
Aligned Collagen-CNT Nanofibrils and the Modulation Effect on Ovarian Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Matrix Preparation
2.2. Cell Culture and Characterization
2.3. Immunofluorescent Imaging
2.4. AFM Measurements
2.5. RT-qPCR Analysis
3. Results
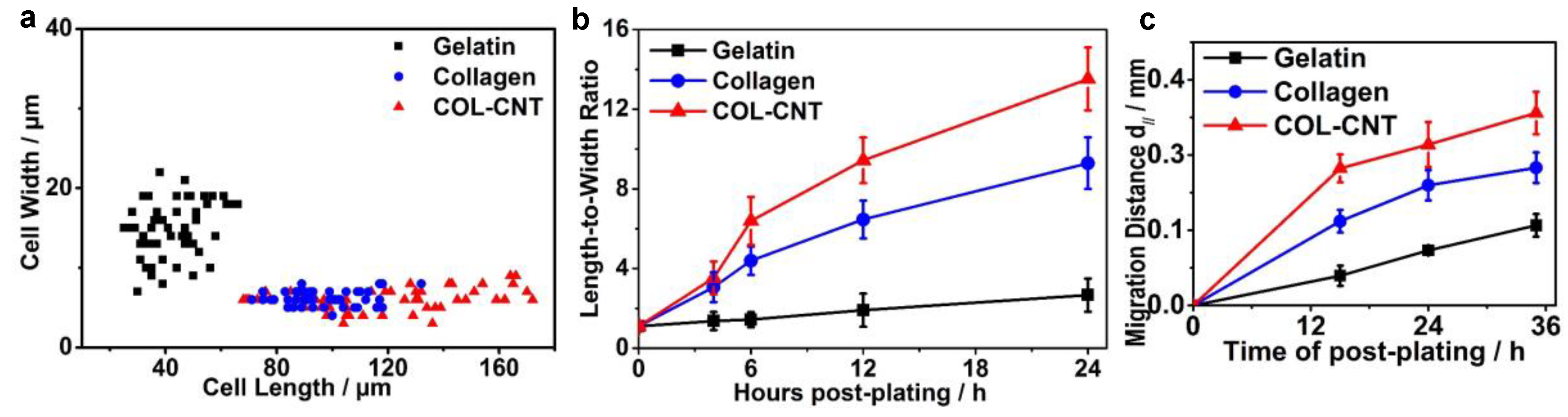
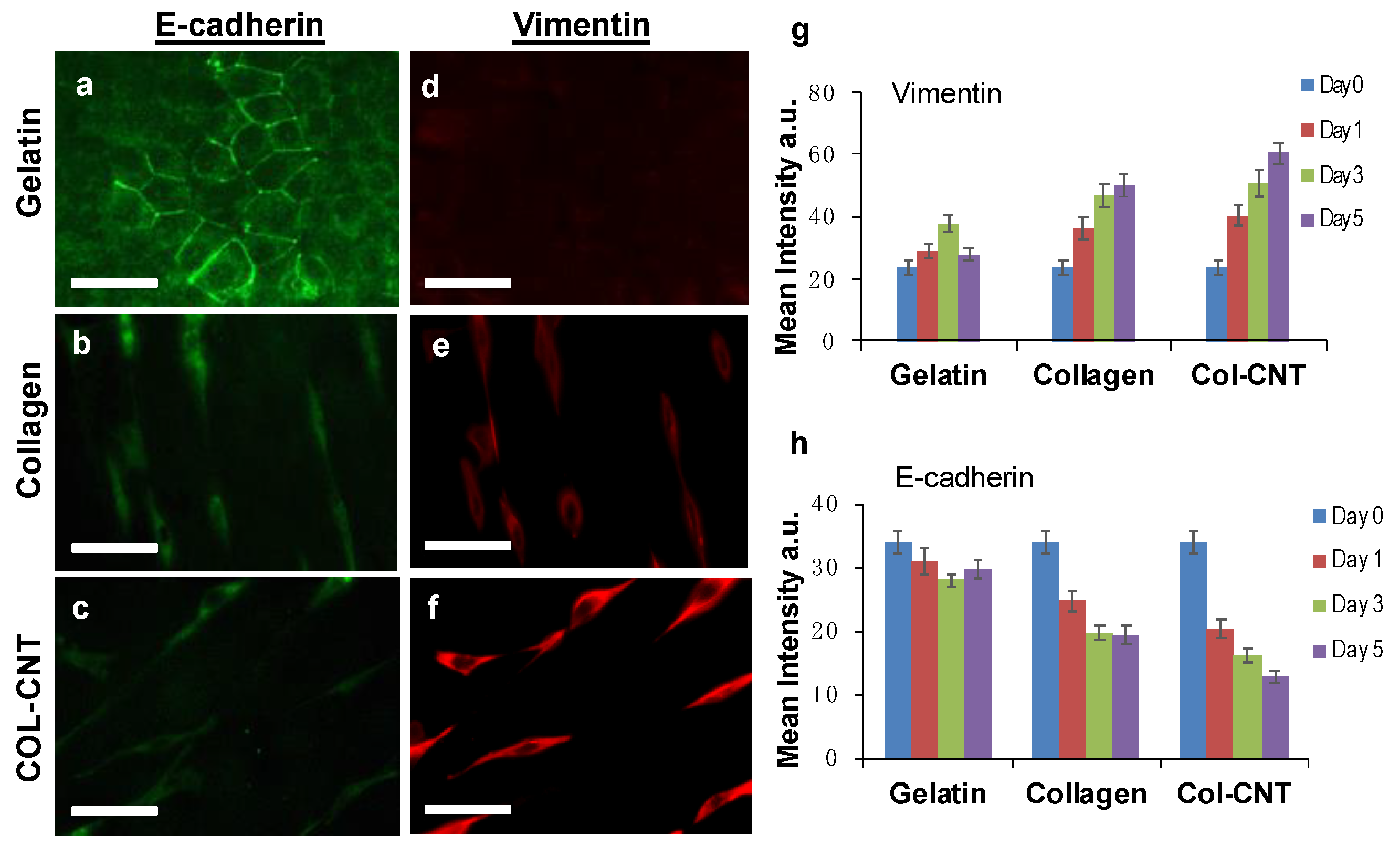
3.1. Aligned Collagen and COL-CNT Fibrils with Matrix-Mediated Cell Polarization and Migration
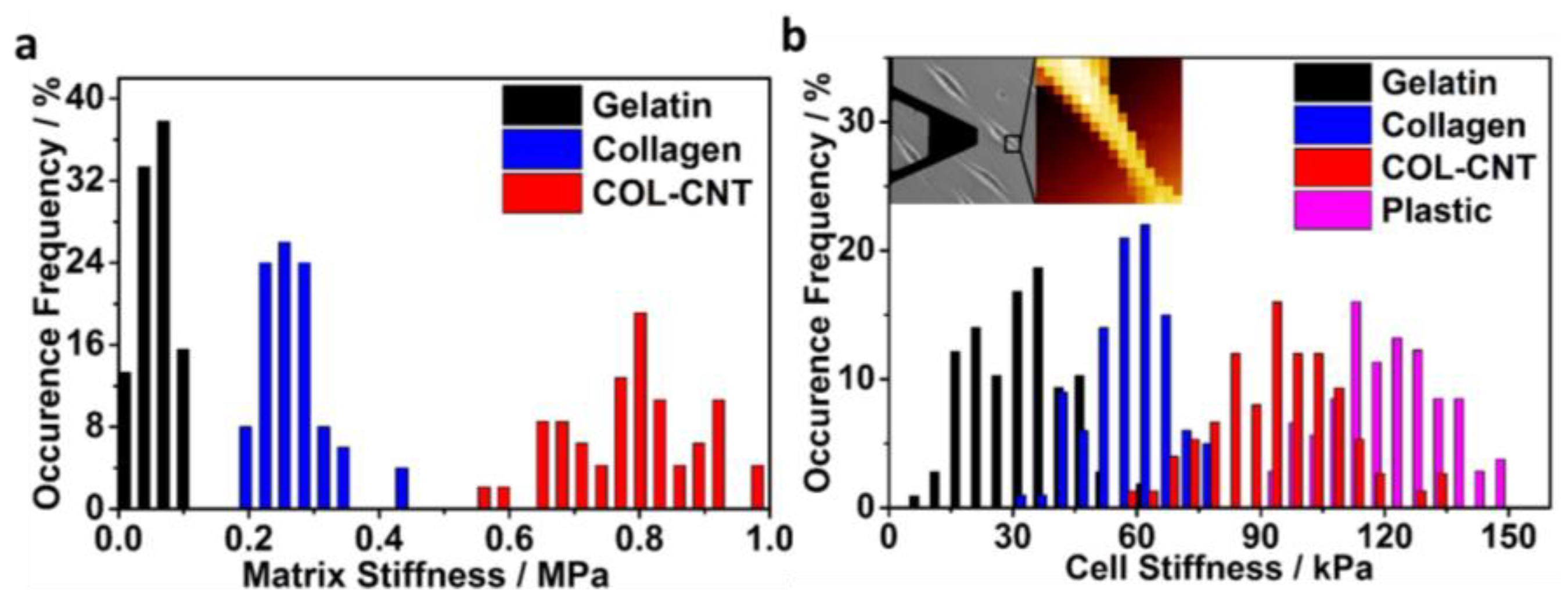
3.2. Characterization of Fibril Stiffness and Cell Stiffness
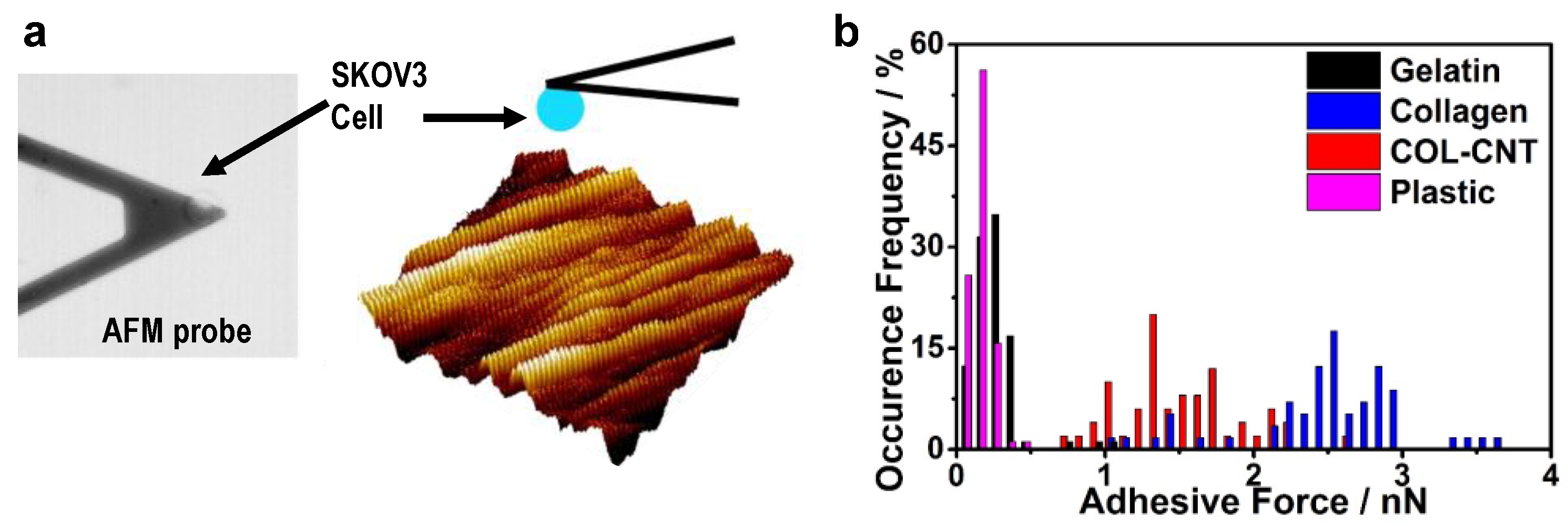
3.3. Cell Adhesion
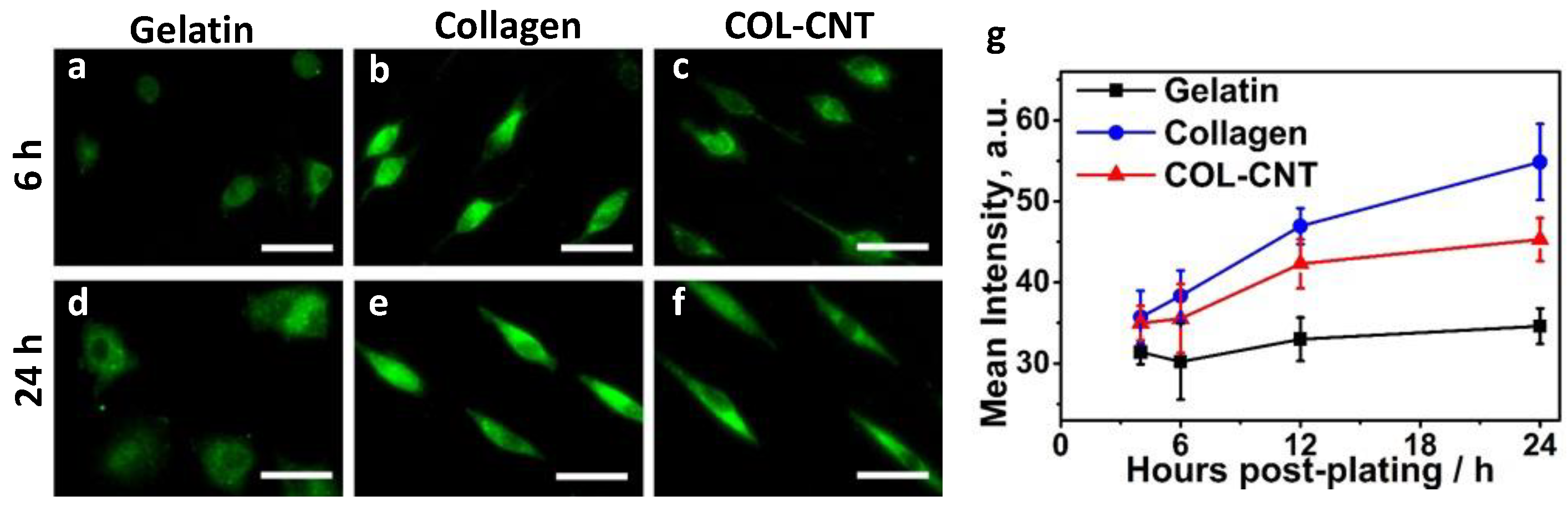
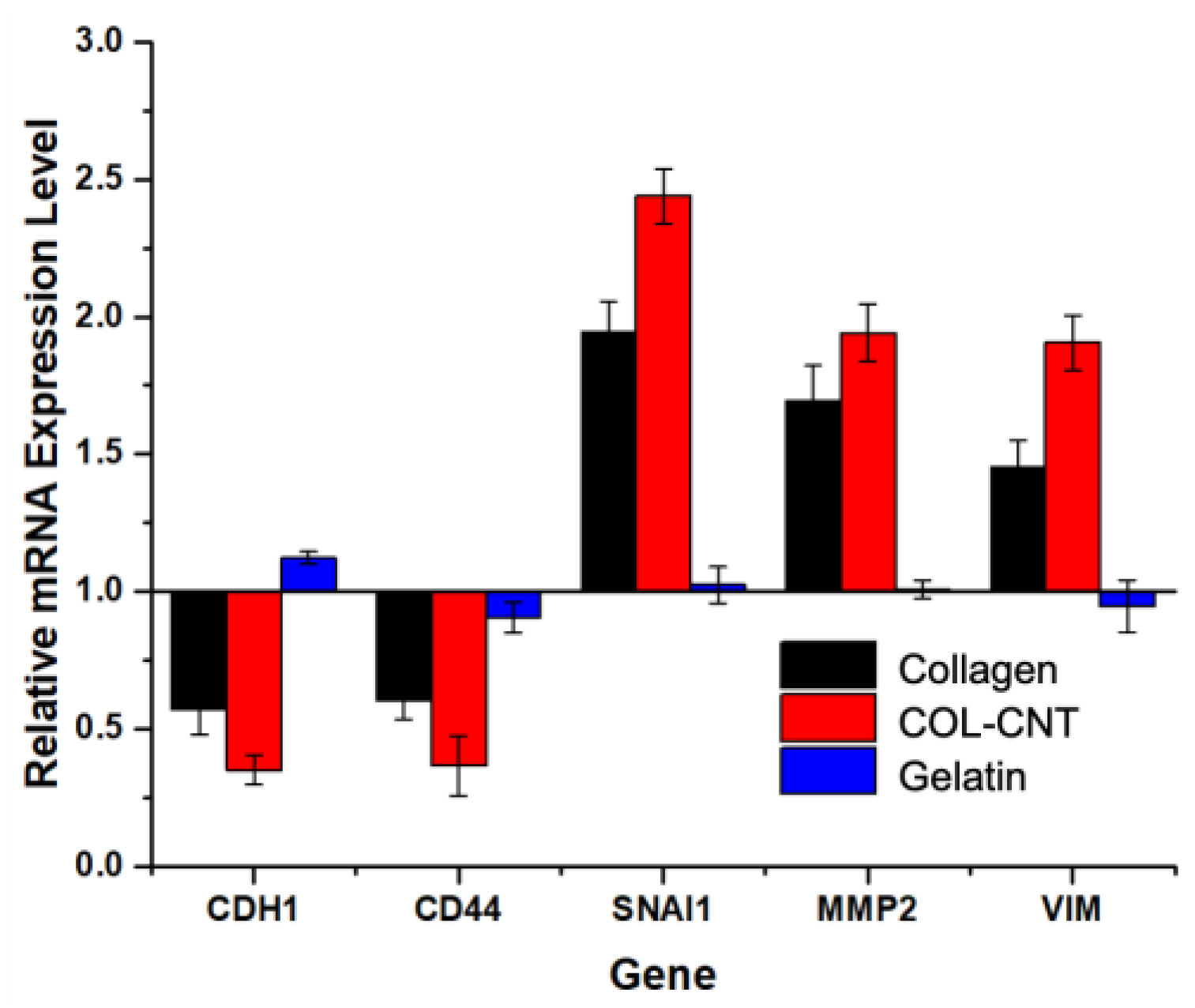
3.4. Collagen and COL-CNT Induced Cell Transformation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Liu, Z.; Cai, W.; He, L.; Nakayama, N.; Chen, K.; Sun, X.; Chen, X.; Dai, H. In Vivo Biodistribution and Highly Efficient Tumour Targeting of Carbon Nanotubes in Mice. Nat. Nanotechnol. 2007, 2, 47–52. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, R.; Laurenzi, B.; Viswanathan, G.; Ajayan, P.; Stegemann, J. Collagen-Carbon Nanotube Composite Materials as Scaffolds in Tissue Engineering. J. Biomed. Mater. Res. A 2005, 74, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Veetil, J.V.; Ye, K. Tailored Carbon Nanotubes for Tissue Engineering Applications. Biotechnol. Prog. 2009, 25, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; Wang, W.; Dunne, N.; Li, X. Applications of Carbon Nanotubes in Bone Tissue Regeneration and Engineering: Superiority, Concerns, Current Advancements, and Prospects. Nanomaterials 2019, 9, 1501. [Google Scholar] [CrossRef]
- Ravanbakhsh, H.; Bao, G.; Mongeau, L. Carbon Nanotubes Promote Cell Migration in Hydrogels. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, F.M.P.; Santos, A.K.; Gomes, K.N.; Lorençon, E.; Guatimosim, S.; Ladeira, L.O.; Resende, R.R. Carbon Nanotube Interaction with Extracellular Matrix Proteins Producing Scaffolds for Tissue Engineering. Int. J. Nanomed. 2012, 7, 4511–4529. [Google Scholar] [CrossRef]
- Chi, N.; Wang, R. Electrospun Protein-CNT Composite Fibers and the Application in Fibroblast Stimulation. Biochem. Biophys. Res. Commun. 2018, 504, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Chi, N.; Zheng, S.; Clutter, E.; Wang, R. Silk-CNT Mediated Fibroblast Stimulation toward Chronic Wound Repair. Recent Prog. Mater. 2019, 1. [Google Scholar] [CrossRef]
- Sridharan, I.; Kim, T.; Wang, R. Adapting Collagen/CNT Matrix in Directing HESC Differentiation. Biochem. Biophys. Res. Commun. 2009, 381, 508–512. [Google Scholar] [CrossRef]
- Sridharan, I.; Kim, T.; Strakova, Z.; Wang, R. Matrix-Specified Differentiation of Human Decidua Parietalis Placental Stem Cells. Biochem. Biophys. Res. Commun. 2013, 437, 489–495. [Google Scholar] [CrossRef][Green Version]
- Kim, T.; Sridharan, I.; Zhu, B.; Orgel, J.; Wang, R. Effect of CNT on Collagen Fiber Structure, Stiffness Assembly Kinetics and Stem Cell Differentiation. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 49, 281–289. [Google Scholar] [CrossRef]
- Bottini, M.; Rosato, N.; Bottini, N. PEG-Modified Carbon Nanotubes in Biomedicine: Current Status and Challenges Ahead. Biomacromolecules 2011, 12, 3381–3393. [Google Scholar] [CrossRef]
- McLane, J.S.; Rivet, C.J.; Gilbert, R.J.; Ligon, L.A. A Biomaterial Model of Tumor Stromal Microenvironment Promotes Mesenchymal Morphology but Not Epithelial to Mesenchymal Transition in Epithelial Cells. Acta Biomater. 2014, 10, 4811–4821. [Google Scholar] [CrossRef]
- Kim, T.; Sridharan, I.; Ma, Y.; Zhu, B.; Chi, N.; Kobak, W.; Rotmensch, J.; Schieber, J.D.; Wang, R. Identifying Distinct Nanoscopic Features of Native Collagen Fibrils towards Early Diagnosis of Pelvic Organ Prolapse. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 667–675. [Google Scholar] [CrossRef]
- Sridharan, I.; Ma, Y.; Kim, T.; Kobak, W.; Rotmensch, J.; Wang, R. Structural and Mechanical Profiles of Native Collagen Fibers in Vaginal Wall Connective Tissues. Biomaterials 2012, 33, 1520–1527. [Google Scholar] [CrossRef]
- Cheng, J.-C.; Leung, P.C.K. Type I Collagen Down-Regulates E-Cadherin Expression by Increasing PI3KCA in Cancer Cells. Cancer Lett. 2011, 304, 107–116. [Google Scholar] [CrossRef]
- Imamichi, Y.; Menke, A. Signaling Pathways Involved in Collagen-Induced Disruption of the E-Cadherin Complex during Epithelial-Mesenchymal Transition. Cells Tissues Organs 2007, 185, 180–190. [Google Scholar] [CrossRef]
- Jordan, N.V.; Johnson, G.L.; Abell, A.N. Tracking the Intermediate Stages of Epithelial-Mesenchymal Transition in Epithelial Stem Cells and Cancer. Cell Cycle Georget. Tex 2011, 10, 2865–2873. [Google Scholar] [CrossRef]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and BHLH Factors in Tumour Progression: An Alliance against the Epithelial Phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Kievit, F.M.; Cooper, A.; Jana, S.; Leung, M.C.; Wang, K.; Edmondson, D.; Wood, D.; Lee, J.S.H.; Ellenbogen, R.G.; Zhang, M. Aligned Chitosan-Polycaprolactone Polyblend Nanofibers Promote the Migration of Glioblastoma Cells. Adv. Healthc. Mater. 2013, 2, 1651–1659. [Google Scholar] [CrossRef]
- Saha, S.; Duan, X.; Wu, L.; Lo, P.-K.; Chen, H.; Wang, Q. Electrospun Fibrous Scaffolds Promote Breast Cancer Cell Alignment and Epithelial-Mesenchymal Transition. Langmuir 2012, 28, 2028–2034. [Google Scholar] [CrossRef]
- Foroni, L.; Vasuri, F.; Valente, S.; Gualandi, C.; Focarete, M.L.; Caprara, G.; Scandola, M.; D’Errico-Grigioni, A.; Pasquinelli, G. The Role of 3D Microenvironmental Organization in MCF-7 Epithelial-Mesenchymal Transition after 7 Culture Days. Exp. Cell Res. 2013, 319, 1515–1522. [Google Scholar] [CrossRef]
- Wei, S.C.; Fattet, L.; Tsai, J.H.; Guo, Y.; Pai, V.H.; Majeski, H.E.; Chen, A.C.; Sah, R.L.; Taylor, S.S.; Engler, A.J.; et al. Matrix Stiffness Drives Epithelial–Mesenchymal Transition and Tumour Metastasis through a TWIST1–G3BP2 Mechanotransduction Pathway. Nat. Cell Biol. 2015, 17, 678–688. [Google Scholar] [CrossRef]
- Li, W.; Zhu, B.; Strakova, Z.; Wang, R. Two-Way Regulation between Cells and Aligned Collagen Fibrils: Local 3D Matrix Formation and Accelerated Neural Differentiation of Human Decidua Parietalis Placental Stem Cells. Biochem. Biophys. Res. Commun. 2014, 450, 1377–1382. [Google Scholar] [CrossRef]
- Leow, W.W.; Hwang, W. Epitaxially Guided Assembly of Collagen Layers on Mica Surfaces. Langmuir ACS J. Surf. Colloids 2011, 27, 10907–10913. [Google Scholar] [CrossRef]
- Lee, C.H.; Shin, H.J.; Cho, I.H.; Kang, Y.-M.; Kim, I.A.; Park, K.-D.; Shin, J.-W. Nanofiber Alignment and Direction of Mechanical Strain Affect the ECM Production of Human ACL Fibroblast. Biomaterials 2005, 26, 1261–1270. [Google Scholar] [CrossRef]
- Li, W.; Chi, N.; Rathnayake, R.A.C.; Wang, R. Distinctive Roles of Fibrillar Collagen I and Collagen III in Mediating Fibroblast-Matrix Interaction: A Nanoscopic Study. Biochem. Biophys. Res. Commun. 2021, 560, 66–71. [Google Scholar] [CrossRef]
- Zhu, B.; Li, W.; Lewis, R.V.; Segre, C.U.; Wang, R. E-Spun Composite Fibers of Collagen and Dragline Silk Protein: Fiber Mechanics, Biocompatibility, and Application in Stem Cell Differentiation. Biomacromolecules 2015, 16, 202–213. [Google Scholar] [CrossRef]
- Touhami, A.; Nysten, B.; Dufrêne, Y.F. Nanoscale Mapping of the Elasticity of Microbial Cells by Atomic Force Microscopy. Langmuir 2003, 19, 4539–4543. [Google Scholar] [CrossRef]
- Heim, A.; Matthews, W.; Koob, T. Determination of the Elastic Modulus of Native Collagen Fibrils via Radial Indentation. Appl. Phys. Lett. 2006, 89, 181902. [Google Scholar] [CrossRef]
- Sariisik, E.; Docheva, D.; Padula, D.; Popov, C.; Opfer, J.; Schieker, M.; Clausen-Schaumann, H.; Benoit, M. Probing the Interaction Forces of Prostate Cancer Cells with Collagen I and Bone Marrow Derived Stem Cells on the Single Cell Level. PLoS ONE 2013, 8, e57706. [Google Scholar] [CrossRef]
- Zamir, E.; Geiger, B. Molecular Complexity and Dynamics of Cell-Matrix Adhesions. J. Cell Sci. 2001, 114, 3583–3590. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Schwartz, M.A.; Burridge, K.; Firtel, R.A.; Ginsberg, M.H.; Borisy, G.; Parsons, J.T.; Horwitz, A.R. Cell Migration: Integrating Signals from Front to Back. Science 2003, 302, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Brakebusch, C.; Fässler, R. The Integrin-Actin Connection, an Eternal Love Affair. EMBO J. 2003, 22, 2324–2333. [Google Scholar] [CrossRef] [PubMed]
- Casey, R.C.; Burleson, K.M.; Skubitz, K.M.; Pambuccian, S.E.; Oegema, T.R.; Ruff, L.E.; Skubitz, A.P. Beta 1-Integrins Regulate the Formation and Adhesion of Ovarian Carcinoma Multicellular Spheroids. Am. J. Pathol. 2001, 159, 2071–2080. [Google Scholar] [CrossRef]
- Jiao, J.; Huang, L.; Ye, F.; Shi, M.; Cheng, X.; Wang, X.; Hu, D.; Xie, X.; Lu, W. Cyclin D1 Affects Epithelial-Mesenchymal Transition in Epithelial Ovarian Cancer Stem Cell-like Cells. OncoTargets Ther. 2013, 6, 667–677. [Google Scholar] [CrossRef]
- Radisky, D.C.; LaBarge, M.A. Epithelial-Mesenchymal Transition and the Stem Cell Phenotype. Cell Stem Cell 2008, 2, 511–512. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.-J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The Epithelial-Mesenchymal Transition Generates Cells with Properties of Stem Cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef]
- Cho, S.H.; Park, Y.S.; Kim, H.J.; Kim, C.H.; Lim, S.W.; Huh, J.W.; Lee, J.H.; Kim, H.R. CD44 Enhances the Epithelial-Mesenchymal Transition in Association with Colon Cancer Invasion. Int. J. Oncol. 2012, 41, 211–218. [Google Scholar] [CrossRef]
- Sacks, J.D.; Barbolina, M.V. Expression and Function of CD44 in Epithelial Ovarian Carcinoma. Biomolecules 2015, 5, 3051–3066. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, L.; Sancho-Torres, I.; Mesonero, C.; Gibbon, D.G.; Shih, W.J.; Zotalis, G. The CD44 Receptor Is a Molecular Predictor of Survival in Ovarian Cancer. Med. Oncol. Northwood Lond. Engl. 2003, 20, 255–263. [Google Scholar] [CrossRef]
- Jeleniewicz, W.; Cybulski, M.; Nowakowski, A.; Stenzel-Bembenek, A.; Guz, M.; Marzec-Kotarska, B.; Kotarski, J.; Stepulak, A. MMP-2 MRNA Expression in Ovarian Cancer Tissues Predicts Patients’ Response to Platinum-Taxane Chemotherapy. Anticancer Res. 2019, 39, 1821–1827. [Google Scholar] [CrossRef]
- Al-Alem, L.; Curry, T.E. Ovarian Cancer: Involvement of the Matrix Metalloproteinases. Reprod. Camb. Engl. 2015, 150, R55–R64. [Google Scholar] [CrossRef]
- Parry, D.A.; Barnes, G.R.; Craig, A.S. A Comparison of the Size Distribution of Collagen Fibrils in Connective Tissues as a Function of Age and a Possible Relation between Fibril Size Distribution and Mechanical Properties. Proc. R. Soc. Lond. B Biol. Sci. 1978, 203, 305–321. [Google Scholar] [CrossRef]
- Parry, D.A. The Molecular and Fibrillar Structure of Collagen and Its Relationship to the Mechanical Properties of Connective Tissue. Biophys. Chem. 1988, 29, 195–209. [Google Scholar] [CrossRef]
- San Antonio, J.D.; Lander, A.D.; Karnovsky, M.J.; Slayter, H.S. Mapping the Heparin-Binding Sites on Type I Collagen Monomers and Fibrils. J. Cell Biol. 1994, 125, 1179–1188. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Tumour-Cell Invasion and Migration: Diversity and Escape Mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- Pathak, A.; Kumar, S. Biophysical Regulation of Tumor Cell Invasion: Moving beyond Matrix Stiffness. Integr. Biol. Quant. Biosci. Nano Macro 2011, 3, 267–278. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Plasticity of Cell Migration: A Multiscale Tuning Model. J. Cell Biol. 2010, 188, 11–19. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Chi, N.; Clutter, E.D.; Zhu, B.; Wang, R.R. Aligned Collagen-CNT Nanofibrils and the Modulation Effect on Ovarian Cancer Cells. J. Compos. Sci. 2021, 5, 148. https://doi.org/10.3390/jcs5060148
Li W, Chi N, Clutter ED, Zhu B, Wang RR. Aligned Collagen-CNT Nanofibrils and the Modulation Effect on Ovarian Cancer Cells. Journal of Composites Science. 2021; 5(6):148. https://doi.org/10.3390/jcs5060148
Chicago/Turabian StyleLi, Wen, Naiwei Chi, Elwin D. Clutter, Bofan Zhu, and Rong R. Wang. 2021. "Aligned Collagen-CNT Nanofibrils and the Modulation Effect on Ovarian Cancer Cells" Journal of Composites Science 5, no. 6: 148. https://doi.org/10.3390/jcs5060148
APA StyleLi, W., Chi, N., Clutter, E. D., Zhu, B., & Wang, R. R. (2021). Aligned Collagen-CNT Nanofibrils and the Modulation Effect on Ovarian Cancer Cells. Journal of Composites Science, 5(6), 148. https://doi.org/10.3390/jcs5060148