0. (In Russian) [Google Scholar]
- Radulescu, C. Artiodactyles du Pliocène et du Pléistocène inférieur de Roumanie. Quaternaire Hors-série 2005, 2, 191–200. [Google Scholar]
- Colbert, E.H. Siwalik Mammals in the American Museum of Natural History. Trans. Am. Philos. Soc. 1935, 26, 1–401. [Google Scholar] [CrossRef]
- Kahlke, H.-D. Die Cerviden-Reste aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meininghen (Thüringen); Verlag des Römisch-Germanischen Zentralmuseums Mainz: Mainz, Germany, 1997; Volume 40, pp. 181–275. [Google Scholar]
- Croitor, R.; Kostopoulos, D.S. On the Systematic Position of the Large-Sized Deer from Apollonia, Early Pleistocene, Greece. Palaeontol. Zeitsch. 2004, 78, 137–159. [Google Scholar] [CrossRef]
- Croitor, R.; Popescu, A. Large-sized ruminants from the Early Pleistocene of Leu (Oltenia, Romania) with remarks on biogeographical aspects of the “Pachycrocuta event”. Neues Jahrb. Geol. Paläontol.-Abhandlungen 2011, 261, 353–371. [Google Scholar] [CrossRef]
- Depéret, C.; Mayet, L. Le gisement de Senèze et sa faune paléomammalogique. Assoc. Fr. Avanc. Sci. C. R. 1911, 2, 261–263. [Google Scholar]
- Tsoukala, E.; Chatzopoulou, K. A New Early Pleistocene (Latest Villafranchian) Site with mammals in Kalamoto (Mygdonia Basin, Macedonia, Greece)—Preliminary Report. Mitt. Komm. Quartärforsch. Österr. Acad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Spassov, N. Brief review of the Pliocene ungulate fauna of Bulgaria. Quaternaire Hors-Série 2005, 2, 201–212. [Google Scholar]
- Croitor, R.; Stefaniak, K.; Pawłowska, K.; Ridush, B.; Wojtal, P.; Stach, M. Giant deer Megaloceros giganteus Blumenbach, 1799 (Cervidae, Mammalia) from Palaeolithic of Eastern Europe. Quat. Int. 2014, 326–327, 91–104. [Google Scholar] [CrossRef]
- Croitor, R. Plio-Pleistocene Deer of Western Palearctic: Taxonomy, Systematics, Phylogeny; Inst. Zool. of the Acad. of Sc. of Moldova: Chișinău, Moldova, 2018; 140p. [Google Scholar]
- Vislobokova, I.A. Fossil deer of Eurasia. Trans. Paleont. Inst. 1990, 240, 1–206. (In Russian) [Google Scholar]
- David, A. A new deer species (Cervidae, Mammalia) from the late Pliocene deposits of Moldova. Bull. Acad. Şti. Rep. Moldova Ser. Şti. Chim. Biol. 1992, 1, 67–68. (In Russian) [Google Scholar]
- Bajgusheva, V.S. On skull construction of the large-sized deer of the Khaprovian mammal assemblage. In Paleoteriologia; Tatarinov, L.P., Ed.; Nauka: Moscow, Russia, 1994; pp. 236–252. ISBN 5-02-005723-1. (In Russian) [Google Scholar]
- Bajgusheva, V.S.; Titov, V.V.; Tesakov, A.S. The sequence of Plio-Pleistocene mammal faunas from the south Russian Plain (the Azov Region). Boll. Soc. Palaeontol. Ital. 2001, 40, 133–138. [Google Scholar]
- Vislobokova, I.; Sotnikova, M.; Dodonov, A. Late Miocene-Pliocene mammalian faunas of Russia and neighbouring countries. Boll. Soc. Palaeontol. Ital. 2001, 40, 307–313. [Google Scholar]
- Stehlin, H.G. Les sables de Rosières, près Saint-Florent (Cher). II. Paléontologie. Bull. Soc. Géol. Fr. 1912, 12, 198–212. [Google Scholar]
- Guerin, C.; Dewolf, Y.; Lautridou, J.-P. Revision of a famous site: Saint-Prest (Chartres, France). Geobios 2003, 36, 55–82. [Google Scholar] [CrossRef]
- Azzaroli, A. The Deer of the Weybourne Crag and Forest Bed of Norfolk. Bull. Br. Mus. 1953, 2, 1–96. [Google Scholar]
- Croitor, R.; Bonifay, M.-F. Etude préliminaire des cerfs du gisement pléistocène inférieur de Ceyssaguet (Haute-Loire). Paléo 2001, 13, 129–144. [Google Scholar]
- Azzaroli, A. Critical observations upon Siwalik Deer. Proc. Linn. Soc. Lond. 1954, 165, 75–83. [Google Scholar] [CrossRef]
- Croitor, R. Description of a new deer species (Cervidae, Mammalia) from the Early Pliocene of Eastern Europe, with a review of early dispersals and palaeobiogeography of the subfamily Cervinae. Neues Jahrb. Geol. Paläontol.-Abhandlungen 2017, 283, 85–108. [Google Scholar] [CrossRef]
- Arif, M.; Shah, S.M.I.; de Vos, J. Cervus rewati sp. nov. (Mammalia, Cervidae) from the Upper Siwaliks of Pakistan. Mem. Geol. Surv. Pak. 1992, 17, 13–23. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Part II. Molar Teeth and other remains of Mammalia. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1876, 1, 19–87. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Part IV. Supplement to crania of ruminants. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1880, 1, 172–181. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1884, 3, 1–258. [Google Scholar]
- Koken, E. Ueber fossile Saugethiere aus China. Dames und Kayser Palaeontologische Abhandlungen 1885, 3, 31–114. [Google Scholar]
- Brown, B. A new deer form from the Siwaliks. Am. Mus. Novit. 1926, 242, 1–6. [Google Scholar]
- Croitor, R.; Stefaniak, K. Early Pliocene deer of Central and Eastern European regions and inferred phylogenetic relationships. Palaeontographica 2009, 287, 1–39. [Google Scholar] [CrossRef]
- Flerov, K.K. Family Cervidae Gray 1821. In Basics of Paleontology, Mammals; Orlov, Y.A., Gromova, V.I., Eds.; Nedra: Moscow, Russia, 1962; Volume 13, pp. 368–378. ISBN 978-5-458-31474-9. (In Russian) [Google Scholar]
- Flerov, K.K.; Shevyreva, N.S. Pseudalces, Pliocene deer from Ciscaucasus. Paleont. J. 1963, 3, 120–123. (In Russian) [Google Scholar] [CrossRef]
- Vislobokova, I.A. Elks in the Pliocene of the USSR. Quartärpaläontologie 1986, 6, 239–242. [Google Scholar]
- Vekua, A. Kvabebi Fauna of Akchagylian Vertebrates; Nauka: Moscow, Russia, 1972; UDK 566(118.2); 351p. (In Russian) [Google Scholar]
- Vekua, A.; Bendukidze, O.; Kiladze, S. Gigantic deer in Plio-Pleistocene. Proc. Georgian Nation Mus. 2010, 2, 38–53. (In Georgian) [Google Scholar]
- Titov, V.V.; Shvyreva, A.K. Deer of the Genus Megaloceros (Mammalia, Cervidae) from the Lower Pleistocene of Ciscaucasia. Paleont. J. 2016, 1, 82–90. (In Russian) [Google Scholar]
- Grubb, P. Cervidae of Southeast Asia. In Horns, Pronghorns, and Antlers: Evolution, Morphology, Physiology, and Social Significance; Bubenik, G.A., Bubenik, A.B., Eds.; Springer: New York, NY, USA, 1990; pp. 169–179. ISBN 978-1-4613-8968-2. [Google Scholar]
- Dawkins, W.B. Contributions to the history of the Deer of the European Miocene and Pliocene strata. Q. J. Geol. Soc. 1878, 34, 402–420. [Google Scholar] [CrossRef]
- Berlioz, É.; Kostopoulos, D.S.; Blondel, C.; Merceron, G. Feeding ecology of Eucladoceros ctenoides as a proxy to track regional environmental variations in Europe during the early Pleistocene. C. R. Palevol. 2018, 17, 320–332. [Google Scholar] [CrossRef]
- Spassov, N. Acinonyx pardinensis (Croizet et Jobert) remains from the Middle Villafranchian locality of Varshets (Bulgaria) and the Plio-Pleistocene history of the cheetahs in Eurasia. Estud. Geol. 2011, 67, 245–253. [Google Scholar] [CrossRef][Green Version]
- Croitor, R. Paleobiogeography of early human dispersal in western Eurasia: Preliminary results. C. R. Palevol. 2018, 17, 276–286. [Google Scholar] [CrossRef]
- Radulescu, C.; Samson, P.M. Les mammifères du Pléistocène inferieur d-Izvoru (Departement d’Olt, Roumanie). Quartärpaläontologie 1986, 6, 157–171. [Google Scholar]
- Croitor, R. Taxonomy and systematics of large-sized deer of the genus Praemegaceros Portis, 1920 (Cervidae, Mammalia). Courier-Forschungsinstitut Senckenberg 2006, 256, 91–116. [Google Scholar]
- Bukhsianidze, M. Dmanisi artiodactyl assemblage. In Programme and Abstract Book, Proceedings of the XIV Annual Meeting of the European Association of Vertebrate Palaeontologists, Haarlem, The Netherlands, 6–10 July 2016; Holwerda, F., Madern, A., Voeten, D., van Heteren, A., Meijer, H., den Ouden, N., Eds.; KNAW: Haarlem, The Netherlands, 2016; p. 37. [Google Scholar]
- Bukhsianidze, M.; Hertler, C. Large Mammals from the Akhalkalaki Site. In The Role of the Southern Caucasus on Early Human Evolution and Expansion—Refuge, Hub, or Source Area? In Proceedings of the Workshop, Tbilisi, Georgia, 15–20 October 2013; Bruch, A.A., Lordkipanidze, D., Eds.; Georgian National Museum: Tbilisi, Georgia, 2013; p. 25. [Google Scholar]
- Kahlke, H.-D. Neufunde fon Cerviden-resten aus dem Unterpleistozän von Untermaßfeld. Das Pleistozän von Untermaßfeld bei Meningen (Thüringen), Teil 2. Romisch-Germanisches Zentralmuseum 2001, 40, 461–482. [Google Scholar]
- Vislobokova, I.A.; Sotnikova, M.V.; Erbaeva, M.A. The Villafranchian mammalian faunas of the Asiatic part of former USSR. Il Quaternario 1995, 8, 367–376. [Google Scholar]
- Boulbes, N.; Mayda, S.; Titov, V.V.; Alçiçek, M.C. Les grands mammifères du Villafranchien supérieur des travertins du Bassin de Denizli (Sud-Ouest Anatolie, Turquie). L’Anthropologie 2014, 118, 44–73. [Google Scholar] [CrossRef]
- Ivanova, S.; Gurova, M.; Spassov, N.; Popov, V.; Makedonska, J.; Tzankov, T.; Strait, D.S. Preliminary Findings of the Balkan Paleo Project: Evidence of Human Activity at the “Gateway” of Europe during the Late Pleistocene. Be-JA 2012, 2, 1–24. [Google Scholar]
- Mosbrugger, V.; Utescher, T.; Dilcher, D.L. Cenozoic continental climatic evolution of Central Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- Gabunia, L.; Vekua, A.; Lordkipanidze, D. The environmental contexts of early human occupation of Georgia (Transcaucasia). J. Hum. Evol. 2000, 38, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Vekua, A. Die Wirbeltierfauna des Villafranchium von Dmanisi und ihre Biostratigraphische Bedeutung. Jahrb. Römisch-Ger. Zent.mus. Mainz. 1995, 42, 77–180. [Google Scholar]
- Geraads, D. Les Ruminants du Pleistocène d’Oubeidiyeh (Israël). In Les Mammifères du Pléistocène Inférieur de la Vallée du Jourdain à Oubeidiyeh; Tchernov, E., Ed.; Mémoires et Travaux du Centre de Recherche Français de Jerusalem: Paris, France, 1986; Volume 5, pp. 145–173. [Google Scholar]
- Teilhard de Chardin, P.; Trassaert, M. The Piocene Camelidae, Giraffidae, and Cervidae of South Eastern Shansi. Palaeont. Sin. 1937, 1, 1–69. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rucervus radulescui | Rucervus gigans | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GR-690 | BUG-692 | BUG-400 | BUG-294 | BUG-776 | RGU-759 | RGU-836 | 52-403 | APL-357 | APL-212 sin | APL-212 dx | |
| DAP antler base | 43.6 | 43.3 | 38.3 | 43.2 | 42.0 | 59.3 | 60.2 | 61.2 | |||
| DLM antler base | 36.6 | 42.7 | 35.0 | 39.5 | 38.8 | 51.4 | 51.7 | ||||
| H of first ramification | 106.2 | 119.0 | 106.4 | 103.8 | 97.5 | 75.5 | 116.0 | ||||
| DAP pedicle | 40.1 | 38.5 | 35.8 | 55.1 | 56.5 | ||||||
| DLM pedicle | 38.0 | 38.1 | 35.3 | 52.0 | 53.2 | ||||||
| H pedicle | 50.0 | 54.5 | 40.0 | 70.0 | 66.1 | ||||||
| Rucervus radulescui | Rucervus gigans | Rucervus sp. | ||||||
|---|---|---|---|---|---|---|---|---|
| GR-29-1960 | BUG-30108 | RGU-578 | RGU-394 | APL-384 | APL-33 | RGU-836 | RGU-627 | |
| L P2-M3 | 126.2 | 131.1 | 188.0 | 162.0 | 143.8 | 142.1 | ||
| L P2-P4 | 50.0 | 52.5 | 73.3 | 64.4 | 55.0 | 55.0 | ||
| L M1-M3 | 74.0 | 81.0 | 84.5 | 116.4 | 102.0 | 90.0 | 87.7 | |
| PP/MM% | 67.6 | 64.8 | 63.0 | 60.2 | 61.1 | 62.7 | ||
| L P2–symphysis | 53.0 | 74.4 | 58.2 | |||||
| L P2-C | 78.0 | ca. 95 | 102.1 | |||||
| H diastemal part | 23.0 | 21.2 | 23.8 | 22.6 | 16.4 | |||
| H below P2 | 30.0 | 35.0 | 37.7 | 34.6 | 30.1 | |||
| D below P2 | 15.5 | 16.5 | 14.2 | 13.3 | ||||
| H below M1 | 44.8 | 34.5 | 43.1 | 41.3 | 37.0 | |||
| D below M1 | 18.1 | 21.6 | 20.8 | 25.4 | 20.5 | 19.3 | ||
| H below M3 | 44.3 | 50.7 | 45.6 | 39.8 | ||||
| D below M3 | 24.1 | 28.1 | 29.8 | 23.2 | 23.3 | |||
| L P4 | 18.0 | 20.0 | 21.7 | 19.3 | ||||
| D P4 | 12.5 | 12.4 | 12.8 | |||||
| Rucervus radulescui | R. simlicidens | R. colberti | R. gigans | |||||
|---|---|---|---|---|---|---|---|---|
| GR-nn | BUG-nn | BUG-nn | Type | AM-19811 | AM-19792 | AM-19829 | APL-274 | |
| L P2-M3 | 130.0 | 157.1 | ||||||
| L P2-P4 | 58.8 | 71.9 | ||||||
| L M1-M3 | 77.2 | 77.6 | 77.0 | 68.0 | 78.0 | 95.1 | ||
| L P3 | 19.3 | 15.5 | 23.9 | |||||
| D P3 | 20.3 | 18.0 | 23.0 | |||||
| L P4 | 18.5 | 16.1 | 14.0 | 18.0 | 21.1 | |||
| D P4 | 22.3 | 19.7 | 19.0 | 24.0 | 20.6 | |||
| L M1 | 25.8 | 26.3 | 26.8 | 17.0 | 20.0 | 22.0 | 31.2 | |
| D M1 | 22.2 | 22.6 | 23.2 | 20.0 | 24.0 | -- | 29.5 | |
| L M2 | 27.3 | 28.0 | 28.2 | 23.0 | 24.0 | 29.0 | 33.8 | |
| D M2 | 25.0 | 24.2 | 24.4 | 24.5 | 26.5 | 27.0 | 33.0 | |
| L M3 | 27.6 | 29.0 | 26.8 | 22.9 | 26.0 | 26.0 | 28.0 | 31.2 |
| D M3 | 25.5 | 23.3 | 24.0 | 22.9 | 23.0 | 26.0 | 25.0 | 31.0 |
| L | DLMprox | DAPprox | DLMdist | DAPdist | |
|---|---|---|---|---|---|
| Radius | |||||
| V.Gr-19/B-1960 | 303.0 | 66.2 | 36.3 | 60.1 | 41.4 |
| BUG9868 | 298.3 | 61.6 | 33.5 | 53.2 | 39.6 |
| BUG9864 | 310.8 | 63.0 | 33.1 | 58.0 | 42.7 |
| BUG9871 | 286.1 | 63.8 | 30.7 | 53.2 | 35.6 |
| BUG9870 | 308.9 | 65.3 | 33.7 | 57.0 | |
| BUG9877 | 293.0 | 60.0 | 31.0 | 55.6 | 40.2 |
| BUG9911 | 278.4 | 59.0 | 31.5 | 53.8 | |
| BUG9875 | 309.6 | 64.0 | 34.2 | 59.7 | |
| BUG9878 | 300.0 | 59.7 | 33.5 | 55.2 | |
| BUG9876 | 295.2 | 65.0 | 35.2 | 58.9 | |
| BUG9865 | 306.6 | 66.3 | 36.0 | 57.8 | |
| Metacarpus | |||||
| V.Gr52-1960 | 297.6 | 50.2 | 38.5 | 49.5 | 32.0 |
| V.Gr24-1960 | 277.0 | 44.0 | 31.7 | 43.2 | 28.3 |
| Gr1965/9743 | 297.8 | 49.5 | 34.1 | 49.7 | 32.0 |
| Gr1965/9716 | 282.5 | 46.1 | 33.1 | 49.9 | 31.2 |
| BUG9732 | 290.8 | 48.5 | 35.0 | 50.0 | 32.4 |
| BUG9752 | 273.7 | 44.0 | 31.0 | 45.7 | 27.5 |
| BUG9799 | 291.0 | 47.5 | 33.5 | 50.4 | 32.0 |
| BUG9754 | 294.5 | 46.2 | 34.8 | 47.2 | 30.5 |
| BUG9757 | 281.5 | 44.8 | 32.5 | 31.0 | |
| BUG9727 | 275.8 | 43.6 | 29.5 | ||
| BUG9724 | 279.7 | 44.5 | 34.0 | 45.0 | 31.0 |
| BUG9744 | 300.0 | 47.9 | 35.0 | 49.8 | 32.2 |
| BUG9722 | 271.4 | 42.0 | |||
| BUG9730 | 288.0 | 45.0 | 49.8 | 33.0 | |
| BUG9738 | 304.5 | 50.0 | 38.2 | 50.1 | 30.1 |
| BUG9741 | 291.3 | 47.6 | 35.5 | 51.1 | 32.3 |
| BUG9729 | 289.7 | 48.0 | 35.0 | ||
| BUG9769 | 283.4 | 48.5 | 34.2 | 50.0 | 31.5 |
| Metatarsus | |||||
| V.Gr47/D-1964 | 312.0 | 44.8 | 46.1 | 49.5 | 32.3 |
| V.Gr53/960 | 305.0 | 43.0 | 44.6 | 46.6 | 32.7 |
| V.Gr36/D-1960 | 316.5 | 44.2 | 46.3 | 52.0 | 33.0 |
| BUG9681 | 310.0 | 45.0 | 50.0 | 52.4 | 33.4 |
| BUG9639 | 308.3 | 45.0 | 45.8 | 49.6 | 31.5 |
| BUG9618 | 300.5 | 43.2 | 44.8 | 33.9 | |
| NN | 317.0 | 49.0 | 50.0 | 55.1 | 35.0 |
| BUG9629 | 290.3 | 40.3 | 45.5 | 48.7 | 31.2 |
| BUG9645 | 311.0 | 46.0 | 49.0 | 50.1 | 32.7 |
| BUG2027 | 40.8 | 45.4 | 49.1 | 30.2 |
| L | DLMdist | DAPdist | |
|---|---|---|---|
| V.Gr16/A/1960 | 64.8 | 40.8 | 30.8 |
| BUG-9242 | 66.7 | 41.6 | 33.0 |
| BUG-9275 | 62.8 | 39.0 | 32.2 |
| BUG-9248 | 59.2 | 40.2 | 30.9 |
| BUG-9251 | 62.0 | 38.0 | 28.5 |
| BUG-9255 | 65.0 | 38.8 | 32.3 |
| BUG-9241 | 56.3 | 36.0 | 27.3 |
| BUG-9257 | 63.0 | 36.1 | 30.5 |
| BUG-9247 | 60.0 | 37.4 | 30.0 |
| BUG-2064 | 61.3 | 37.6 | 30.0 |
| BUG-nn | 53.3 | 34.7 | 24.7 |
| BUG-9256 | 63.1 | 38.1 | 30.4 |
| BUG-9246 | 61.0 | 37.3 | 30.2 |
| BUG-9259 | 67.7 | 40.1 | 32.0 |
| BUG-9252 | 63.0 | 36.2 | 31.4 |
| BUG-9250 | 59.0 | 34.2 | |
| BUG-61 | 62.6 | 38.2 | 31.0 |
| ISER-nn | 55.0 | 37.4 | 27.7 |
| ISER-nn | 56.3 | 37.8 | 26.0 |
| ISER-nn | 63.2 | 37.2 | 29.6 |
| ISER-nn | 62.2 | 37. | 29.8 |
| Bug60/10 | 59.0 | 35.5 | 28.4 |
| Bug86/29 | 63.3 | 37.7 | 28.9 |
| Bug60/137 | 67.0 | 41.0 | 53.0 |
| Bug60/5ia-C2-6 | 61.0 | 35.5 | 28.2 |
| Bug60/172 | 65.6 | 38.8 | 30.3 |
| B60/127 | 60.4 | 38.0 | 30.3 |
| B60/139 | 62.7 | 37.5 | 30.0 |
| Bug60/131 | 59.3 | 35.9 | 28.4 |
| Bug60/5iaC26-10 | 57.5 | 34.5 | 26.4 |
| Bug60 | 60.1 | 39.9 | 29.5 |
| Bug60/128 | 57.8 | 37.9 | 29.0 |
| Bug60/207 | 50.0 | 34.1 | 25.8 |
| Gr963 | 58.8 | 36.8 | 27.5 |
| Bug60/132 | 60.4 | 35.0 | 28.6 |
| Bug60/130 | 59.8 | 40.1 | 31.0 |
| Bug60/135 | 60.1 | 35.5 | 28.0 |
| Bug60/136 | 61.2 | 36.4 | |
| ISER-nn | 61.5 | 37.8 | 30.1 |
| Frontal Breadth (FB) | Breadth Behind Pedicles (BBP) | BBP/FB % | |
|---|---|---|---|
| APL-212 | 168.7 | 122.7 | 73.0 |
| RGU-774 | 120.0 | 100.0 | 83.3 |
| RGU-239 | 123.4 | 96.3 | 78.0 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Croitor, R. A Description of Two New Species of the Genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with Comments on Hominin and South Asian Ruminants Dispersals. Quaternary 2018, 1, 17. https://doi.org/10.3390/quat1020017
Croitor R. A Description of Two New Species of the Genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with Comments on Hominin and South Asian Ruminants Dispersals. Quaternary. 2018; 1(2):17. https://doi.org/10.3390/quat1020017
Chicago/Turabian StyleCroitor, Roman. 2018. "A Description of Two New Species of the Genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with Comments on Hominin and South Asian Ruminants Dispersals" Quaternary 1, no. 2: 17. https://doi.org/10.3390/quat1020017
APA StyleCroitor, R. (2018). A Description of Two New Species of the Genus Rucervus (Cervidae, Mammalia) from the Early Pleistocene of Southeast Europe, with Comments on Hominin and South Asian Ruminants Dispersals. Quaternary, 1(2), 17. https://doi.org/10.3390/quat1020017