
Dynamics of SPAD Index, Leaf Pigment, and Macronutrient Relationships in Sugar Beet Leaves Under Spring Nitrogen Fertilization

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Crop Management
2.2. Plant Sampling
2.3. SPAD Measurements
2.4. Pigments Determination
2.5. Macronutrients Determination
2.6. Statistical Analysis
3. Results
3.1. The ANOVA of Sugar Beet Leaf and SPAD Index
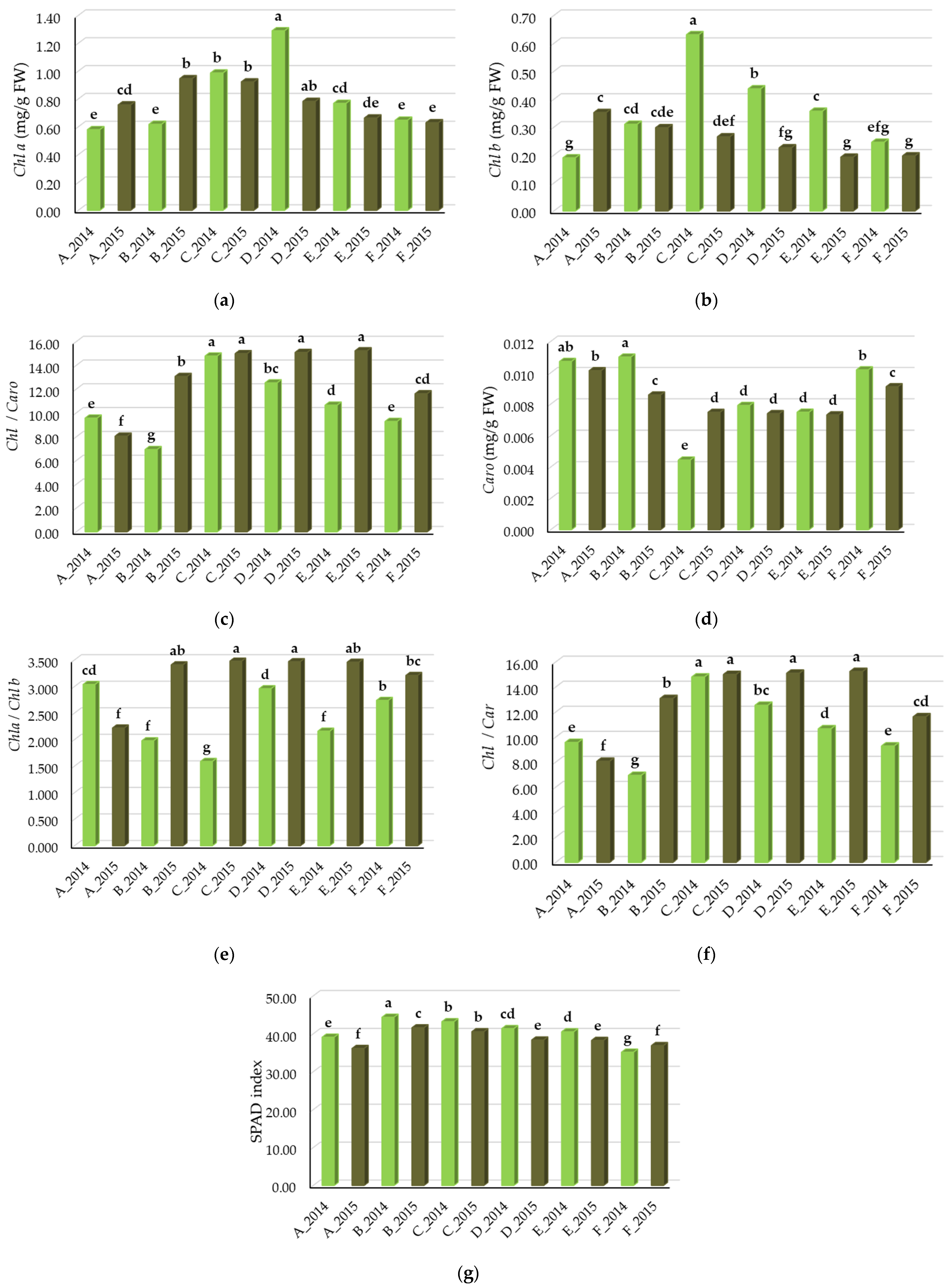
3.1.1. Leaf Pigments of the Vegetative Growth and SPAD Index
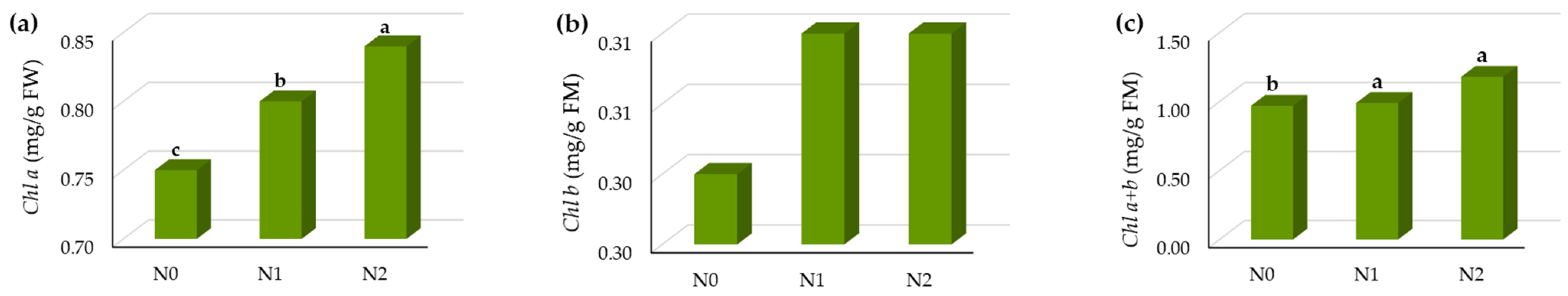
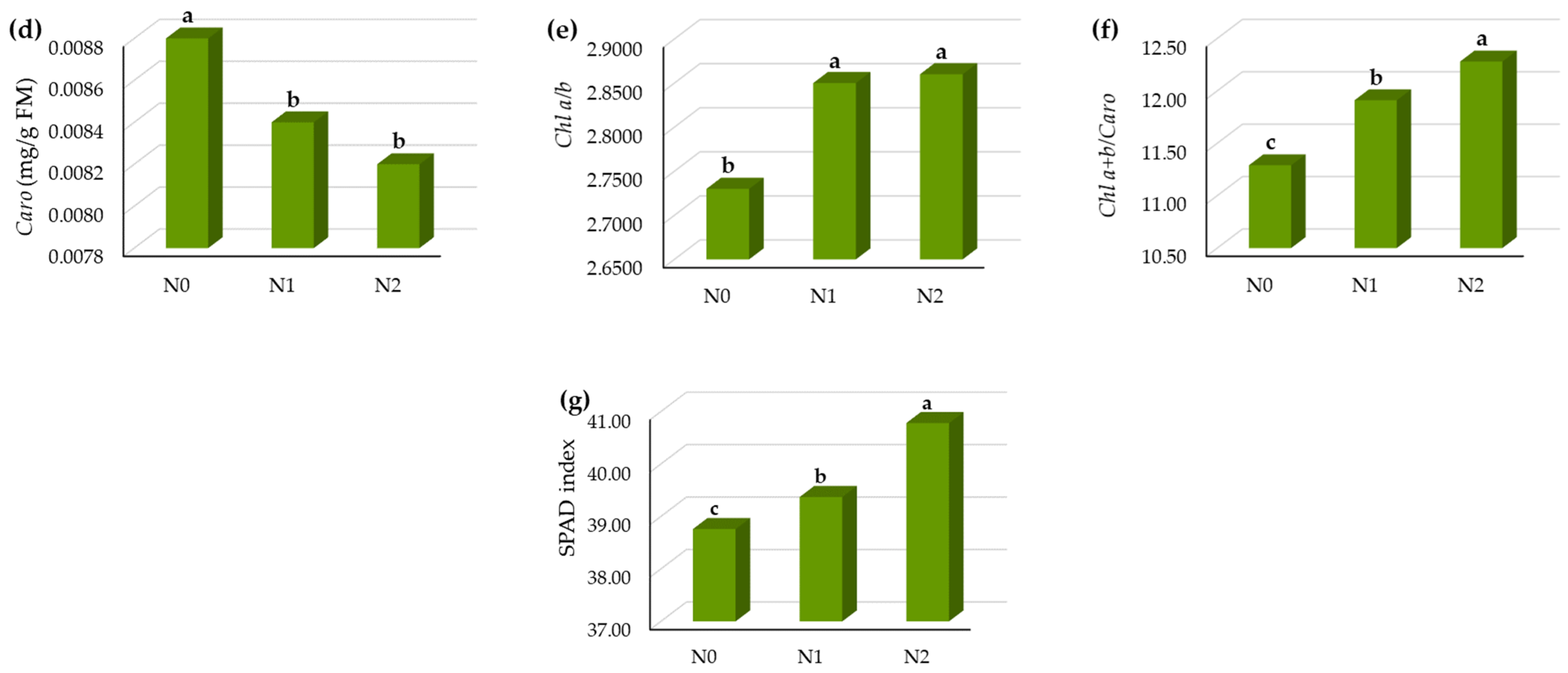
3.1.2. Influence of N Fertilization on the Leaf Pigments and SPAD Index
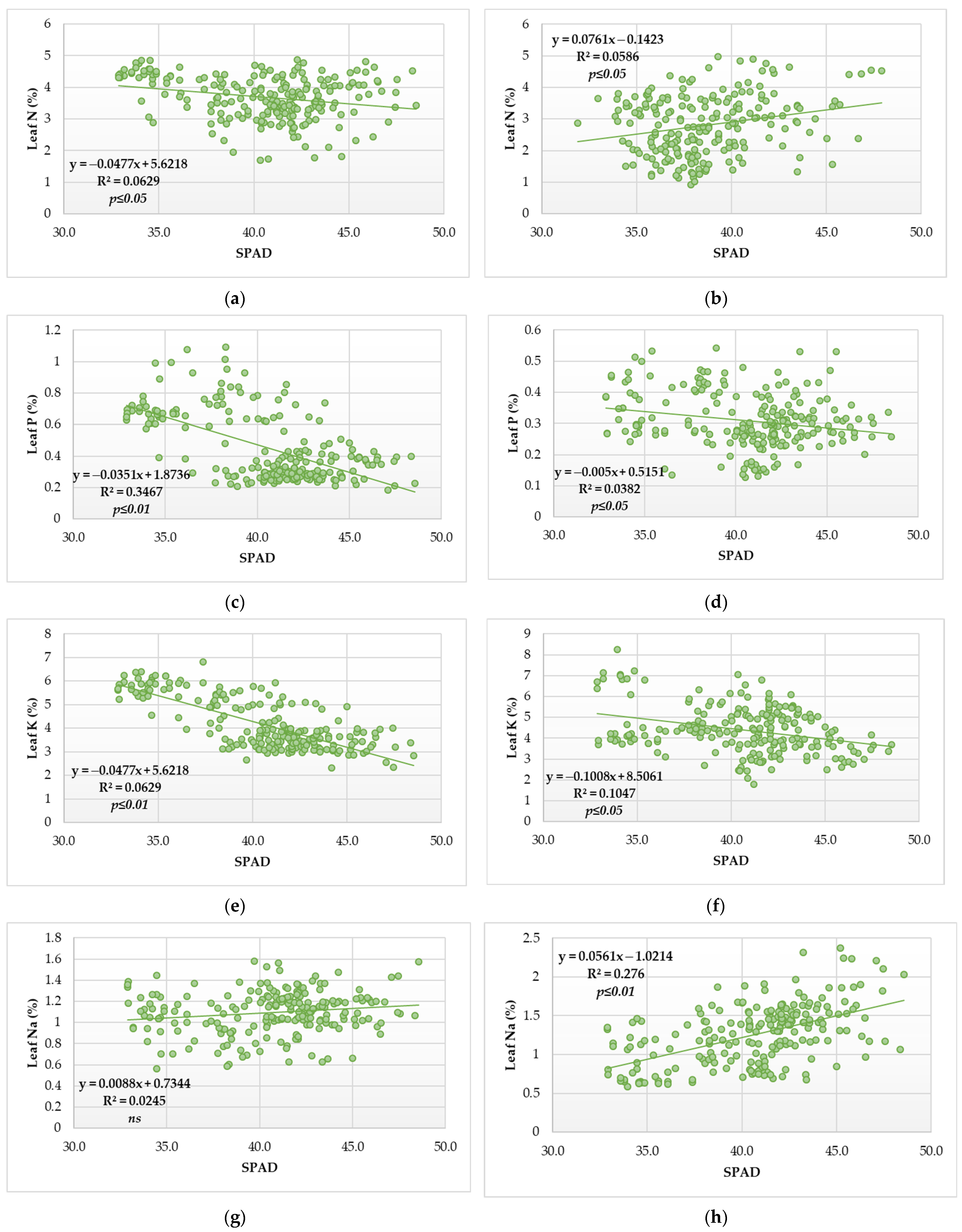
3.2. Relationship of Leaf N Status and SPAD Index
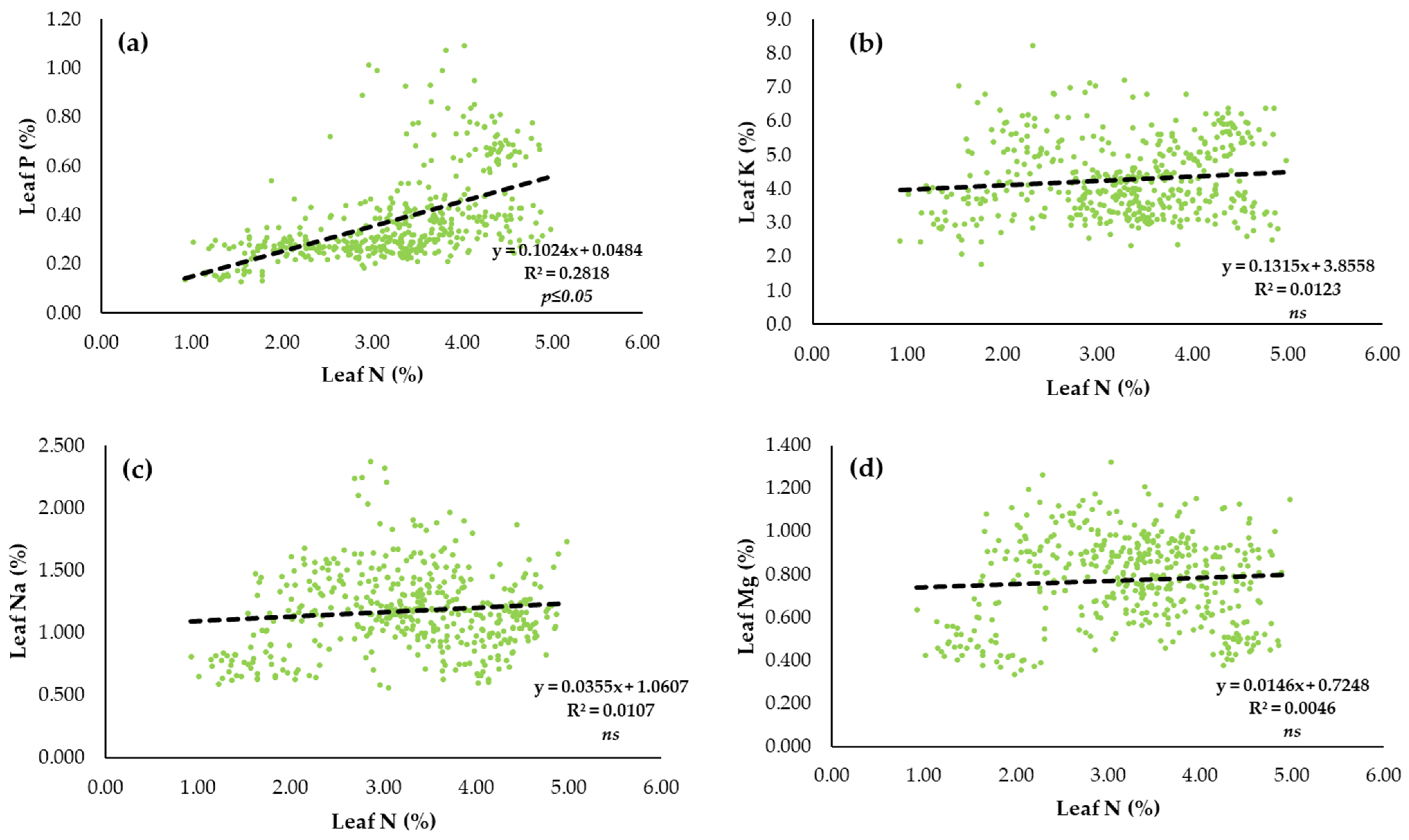
3.3. Relationship of Macronutrient Status in the Leaf and Macronutrient Status of Vegetative Growth
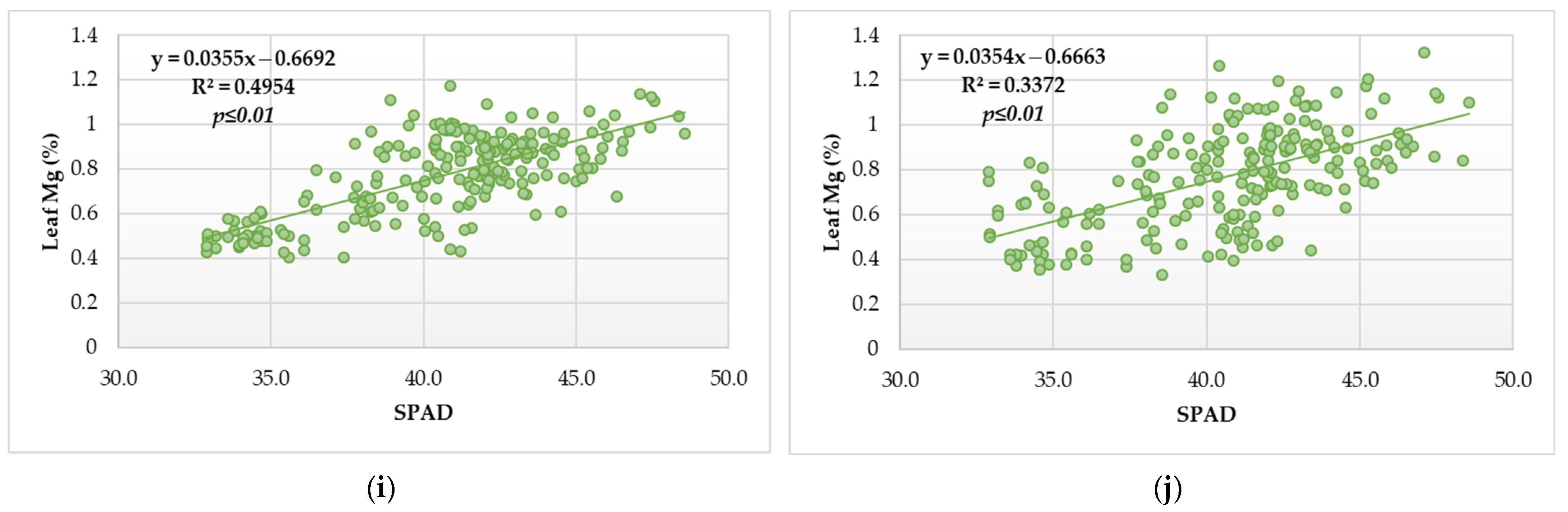
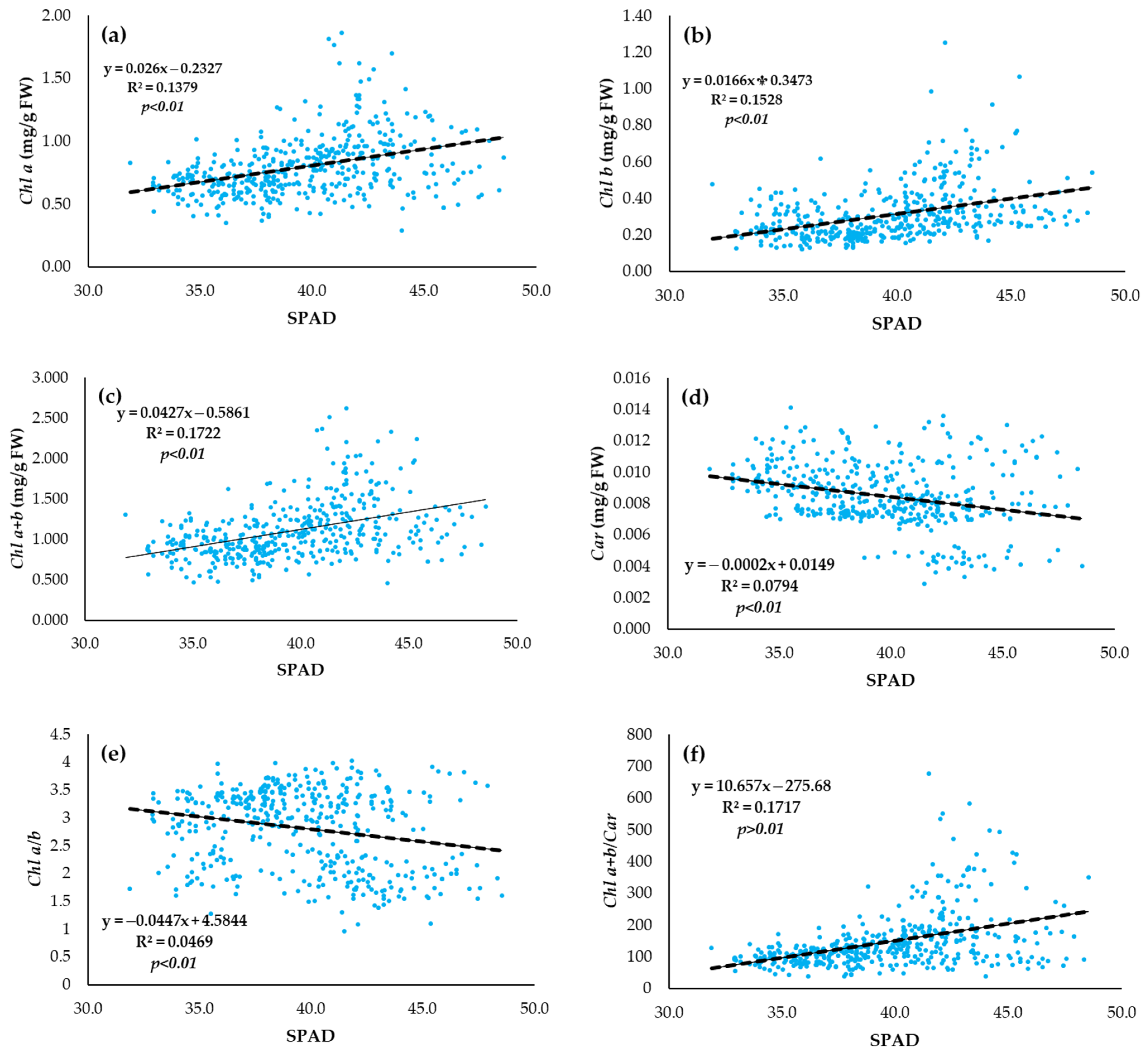
3.4. Relationship of Leaf Pigment Status and SPAD Index of Vegetative Growth
3.5. Correlation Analysis of Macronutrients Status, SPAD Index, and Leaf Pigments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kusá, H.; Mühlbachová, G.; Káš, M.; Růžek, P. Vliv organického a minerálního hnojení a plečkování na kvalitu půdy a výnos cukrové řepy. Listy Cukrov. Reparské 2024, 140, 264–270. [Google Scholar]
- Varga, I.; Kulundžić Markulj, A.; Banović, N.; Herman, G.; Iljkić, D.; Antunović, M. Sugar Beet Germination Speed and Morphometric Indicators in Relation to Temperature/Rychlost klíčení a morfometrické ukazatele cukrové řepy v závislosti na teplotě. Listy Cukrov. Řepařské 2024, 140, 326–329. [Google Scholar]
- Markulj Kulundžić, A.; Kovačević, J.; Viljevac Vuletić, M.; Josipović, A.; Liović, I.; Mijić, A.; Lepeduš, H.; Matoša Kočar, M. Impact of abiotic stress on photosynthetic efficiency and leaf temperature in sunflower. Poljoprivreda 2016, 22, 17–22. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Romanova, A.K.; Novichkova, N.S.; Vysotskaya, L.B.; Akhtyamova, Z.; Akhiyarova, G.R.; Veselov, S.Y.; Ivanov, B.N. Development of sugar beet leaves: Contents of hormones, localization of abscisic acid, and the level of products of photosynthesis. Plant Signal. Behav. 2018, 13, e1482175. [Google Scholar] [CrossRef]
- Tsialtas, J.T.; Baxevanos, D.; Maslaris, N. Chlorophyll Meter Readings, Leaf Area Index, and Their Stability as Assessments of Yield and Quality in Sugar Beet Cultivars Grown in Two Contrasting Environments. Crop Sci. 2014, 54, 265–273. [Google Scholar] [CrossRef]
- Morales, A.; Kaiser, E. Photosynthetic Acclimation to Fluctuating Irradiance in Plants. Front. Plant Sci. 2020, 11, 268. [Google Scholar]
- Markulj Kulundžić, A.; Iljkić, D.; Antunović, M.; Sudarić, A.; Varga, I. The relationship between chlorophyll a fluorescence parameters and yield components in sunflower hybrids. Bot. Serb. 2023, 47, 103–111. [Google Scholar] [CrossRef]
- Kovačević, J.; Mazur, M.; Drezner, G.; Lalić, A.; Sudarić, A.; Dvojsković, K.; Viljevac Vuletić, M.; Josipović, M.; Josipović, A.; Markulj Kulundžić, A.; et al. Photosynthetic efficiency parameters as indicators of agronomic traits of winter wheat cultivars in different soil water conditions. Genetika 2017, 49, 891–910. [Google Scholar] [CrossRef]
- Noor, H.; Ding, P.; Ren, A.; Sun, M.; Gao, Z. Effects of Nitrogen Fertilizer on Photosynthetic Characteristics and Yield. Agronomy 2023, 13, 1550. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, F.; Wang, G.; Zhang, G.; Wang, Y.; Chen, X.; Mao, Z. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. Seedlings under replant conditions. Sci. Hortic. 2014, 175, 9–15. [Google Scholar] [CrossRef]
- Draycott, A.P.; Christenson, D.R. Nutrients for Sugar Beet Production: Soil–Plant Relationships; CABI Publishing: Wallingford, UK, 2003; 242p. [Google Scholar]
- Abd El-Lateef, E.M.; Abd El-Salam, M.S.; Elewa, T.A.; Farrag, A.A. Effect of organic manure and nitrogen level on sugar beet (Beta vulgaris var. saccharifera L.) yield and root nitrate content. Am. Eurasian J. Agron. 2019, 12, 1–5. [Google Scholar]
- Leilah, A.A.A.; Khan, N. Interactive Effects of Gibberellic Acid and Nitrogen Fertilization on the Growth, Yield, and Quality of Sugar Beet. Agronomy 2021, 11, 137. [Google Scholar] [CrossRef]
- Varga, I.; Jović, J.; Rastija, M.; Markulj Kulundžić, A.; Zebec, V.; Lončarić, Z.; Iljkić, D.; Antunović, M. Efficiency and Management of Nitrogen Fertilization in Sugar Beet as Spring Crop: A Review. Nitrogen 2022, 3, 170–185. [Google Scholar] [CrossRef]
- Hadir, S.; Gaiser, T.; Hüging, H.; Athmann, M.; Pfarr, D.; Kemper, R.; Ewert, F.; Seidel, S. Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission. Agriculture 2021, 11, 21. [Google Scholar] [CrossRef]
- Liu, X.; Lu, Z.; Yao, Q.; Xu, L.; Fu, J.; Yin, X.; Bai, Q.; Liu, D.; Xing, W. MicroRNAs Participate in Morphological Acclimation of Sugar Beet Roots to Nitrogen Deficiency. Int. J. Mol. Sci. 2024, 25, 9027. [Google Scholar] [CrossRef] [PubMed]
- Salami, M.; Saadat, S. Study of potassium and nitrogen fertilizer levels on the yield of sugar beet in Jolge cultivar. J. Novel Appl. Sci. 2013, 2, 94–100. [Google Scholar]
- Fueki, N.; Sato, K.; Takeuchi, H.; Sato, H.; Nakatsu, S.; Kato, J. Prediction of nitrogen uptake by sugar beet (Beta vulgaris L.) by scoring organic matter and nitrogen management (N-score), in Hokkaido, Japan. Soil. Sci. Plant Nutr. 2011, 57, 411–420. [Google Scholar] [CrossRef]
- Wang, N.; Fu, F.; Wang, H.; Wang, P.; He, S.; Shao, H.; Ni, Z.; Zhang, X. Effects of irrigation and nitrogen on chlorophyll content, dry matter and nitrogen accumulation in sugar beet (Beta vulgaris L.). Sci. Rep. 2021, 11, 16651. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, H.; Wang, D.; Li, H.; Mouazen, A.M. A novel spectral index for estimation of relative chlorophyll content of sugar beet. Comput. Electron. Agric. 2021, 184, 106088. [Google Scholar] [CrossRef]
- Villaño, D.; García-Viguera, C.; Mena, P. Colors: Health Effects. In The Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 2, pp. 265–272. [Google Scholar]
- Yuan, Z.; Cao, Q.; Zhang, K.; Ata-Ul-Karim, S.T.; Tian, Y.; Zhu, Y.; Cao, W.; Liu, X. Optimal Leaf Positions for SPAD Meter Measurement in Rice. Front. Plant Sci. 2016, 7, 719. [Google Scholar] [CrossRef] [PubMed]
- Campos, W.V.; Filho, J.T.; Filho, J.; São José, A.R. Regression equation for correlation between Spad index and photosynthetic pigments under saline stress in African mahogany culture. Int. J. Sci. Res. Arch. 2023, 8, 647–655. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, X.; Nawaz, G.; Yin, J.; Yang, J. Physiological and Biochemical Responses of four cassava cultivars to drought stress. Sci. Rep. 2020, 10, 6968. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Meng, J.; Tao, J.; Zhao, D. Osmotic Regulation, Antioxidant Enzyme Activities and Photosynthetic Characteristics of Tree Peony (Paeonia suffruticosa Andr.) in Response to High-Temperature Stress. Phyton-Int. J. Exp. Bot. 2023, 92, 3133–3147. [Google Scholar] [CrossRef]
- Wang, A.; Song, Z.; Xie, Y.; Hu, J.; Zhang, L.; Zhu, Q. Detection of Rice Leaf SPAD and Blast Disease Using Integrated Aerial and Ground Multiscale Canopy Reflectance Spectroscopy. Agriculture 2024, 14, 1471. [Google Scholar] [CrossRef]
- Radočaj, D.; Rapčan, I.; Jurišić, M. Indoor Plant Soil-Plant Analysis Development (SPAD) Prediction Based on Multispectral Indices and Soil Electroconductivity: A Deep Learning Approach. Horticulturae 2023, 9, 1290. [Google Scholar] [CrossRef]
- Monostori, I.; Árendás, T.; Hoffman, B.; Galiba, G.; Gierczik, K.; Szira, F.; Vágújfalvi, A. Relationship between SPAD value and grain yield can be affected by cultivar, environment and soil nitrogen content in wheat. Euphytica 2016, 211, 103–112. [Google Scholar] [CrossRef]
- Markulj Kulundžić, A.; Viljevac Vuletić, M.; Matoša Kočar, M.; Antunović Dunić, J.; Varga, I.; Zdunić, Z.; Sudarić, A.; Cesar, V.; Lepeduš, H. Effect of Elevated Temperature and Excess Light on Photosynthetic Efficiency, Pigments, and Proteins in the Field-Grown Sunflower during Afternoon. Horticulturae 2022, 8, 392. [Google Scholar] [CrossRef]
- Szulc, P.; Bocianowski, J.; Nowosad, K.; Zielewicz, W.; Kobus-Cisowska, J. SPAD Leaf Greenness Index: Green Mass Yield Indicator of Maize (Zea mays L.), Genetic and Agriculture Practice Relationship. Plants 2021, 10, 830. [Google Scholar] [CrossRef]
- Gao, W.; Zeng, W.; Li, S.; Zhang, L.; Wang, W.; Song, J.; Wu, H. Remote sensing estimation of sugar beet SPAD based on un-manned aerial vehicle multispectral imagery. PLoS ONE 2024, 19, e0300056. [Google Scholar] [CrossRef] [PubMed]
- Kun, A.; Kolozsvári, I.; Jancsó, M.; Túri, N.; Bozán, C. Effect of irrigation and water quality on the physiological status of sugar beet and fodder beet using SPAD-502 chlorophyll meter. Columella–J. Agric. Environ. Sci. 2022, 9, 2. [Google Scholar] [CrossRef]
- Mielke, M.S.; Schaffer, B.; LI, C. Use of a SPAD meter to estimate chlorophyll content in Eugenia uniflora L. leaves as affected by contrasting light environments and soil flooding. Photosynthetica 2010, 48, 332–338. [Google Scholar] [CrossRef]
- Verdenal, T.; Zufferey, V.; Reynard, J.S.; Spring, J.L. Nitrogen nutrition status of the vine: Correlation between N-tester and SPAD chlorophyll indices: This is a translation of an article originally written in French. IVES Tech. Rev. Vine Wine 2023. [Google Scholar] [CrossRef]
- Tsialtas, J.T.; Maslaris, N. Sugar beet response to N fertilization as assessed by late season chlorophyll and leaf area index measurements in a semi-arid environment. Int. J. Plant Prod. 2008, 2, 57–70. [Google Scholar]
- Lentz, R.D.; Lehrsch, G.A. Nitrogen availability and uptake by sugarbeet in years following a manure application. Int. J. Agron. 2012, 2012, 120429. [Google Scholar] [CrossRef]
- Ebmeyer, H.; Hoffmann, C.M. Efficiency of nitrogen uptake and utilization in sugar beet genotypes. Field Crops Res. 2021, 274, 108334. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Xu, P.; Liu, D.; Zhang, L.; Hao, Y.; Wang, K.; Fan, H. Nitrogen use efficiency of drip irrigated sugar beet as affected by sub-optimal levels of nitrogen and irrigation. Agric. Water Manag. 2024, 298, 108849. [Google Scholar] [CrossRef]
- Monostori Zhang, R.; Yang, P.; Liu, S.; Wang, C.; Liu, J. Evaluation of the Methods for Estimating Leaf Chlorophyll Content with SPAD Chlorophyll Meters. Remote Sens. 2022, 14, 5144. [Google Scholar] [CrossRef]
- Meskinivishkaee, F.; Mohammadi, M.H.; Neyshabouri, M.R.; Shekari, F. Evaluation of canola chlorophyll index and leaf nitrogen under a wide range of soil moisture. Int. Agrophys. 2015, 29, 83–90. [Google Scholar] [CrossRef]
- Varga, I.; Lončarić, Z.; Kristek, S.; Kulundžić, A.M.; Rebekić, A.; Antunović, M. Sugar Beet Root Yield and Quality with Leaf Seasonal Dynamics in Relation to Planting Densities and Nitrogen Fertilization. Agriculture 2021, 11, 407. [Google Scholar] [CrossRef]
- State Hydrometeorological Service. Available online: https://meteo.hr/ (accessed on 11 November 2020).
- Holm, G. Chlorophyll mutations in barley. Acta Agric. Scand. 1954, 4, 457–471. [Google Scholar] [CrossRef]
- Wettstein, D. Chlorophyll letale und der submikroskopische Formwechsel der Plastiden. Exp. Cell Res. 1957, 12, 427–434. [Google Scholar] [CrossRef] [PubMed]
- SAS®. STAT 9.3; SAS Institute Inc.: Cary, NC, USA, 2024. [Google Scholar]
- Maslova, T.G.; Markovskaya, E.F.; Slemnev, N.N. Functions of carotenoids in leaves of higher plants. Biol. Bull. Rev. 2021, 11, 476–487. [Google Scholar] [CrossRef]
- Elsayed, S.; El-Hendawy, S.; Elsherbiny, O.; Okasha, A.M.; Elmetwalli, A.H.; Elwakeel, A.E.; Memon, M.S.; Ibrahim, M.E.M.; Ibrahim, H.H. Estimating Chlorophyll Content, Production, and Quality of Sugar Beet under Various Nitrogen Levels Using Machine Learning Models and Novel Spectral Indices. Agronomy 2023, 13, 2743. [Google Scholar] [CrossRef]
- Salaić, M.; Galić, V.; Jambrović, A.; Zdunić, Z.; Šimić, D.; Brkić, A.; Petrović, S. Assessing Genetic Variability for NUE in Maize Lines from Agricultural Institute Osijek. Poljoprivreda 2024, 30, 13–20. [Google Scholar] [CrossRef]
- Radočaj, D.; Tuno, N.; Mulahusić, A.; Jurišić, M. Evaluacija kombinacije strojnog učenja za geoprostorno predviđanje sadržaja željeza u tlu u Hrvatskoj. Poljoprivreda 2023, 29, 53–61. [Google Scholar] [CrossRef]
- Varga, I.; Markulj Kulundžić, A.; Tkalec Kojić, M.; Antunović, M. Does the Amount of Pre-Sowing Nitrogen Fertilization Affect Sugar Beet Root Yield and Quality of Different Genotypes? Nitrogen 2024, 5, 386–408. [Google Scholar] [CrossRef]
- Bergmann, W. Nutritional Disorders of Plants–Development, Visual and Analytical Diagnosis; VCH Publishers Inc.: Hoboken, NJ, USA, 1992. [Google Scholar]
- Draycott, A.P. Sugar Beet; Blackwell Publishing Ltd.: Oxford, UK, 2006. [Google Scholar]
- Pogłodziński, R.; Barłóg, P.; Grzebisz, W. Effect of nitrogen and magnesium sulfate application on sugar beet yield and quality. Plant Soil. Environ. 2021, 67, 507–513. [Google Scholar] [CrossRef]
- Grzegorzewski, K.; Ciećko, Z.; Szostek, R. Influence of mineral fertilization on the yield and macroelement content in sugar beet. Acta Agroph. 2017, 24, 221–237. [Google Scholar]
- Zewail, R.M.Y.; El-Gmal, I.S.; Khaitov, B.; El-Desouky, H.S.A. Micronutrients through foliar application enhance growth, yield, and quality of sugar beet (Beta vulgaris L.). J. Plant Nutr. 2020, 43, 2275–2285. [Google Scholar] [CrossRef]
- Varga, I.; Iljkić, D.; Krolo, P.; Perić Fekete, A.; Kraus, I. The Source of K Fertilizer for Industrial Hemp (Cannabis sativa L.): Mechanical and Chemical Properties of Stem for Rammed Earth Walls. Agriculture 2024, 14, 2196. [Google Scholar] [CrossRef]
- Füllgrabe, H.; Claassen, N.; Hilmer, R.; Koch, H.J.; Dittert, K.; Kreszies, T. Potassium deficiency reduces sugar yield in sugar beet through decreased growth of young plants. J. Plant Nutr. Soil. Sci. 2022, 185, 545–553. [Google Scholar] [CrossRef]
- Enan, S.A.A.M. Response of sugar beet to different levels of potassium and magnesium fertilization under sandy soil conditions. J. Plant Prod. 2016, 7, 963–972. [Google Scholar] [CrossRef]
- Ishfaq, M.; Wang, Y.; Yan, M.; Wang, Z.; Wu, L.; Li, C.; Li, X. Physiological essence of magnesium in plants and its widespread deficiency in the farming system of China. Front. Plant Sci. 2022, 13, 802274. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Johnson, G.N.; Strasser, R.J.; Verbruggen, N. Physiological characterisation of magnesium deficiency in sugar beet: Acclimation to low magnesium differentially affects photosystems I and II. Planta 2004, 220, 344–355. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chl a | Chl b | Chl a + b | Caro | Chl a/Chl b | Chl a + b/Car | SPAD | |
|---|---|---|---|---|---|---|---|
| Y | ** | <0.001 | <0.001 | ** | <0.001 | <0.001 | <0.001 |
| D | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| N | <0.001 | ns | ** | <0.001 | ** | <0.001 | <0.001 |
| Y × D | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Y × N | ns | ns | ns | ns | ns | ns | ns |
| N × D | ns | ** | <0.001 | <0.001 | <0.001 | * | |
| Y × D × N | <0.001 | ** | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| N | P | K | Na | Mg | SPAD | Chl a | Chl b | Chl a + b | Caro | Chl a/b | Chl/Caro | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | 1 | 0.531 *** | 0.111 * | 0.103 * | 0.067 ns | 0.144 ** | −0.034 ns | 0.103 * | 0.020 ns | 0.356 *** | −0.281 *** | −0.079 ns |
| P | 1 | 0.568 *** | 0.313 *** | −0.383 *** | −0.306 *** | −0.393 *** | −0.261 *** | −0.374 *** | 0.595 *** | −0.054 ns | −0.401 *** | |
| K | 1 | 0.010 ns | −0.206 *** | −0.543 *** | −0.425 *** | −0.461 *** | −0.481 *** | 0.323 *** | 0.276 *** | −0.441 *** | ||
| Na | 1 | 0.647 *** | 0.141 ** | 0.219 *** | 0.060 ns | 0.174 ** | −0.277 *** | 0.144 ** | 0.160 *** | |||
| Mg | 1 | 0.520 *** | 0.338 *** | 0.224 *** | 0.322 *** | −0.398 *** | −0.011 ns | 0.326 *** | ||||
| SPAD | 1 | 0.371 *** | 0.391 *** | 0.415 *** | −0.282 *** | −0.217 *** | 0.414 *** | |||||
| Chl a | 1 | 0.650 *** | 0.949 *** | −0.473 *** | 0.022 ns | 0.705 *** | ||||||
| Chl b | 1 | 0.855 *** | −0.478 *** | −0.666 *** | ||||||||
| Chl a + b | 1 | −0.518 *** | −0.260 *** | 0.836 *** | ||||||||
| Caro | 1 | 0.031 ns | −0.774 *** | |||||||||
| Chl a/b | 1 | 0.863 *** | ||||||||||
| Chl/Caro | 1 | |||||||||||
| * p ≤ 0.05 | ** p ≤ 0.01 | *** p ≤ 0.001 | ns—non significant | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, I.; Pospišil, M.; Iljkić, D.; Markulj Kulundžić, A.; Tkalec Kojić, M.; Antunović, M. Dynamics of SPAD Index, Leaf Pigment, and Macronutrient Relationships in Sugar Beet Leaves Under Spring Nitrogen Fertilization. Nitrogen 2025, 6, 10. https://doi.org/10.3390/nitrogen6010010
Varga I, Pospišil M, Iljkić D, Markulj Kulundžić A, Tkalec Kojić M, Antunović M. Dynamics of SPAD Index, Leaf Pigment, and Macronutrient Relationships in Sugar Beet Leaves Under Spring Nitrogen Fertilization. Nitrogen. 2025; 6(1):10. https://doi.org/10.3390/nitrogen6010010
Chicago/Turabian StyleVarga, Ivana, Milan Pospišil, Dario Iljkić, Antonela Markulj Kulundžić, Monika Tkalec Kojić, and Manda Antunović. 2025. "Dynamics of SPAD Index, Leaf Pigment, and Macronutrient Relationships in Sugar Beet Leaves Under Spring Nitrogen Fertilization" Nitrogen 6, no. 1: 10. https://doi.org/10.3390/nitrogen6010010
APA StyleVarga, I., Pospišil, M., Iljkić, D., Markulj Kulundžić, A., Tkalec Kojić, M., & Antunović, M. (2025). Dynamics of SPAD Index, Leaf Pigment, and Macronutrient Relationships in Sugar Beet Leaves Under Spring Nitrogen Fertilization. Nitrogen, 6(1), 10. https://doi.org/10.3390/nitrogen6010010