
Simulation Study on Dose and LET of Neutron Irradiation for Biological Experiments Using Spallation, Reactor, and Compact Neutron Sources

 ,
,
Abstract
1. Introduction
2. Method and Results
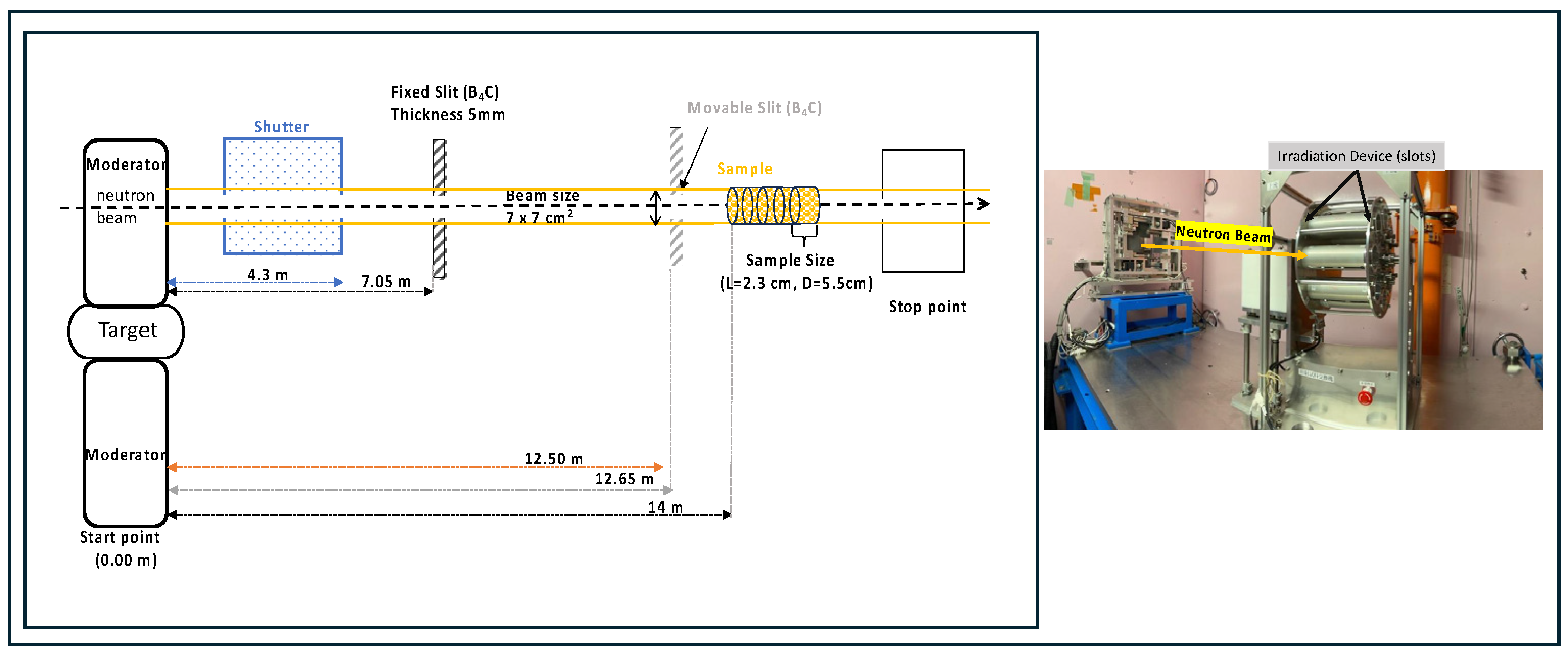
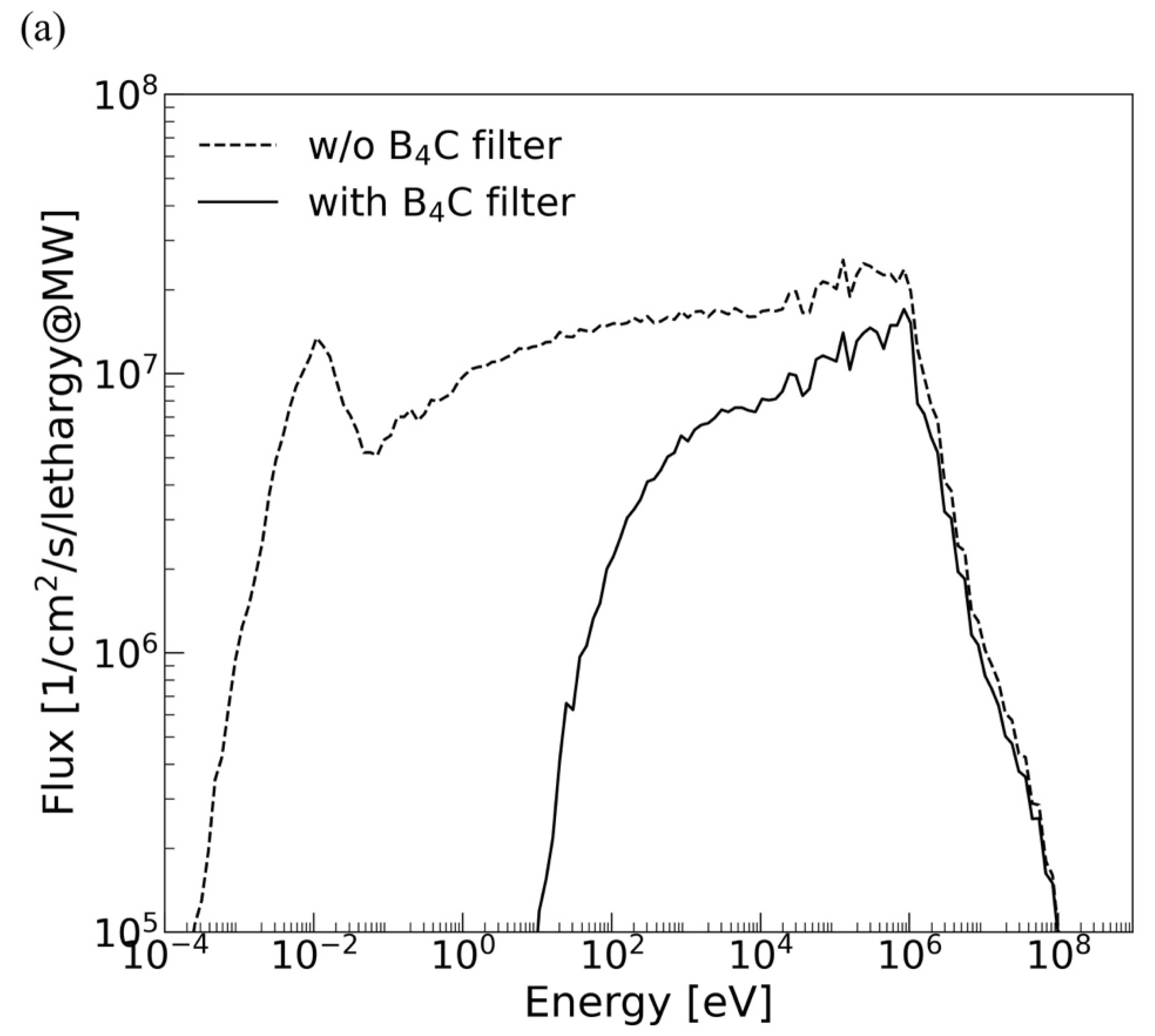
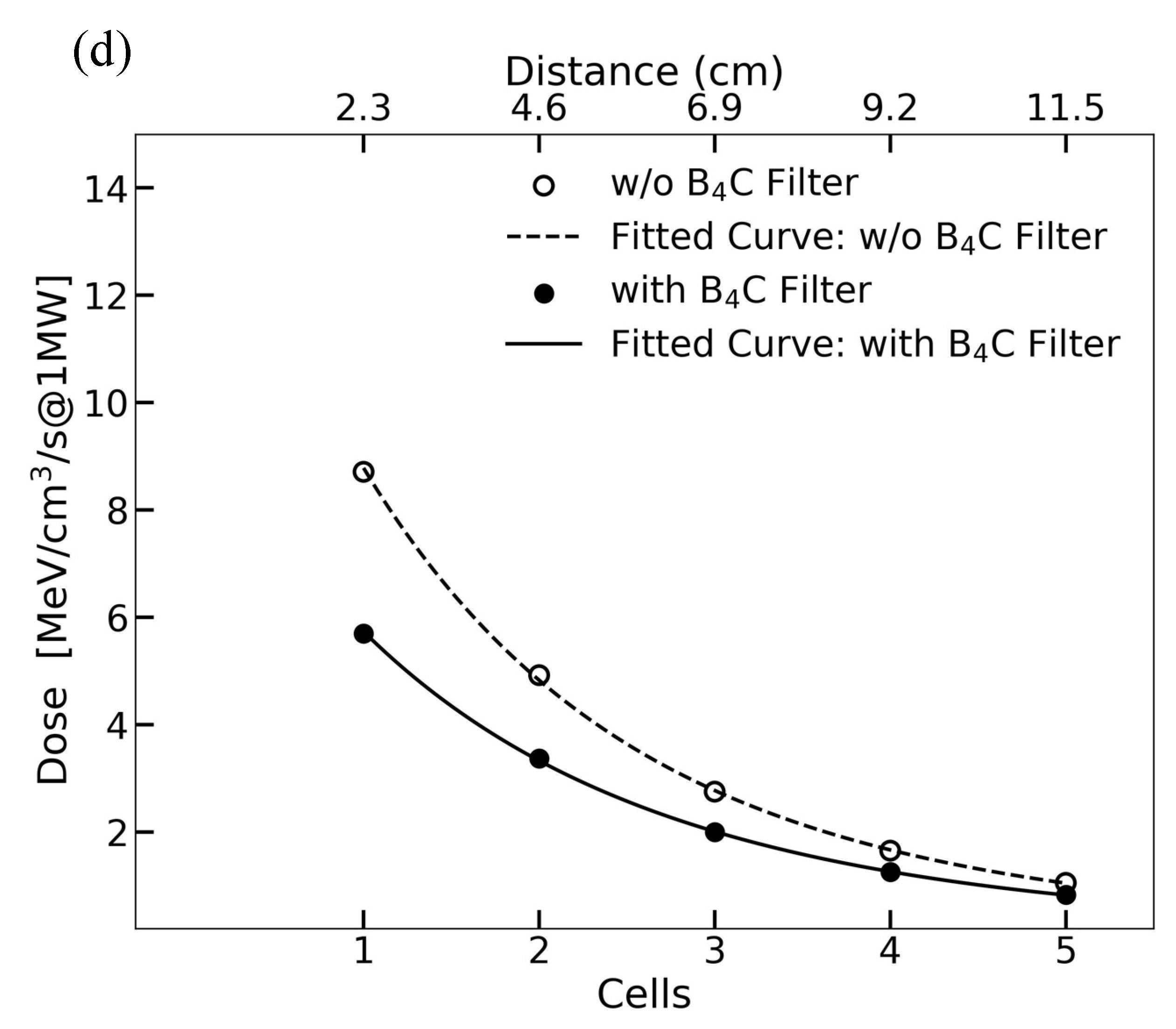
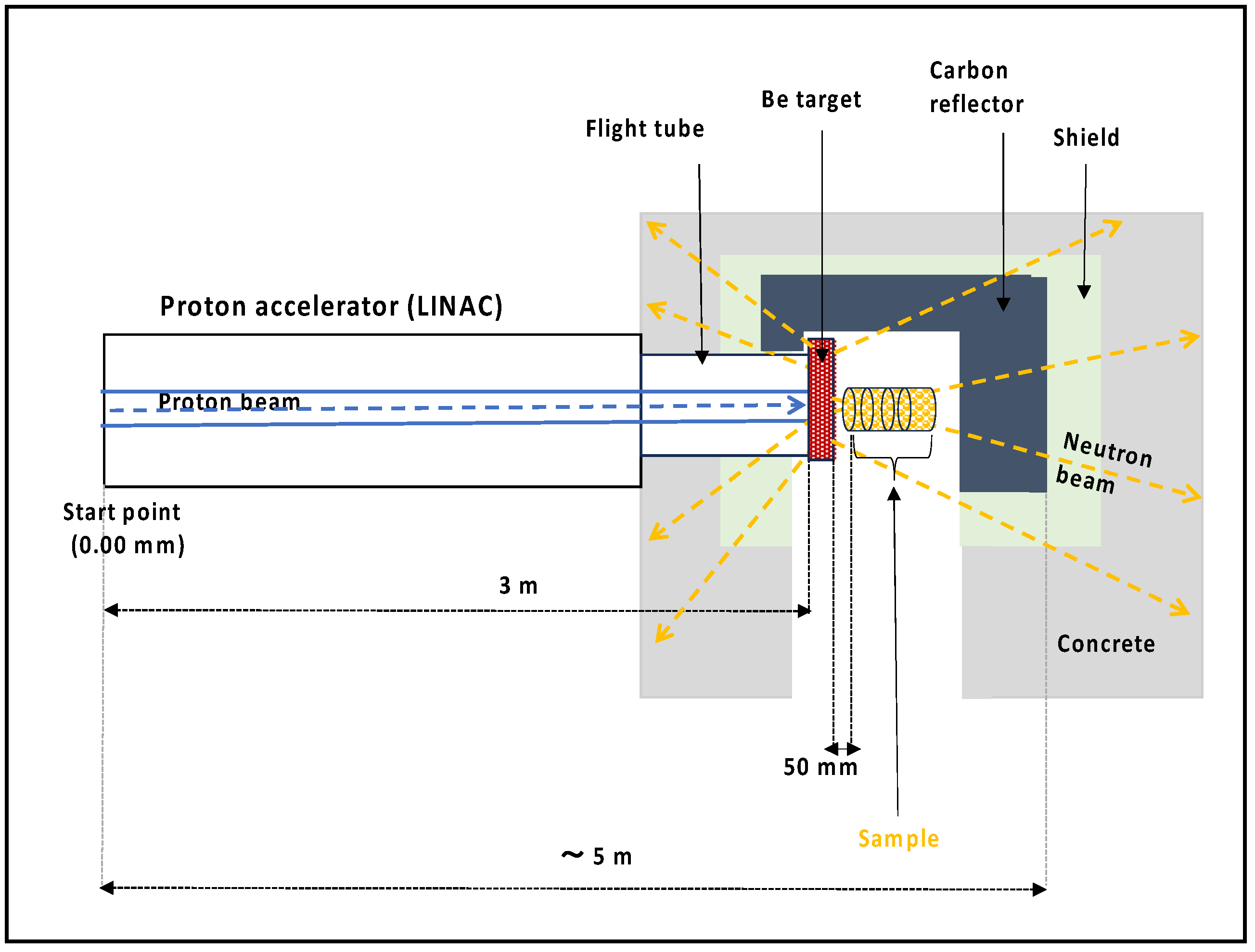
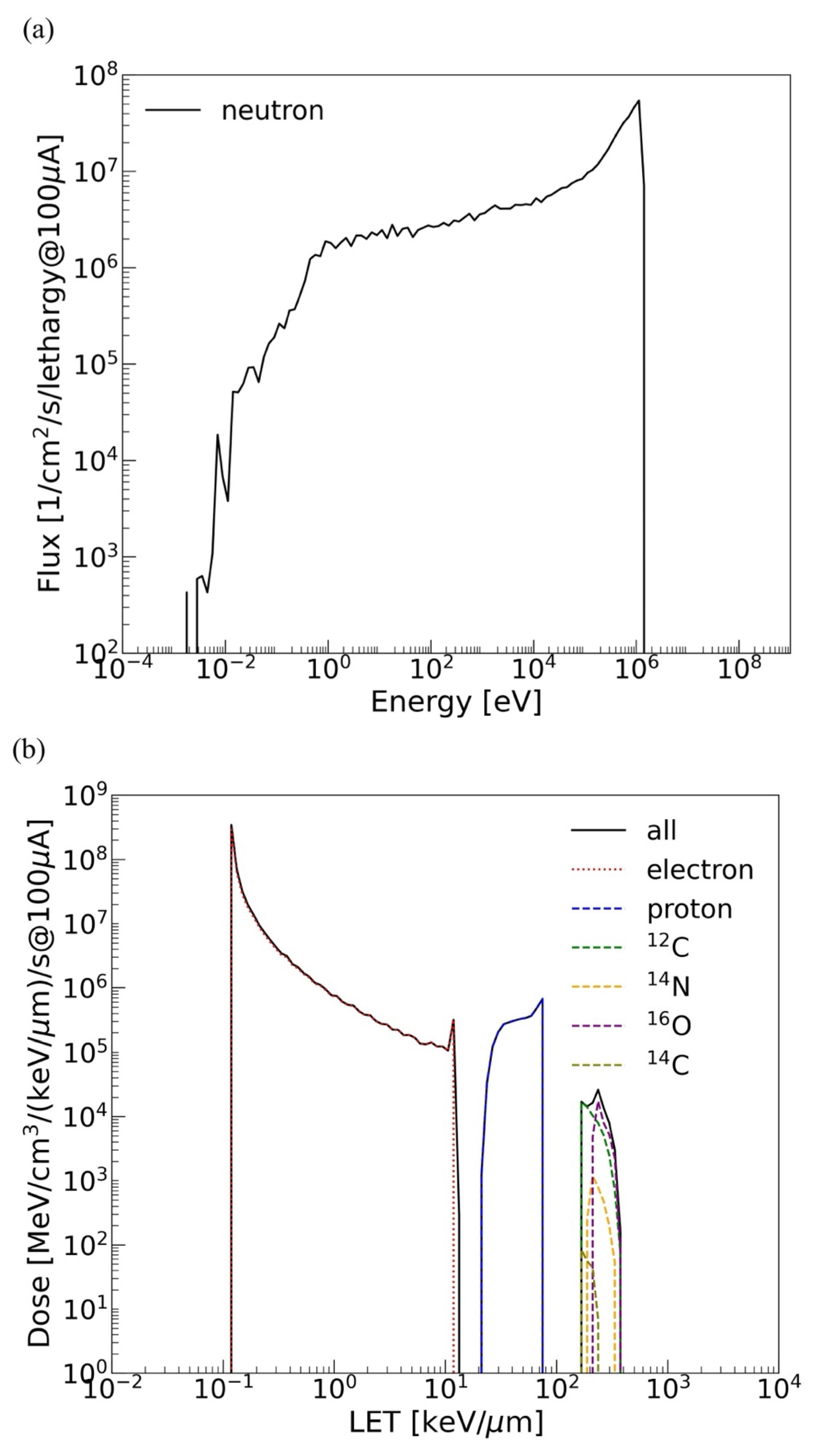
2.1. Irradiation Setup at J-PARC and Simulation
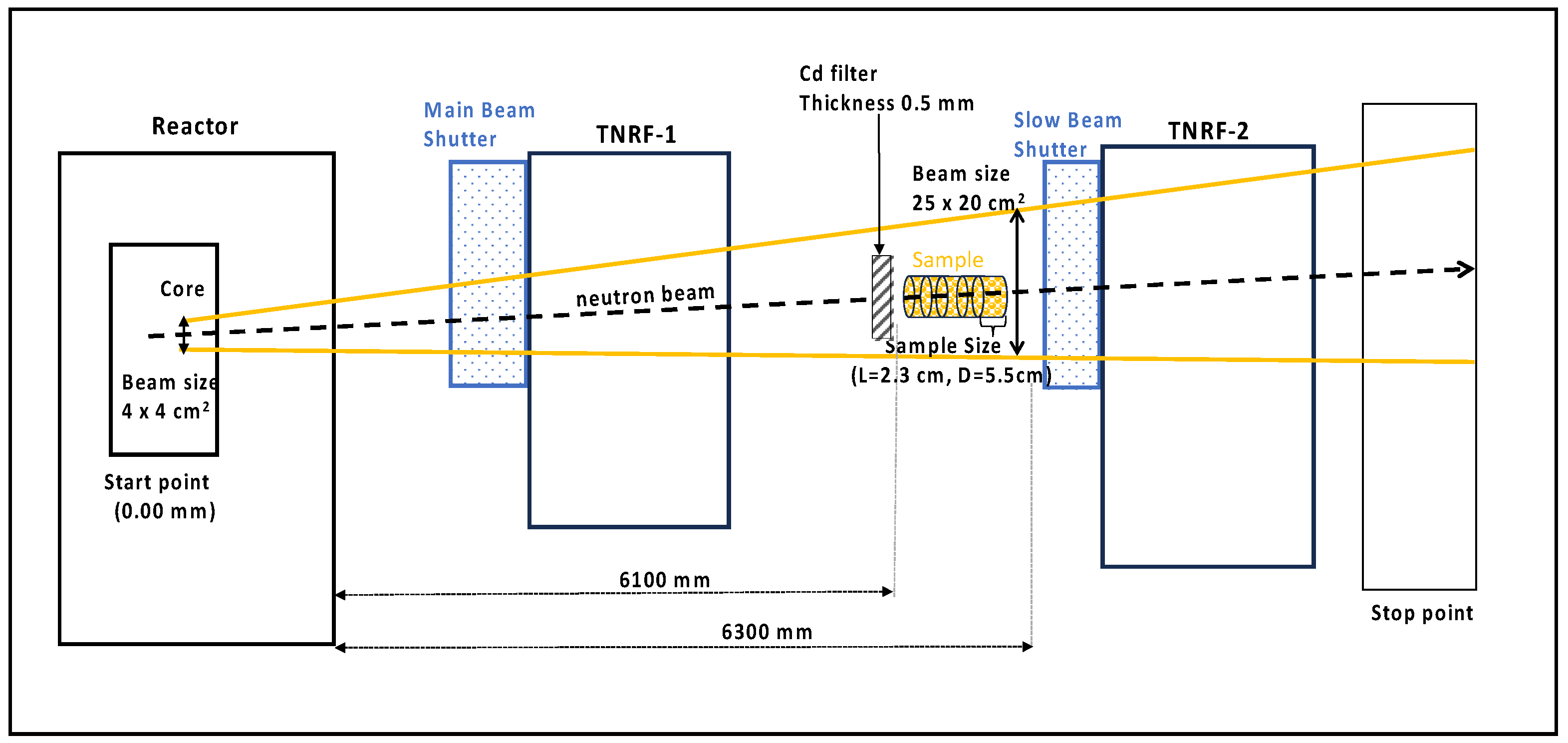
2.2. Irradiation Setup at JRR-3 (TNRF)
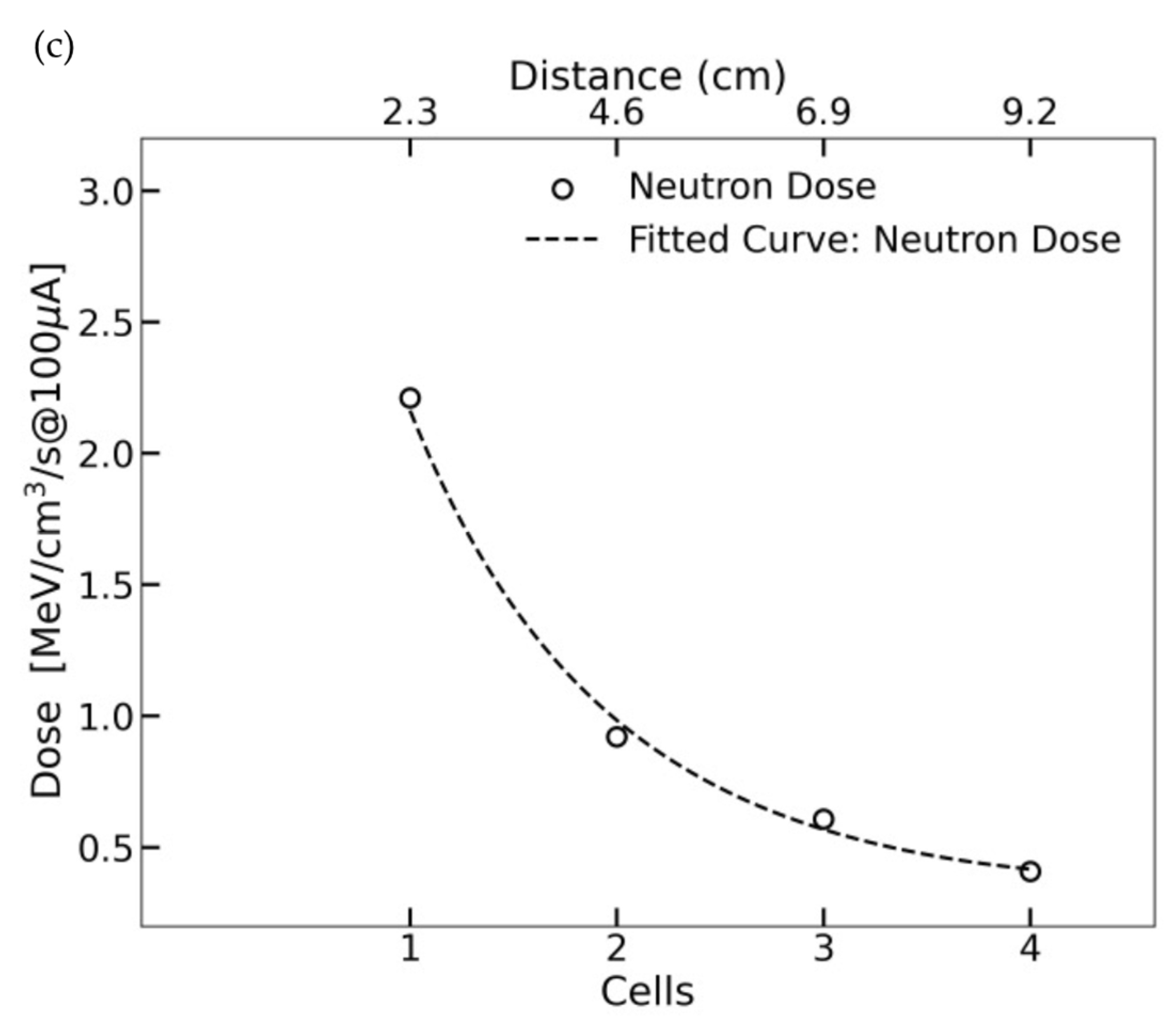
2.3. Simulation of a Compact Neutron Source (KUANS)
3. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamada, R.; Kashihara, T.; Ogino, H. Improvement of lipid production by the oleaginous yeast Rhodosporidium toruloides through UV mutagenesis. World J. Microbiol. Biotechnol. 2017, 33, 99. [Google Scholar] [CrossRef]
- Jo, Y.D.; Kim, J.B. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses of plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef]
- Yuan, N.; Liang, S.; Zhou, L.; Yuan, X.; Li, C.; Chen, X.; Zhao, H. Comparison of Mutations Induced by Different Doses of Fast-Neutron Irradiation in the M1 Generation of Sorghum (Sorghum bicolor). Genes 2024, 15, 976. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Zhang, Y.; Huang, M.; Guo, T.; Yang, G. Genome-Wide Profile of Mutations Induced by Carbon Ion Beam Irradiation of Dehulled Rice Seeds. Int. J. Mol. Sci. 2024, 25, 5195. [Google Scholar] [CrossRef]
- Jankowicz-Cieslak, J.; Hofinger, B.J.; Jarc, L.; Junttila, S.; Galik, B.; Gyenesei, A.; Ingelbrecht, I.L.; Till, B.J. Spectrum and Density of Gamma and X-ray Induced Mutations in a Non-Model Rice Cultivar. Plants 2022, 11, 3232. [Google Scholar] [CrossRef]
- Ogawa, H.; Shilviada, K.; Tomizawa, J.-I. Studies on Radiation-sensitive Mutants of E. coli I. Mutants Defective in the Repair Synthesis. Mol. Gen. Genet. 1968, 101, 227–244. [Google Scholar] [CrossRef]
- Goodhead, D.T. Mechanisms for the Biological Effectiveness of High-LET Radiations. J. Radiat. Res. 1999, 40, S1–S13. [Google Scholar] [CrossRef]
- Kazama, Y.; Saito, H.; Yamamoto, Y.Y.; Hayashi, Y.; Ichida, H.; Ryuto, H.; Fukunishi, N.; Abe, T. LET-dependent effects of heavy-ion beam irradiation in Arabidopsis thaliana. Plant Biotechnol. 2008, 25, 113–117. [Google Scholar] [CrossRef]
- Ma, L.; Kong, F.; Sun, K.; Wang, T.; Guo, T. From Classical Radiation to Modern Radiation: Past, Present, and Future of Radiation Mutation Breeding. Front. Public Health 2021, 9, 768071. [Google Scholar] [CrossRef]
- Ishii, K.; Kazama, Y.; Morita, R.; Hirano, T.; Ikeda, T.; Usuda, S.; Hayashi, Y.; Ohbu, S.; Motoyama, R.; Nagamura, Y.; et al. Linear energy transfer-dependent change in rice gene expression profile after heavy-ion beam irradiation. PLoS ONE 2016, 11, e0160061. [Google Scholar] [CrossRef]
- Hwang, W.J.; Kim, M.Y.; Kang, Y.J.; Shim, S.; Stacey, M.G.; Stacey, G.; Lee, S.-H. Genome-wide analysis of mutations in a dwarf soybean mutant induced by fast neutron bombardment. Euphytica 2015, 203, 399–408. [Google Scholar] [CrossRef]
- Nikitaki, Z.; Velalopoulou, A.; Zanni, V.; Tremi, I.; Havaki, S.; Kokkoris, M.; Gorgoulis, V.G.; Koumenis, C.; Georgakilas, A.G. Key biological mechanisms involved in high-LET radiation therapies with a focus on DNA damage and repair. Expert. Rev. Mol. Med. 2022, 24, e15. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, S.; Rana, N.; Bansal, R.; Vishwakarma, G.; Mehetre, S.T.; Das, B.K.; Kumar, M.; Yadav, S.K.; Sonah, H.; Sharma, T.R.; et al. Expanding avenue of fast neutron mediated mutagenesis for crop improvement. Plant 2019, 8, 164. [Google Scholar] [CrossRef]
- Feng, Z.; Du, Y.; Chen, J.; Chen, X.; Ren, W.; Wang, L.; Zhou, L. Comparison and Characterization of Phenotypic and Genomic Mutations Induced by a Carbon-Ion Beam and Gamma-ray Irradiation in Soybean (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2023, 24, 8825. [Google Scholar] [CrossRef]
- Marcu, D.; Damian, G.; Cosma, C.; Cristea, V. Gamma radiation effects on seed germination, growth and pigment content, and ESR study of induced free radicals in maize (Zea mays). J. Biol. Phys. 2013, 39, 625–634. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Century, K.; Straight, S.; Ronald, P.; Dong, X.; Lassner, M.; Zhang, Y. A fast neutron deletion mutagenesis-based reverse genetics system for plants. Plant J. 2001, 27, 235–242. [Google Scholar] [CrossRef]
- Maekawa, F.; Oikawa, K.; Harada, M.; Kai, T.; Meigo, S.; Kasugai, Y.; Ooi, M.; Sakai, K.; Teshigawara, M.; Hasegawa, S.; et al. NOBORU: J-PARC BL10 for facility diagnostics and its possible extension to innovative instruments. Nucl. Instrum. Methods Phys. Res. Sect. A 2009, 600, 335–337. [Google Scholar] [CrossRef]
- Matsubayashi, M.; Kobayashi, H.; Hibiki, T.; Mishima, K. Design and characteristics of the JRR-3M thermal neutron radiography facility and its imaging systems. Nucl. Technol. 2000, 132, 309–324. [Google Scholar] [CrossRef]
- IAEA (International Atomic Energy Agency). Compact Accelerator Based Neutron Sources; IAEA Nuclear Energy Series No. NP-T-5.8. International Atomic Energy Agency: Vienna, Austria, 2021. Available online: https://www.iaea.org/publications/14709/compact-accelerator-based-neutron-sources (accessed on 8 March 2025).
- Tasaki, S.; Nagae, T.; Hirose, M.; Yamashita, Y.; Hironaka, K.; Abe, Y.; Yamagata, Y.; Otake, Y.; Hirota, K. Properties and possible applications of Kyoto University accelerator-based neutron source (KUANS). Phys. Procedia 2014, 60, 181–185. [Google Scholar] [CrossRef]
- Sato, T.; Iwamoto, Y.; Hashimoto, S.; Ogawa, T.; Furuta, T.; Abe, S.-I.; Kai, T.; Matsuya, Y.; Matsuda, N.; Hirata, Y.; et al. Recent improvements of the particle and heavy ion transport code system—PHITS version 3.33. J. Nucl. Sci. Technol. 2024, 61, 127–135. [Google Scholar] [CrossRef]
- Harada, M.; Teshigawara, M.; Ohi, M.; Klinkby, E.; Zanini, L.; Batkov, K.; Oikawa, K.; Toh, Y.; Kimura, A.; Ikeda, Y. Experimental validation of the brightness distribution on the surfaces of coupled and decoupled moderators composed of 99.8% parahydrogen at the J-PARC pulsed spallation neutron source. Nucl. Instrum. Methods Phys. Res. Sect. A 2018, 903, 38–45. [Google Scholar] [CrossRef]
- Quantum Flowers & Foods Co., Ltd. Neutron Ray Irradiation Target Apparatus, Mutation Induction Method, and Irradiation Target Manufacturing Method. U.S. Patent No. 11980148 B2, 14 May 2024. Available online: https://qff.jp/en/news/notice-of-obtaining-u-s-patent (accessed on 8 March 2025).
- Harada, M.; Teshigawara, M.; Ooi, M.; Oikawa, K.; Takada, H.; Ikeda, Y. Experimental characterization of high-energy component in extracted pulsed neutrons at the J-PARC spallation neutron source. Nucl. Instrum. Methods Phys. Res. Sect. A 2021, 1000, 165252. [Google Scholar] [CrossRef]
- Ziegler, J.F.; Manoyan, J.M. The Stopping of Ions in Compounds. Nucl. Instrum. Methods Phys. Res. Sect. B 1988, 35, 215–228. [Google Scholar] [CrossRef]
- Xu, D.; Li, Y.; Yao, Z.; Yin, Y.; Feng, H.; Wei, Z. Study on the Relationship between Seed Absorbed Dose and Seed Composition of 252Cf Neutron Source Irradiated Bean Seed. Sci. Rep. 2019, 9, 9635. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.H.; Macklin, R.L. Total Neutron Yields from Light Elements under Proton and Alpha Bombardment. Phys. Rev. 1959, 114, 571. [Google Scholar] [CrossRef]
- Kadam, S.T.; Vishwakarma, G.; Kashyap, Y.; Shukla, M.; Roy, T.; Sahu, P.K.; Sharma, D.; Shitre, A.S.; Kumar, V.; Das, B.K. Thermal neutron as a potential mutagen for induced plant mutation breeding: Radiosensitivity response on wheat and rice. Genet. Resour. Crop Evol. 2022, 70, 789–798. [Google Scholar] [CrossRef]
- Ma, L.; Kazama, Y.; Hirano, T.; Morita, R.; Tanaka, S.; Abe, T.; Hatakeyama, S. LET dependence on killing effect and mutagenicity in the model filamentous fungus Neurospora crassa. Int. J. Radiat. Biol. 2018, 94, 1125–1133. [Google Scholar] [CrossRef]
- Kazama, Y.; Hirano, T.; Saito, H.; Liu, Y.; Ohbu, S.; Hayashi, Y.; Abe, T. Characterization of highly efficient heavy-ion mutagenesis in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 161. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, M.; Gao, Y.; Cao, G.; Yang, Y.; Lu, D.; Li, W. A genome-wide view of mutations in respiration-deficient mutants of Saccharomyces cerevisiae selected following carbon ion beam irradiation. Appl. Microbiol. Biotechnol. 2019, 103, 1851–1864. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Shimizu, A.; Hase, Y.; Tanaka, A.; Shikazono, N.; Degi, K.; Morishita, T. Effects of ion beam irradiation on mutation induction and nuclear DNA content in Chrysanthemum. Breed. Sci. 2010, 60, 398–404. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Facility | Beam Power | Neutron Flux [n/cm2/s] | Total Energy Deposited [MeV/cm3/s] ([Gy/h]) | Energy Deposited by Recoil Protons [MeV/cm3/s] | Energy Deposited by Electrons [MeV/cm3/s] |
|---|---|---|---|---|---|
| J-PARC BL10 (w/o B4C) | 1 MW | 3.2 × 108 | 6.7 × 106 (3.86) | 5.7 × 106 | 1.0 × 105 |
| J-PARC BL10 (w/B4C) | 1 MW | 1.0 × 108 | 4.4 × 106 (2.5) | 3.7 × 106 | 8.8 × 103 |
| JRR-3 TNRF (w/o Cd) | 20 MW (thermal power) | 1.4 × 108 | 6.6 × 105 (0.38) | 4.3 × 105 | 1.9 × 105 |
| JRR-3 TNRF (w/Cd) | 20 MW (thermal power) | 2.1 × 107 | 1.4 × 105 (0.08) | 1.0 × 105 | 2.3 × 104 |
| KUANS | 3.5 MeV × 100 µA | 4.6 × 108 | 3.0 × 107 (17) | 2.1 × 107 | 5.3 × 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sweet, M.; Mishima, K.; Harada, M.; Kurita, K.; Iikura, H.; Tasaki, S.; Kikuchi, N. Simulation Study on Dose and LET of Neutron Irradiation for Biological Experiments Using Spallation, Reactor, and Compact Neutron Sources. Quantum Beam Sci. 2025, 9, 11. https://doi.org/10.3390/qubs9020011
Sweet M, Mishima K, Harada M, Kurita K, Iikura H, Tasaki S, Kikuchi N. Simulation Study on Dose and LET of Neutron Irradiation for Biological Experiments Using Spallation, Reactor, and Compact Neutron Sources. Quantum Beam Science. 2025; 9(2):11. https://doi.org/10.3390/qubs9020011
Chicago/Turabian StyleSweet, May, Kenji Mishima, Masahide Harada, Keisuke Kurita, Hiroshi Iikura, Seiji Tasaki, and Norio Kikuchi. 2025. "Simulation Study on Dose and LET of Neutron Irradiation for Biological Experiments Using Spallation, Reactor, and Compact Neutron Sources" Quantum Beam Science 9, no. 2: 11. https://doi.org/10.3390/qubs9020011
APA StyleSweet, M., Mishima, K., Harada, M., Kurita, K., Iikura, H., Tasaki, S., & Kikuchi, N. (2025). Simulation Study on Dose and LET of Neutron Irradiation for Biological Experiments Using Spallation, Reactor, and Compact Neutron Sources. Quantum Beam Science, 9(2), 11. https://doi.org/10.3390/qubs9020011