Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains †


 and
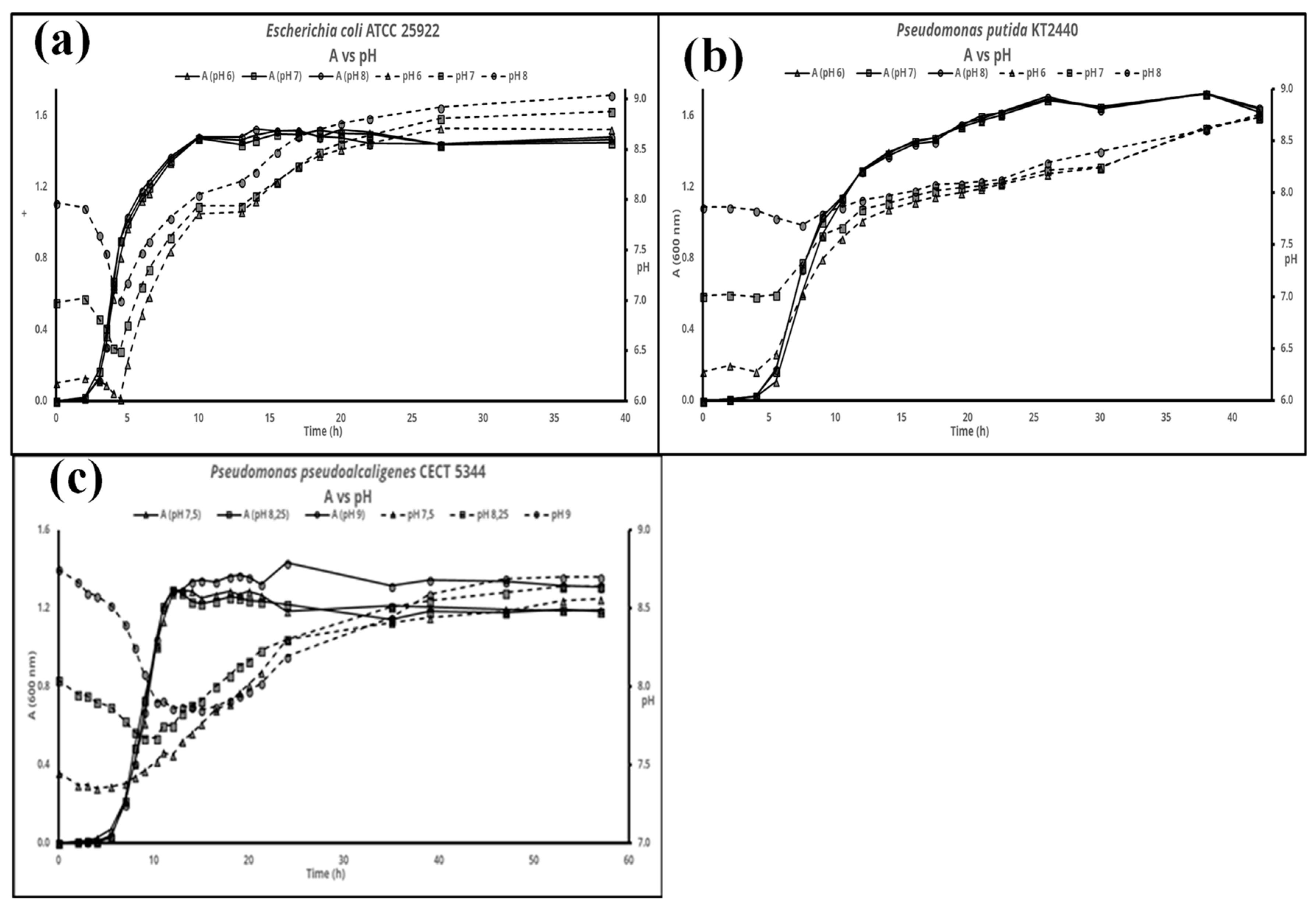
and 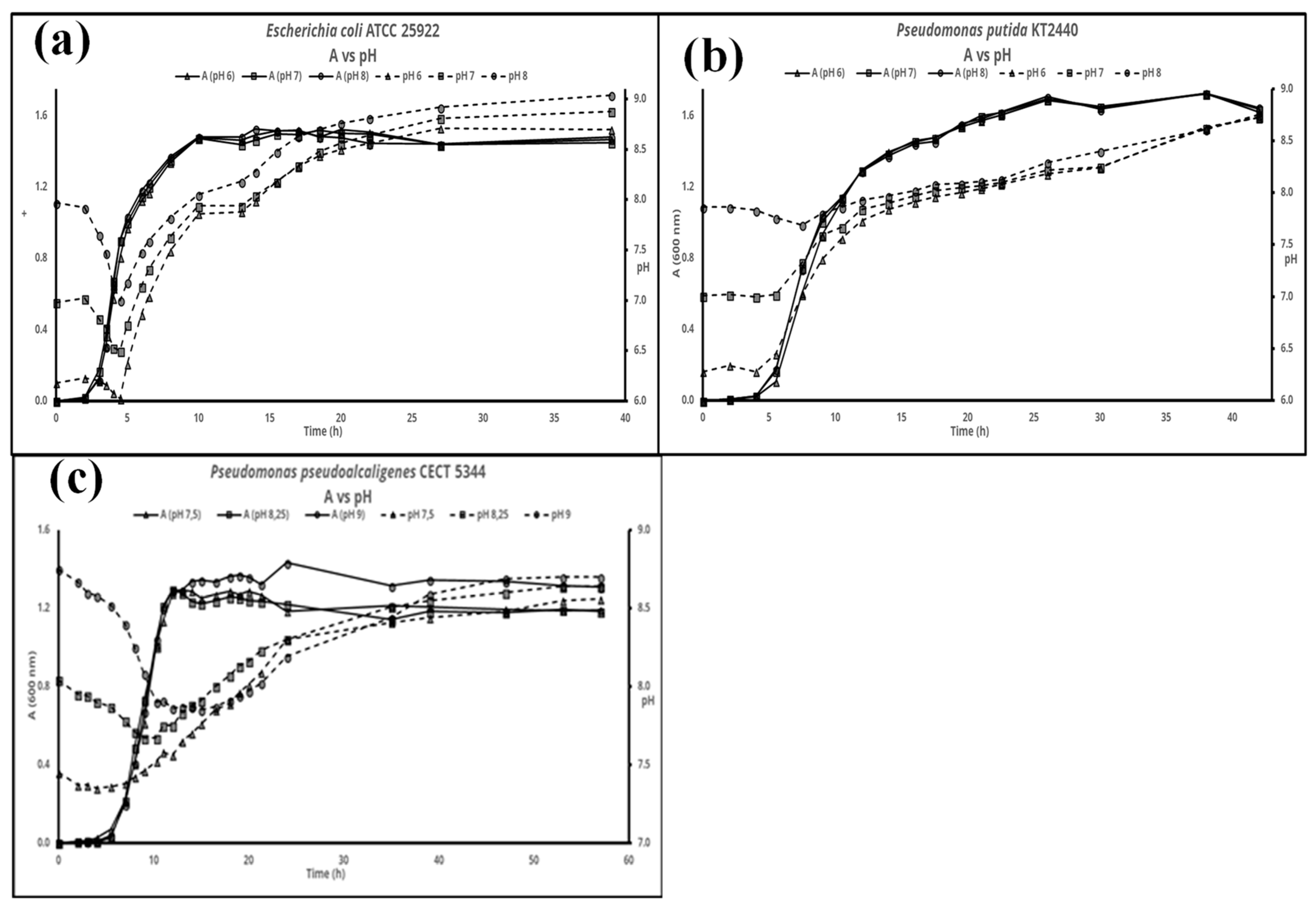{kind=link}
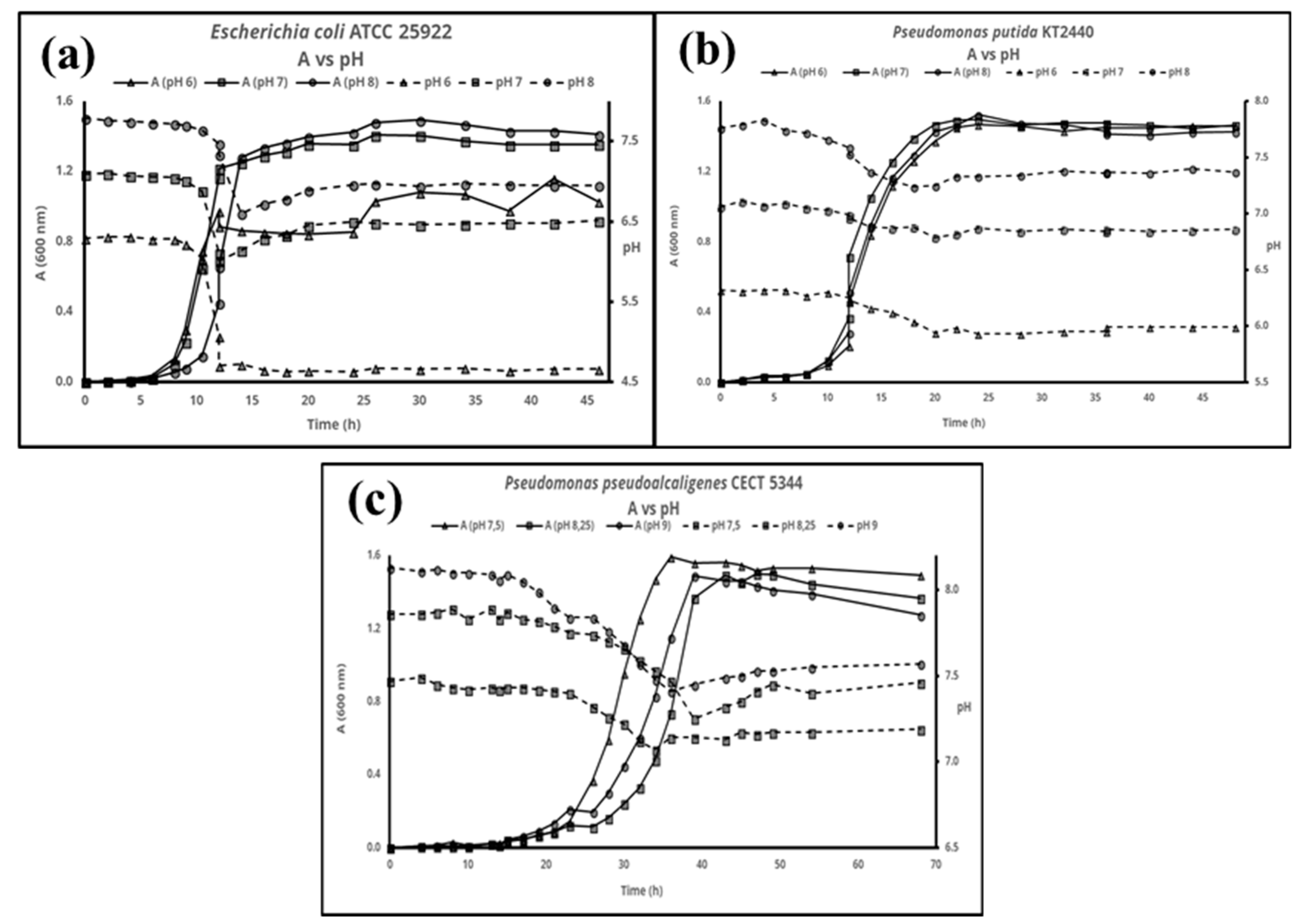{kind=link}
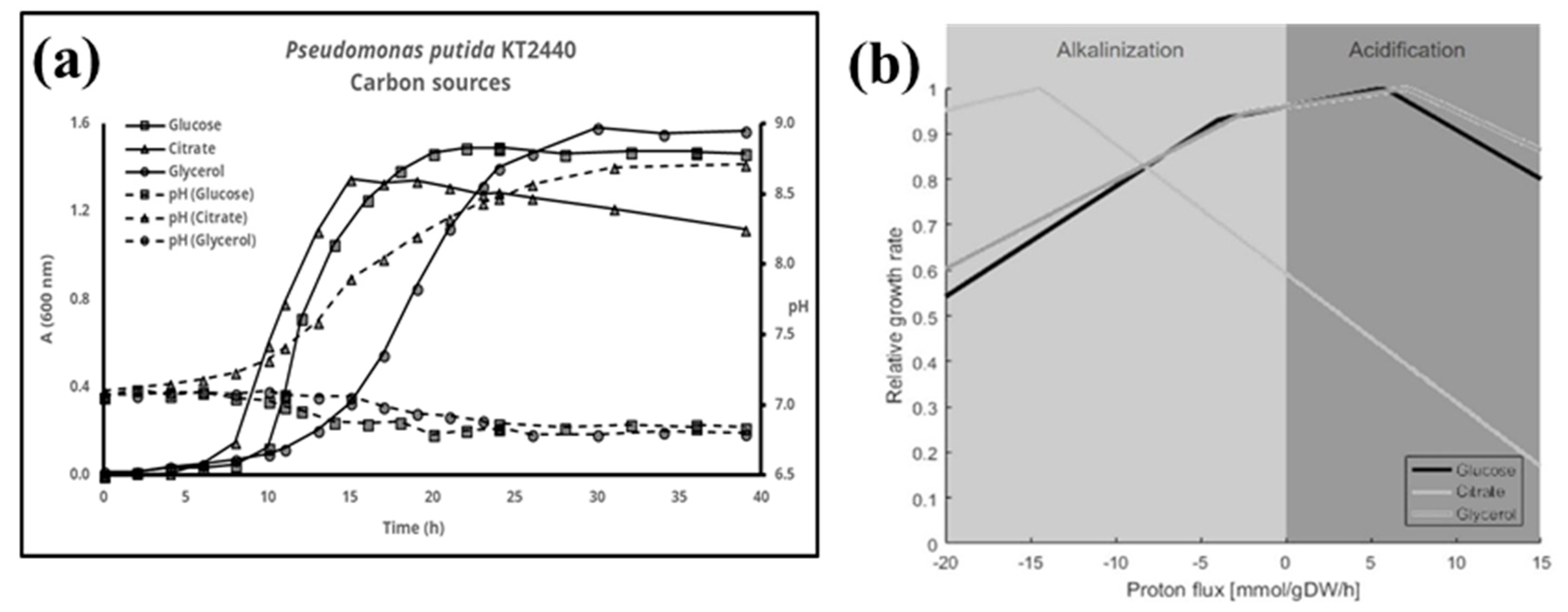{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
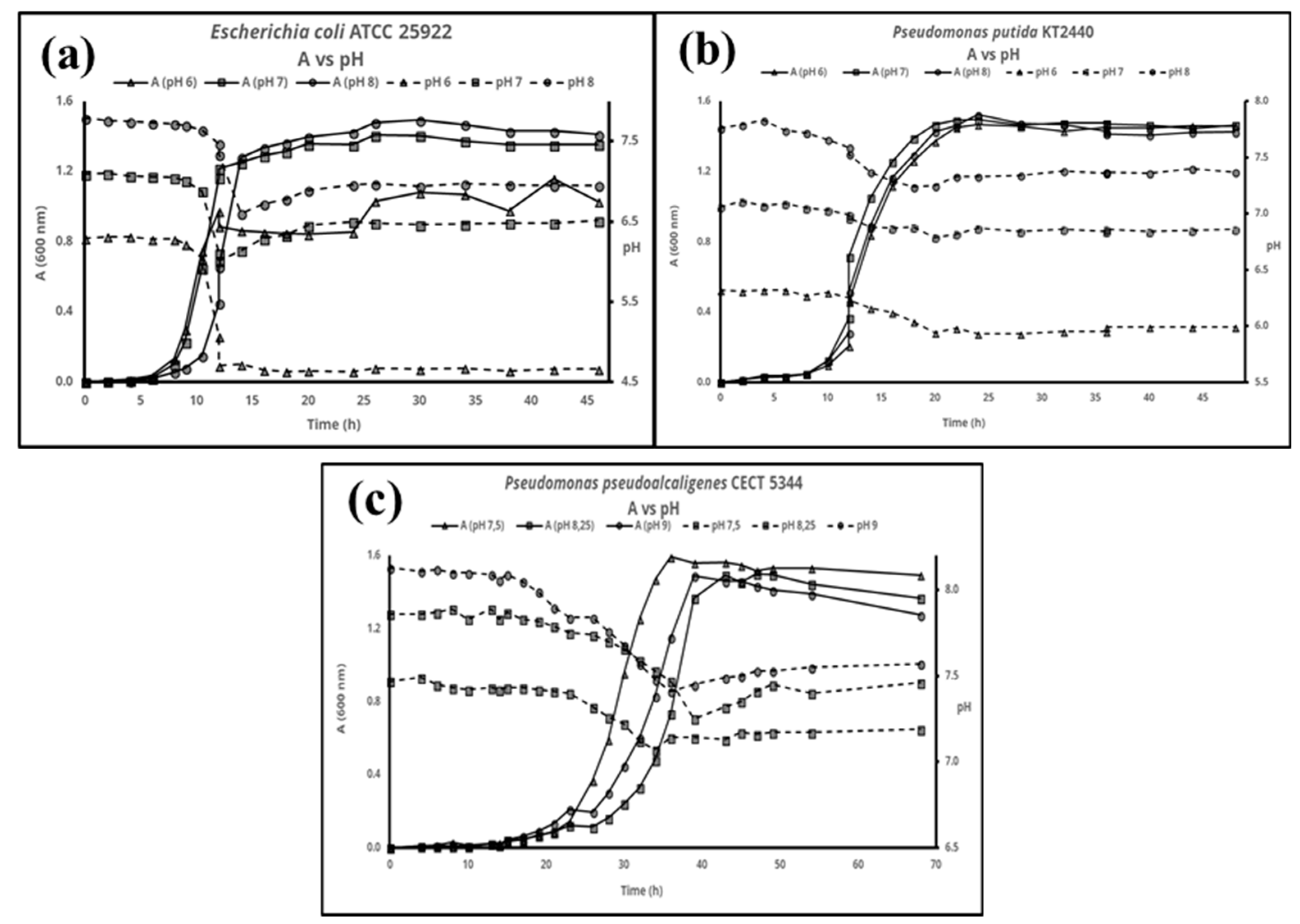
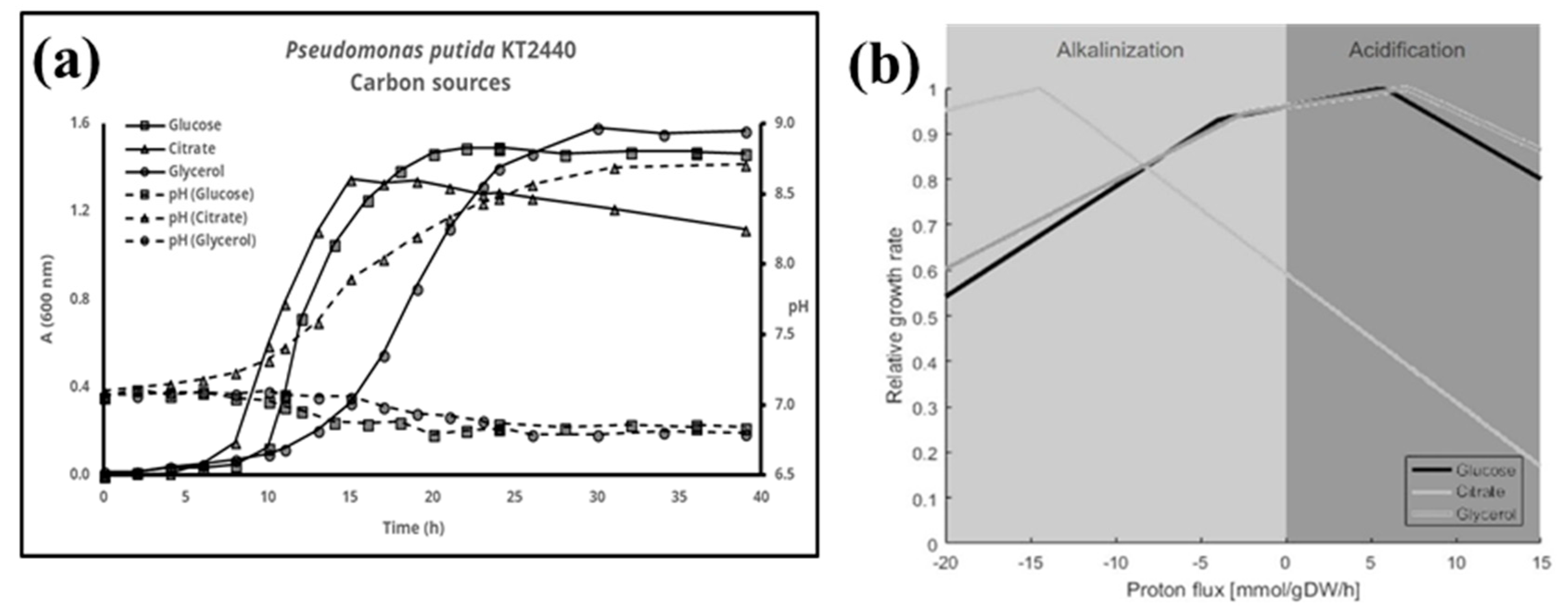
3. Results
4. Discussion
Acknowledgments
Conflicts of Interest
References
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2009, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nat. Rev. Microbiol. 2011, 9, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Ratzke, C.; Gore, J. Modifying and Reacting to the Environmental pH Drives Bacterial Interactions. bioRxiv 2017. [Google Scholar] [CrossRef]
- Nogales, J.; Gudmundsson, S.; Duque, E.; Ramos, J.L.; Palsson, B.O. Expanding The Computable Reactome In Pseudomonas putida Reveals Metabolic Cycles Providing Robustness. bioRxiv 2017. [Google Scholar] [CrossRef]
- Nogales, J.; Gudmundsson, S.; Knight, E.M.; Palsson, B.O.; Thiele, I. Detailing the optimality of photosynthesis in cyanobacteria through systems biology analysis. Proc. Natl. Acad. Sci. USA 2012, 109, 2678–2683. [Google Scholar] [CrossRef]
- Luque-Almagro, V.M.; Acera, F.; Igeño, M.I.; Wibberg, D.; Roldán, M.D.; Sáez, L.P.; Hennig, M.; Quesada, A.; Huertas, M.J.; Blom, J.; et al. Draft whole genome sequence of the cyanide-degrading bacterium Pseudomonas pseudoalcaligenes CECT5344. Environ. Microbiol. 2013, 15, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli Physiology in Luria-Bertani Broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [PubMed]
- Luli, G.W.; Strohl, W.R. Comparison of growth, acetate production, and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentations. Appl. Environ. Microbiol. 1990, 56, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Clemente, R.; Igeño, M.I.; Población, A.G.; Guijo, M.I.; Merchán, F.; Blasco, R. Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains. Proceedings 2018, 2, 1297. https://doi.org/10.3390/proceedings2201297
Sánchez-Clemente R, Igeño MI, Población AG, Guijo MI, Merchán F, Blasco R. Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains. Proceedings. 2018; 2(20):1297. https://doi.org/10.3390/proceedings2201297
Chicago/Turabian StyleSánchez-Clemente, Rubén, María Isabel Igeño, Ana G. Población, María Isabel Guijo, Faustino Merchán, and Rafael Blasco. 2018. "Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains" Proceedings 2, no. 20: 1297. https://doi.org/10.3390/proceedings2201297
APA StyleSánchez-Clemente, R., Igeño, M. I., Población, A. G., Guijo, M. I., Merchán, F., & Blasco, R. (2018). Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains. Proceedings, 2(20), 1297. https://doi.org/10.3390/proceedings2201297