Color Can Shorten Breakthrough Times in Continuous Flash Suppression through Increased Salience and Task Relevance


Abstract
1. Introduction
1.1. Background
1.2. The Present Study
2. Materials and Methods
2.1. Participants
2.2. Apparatus
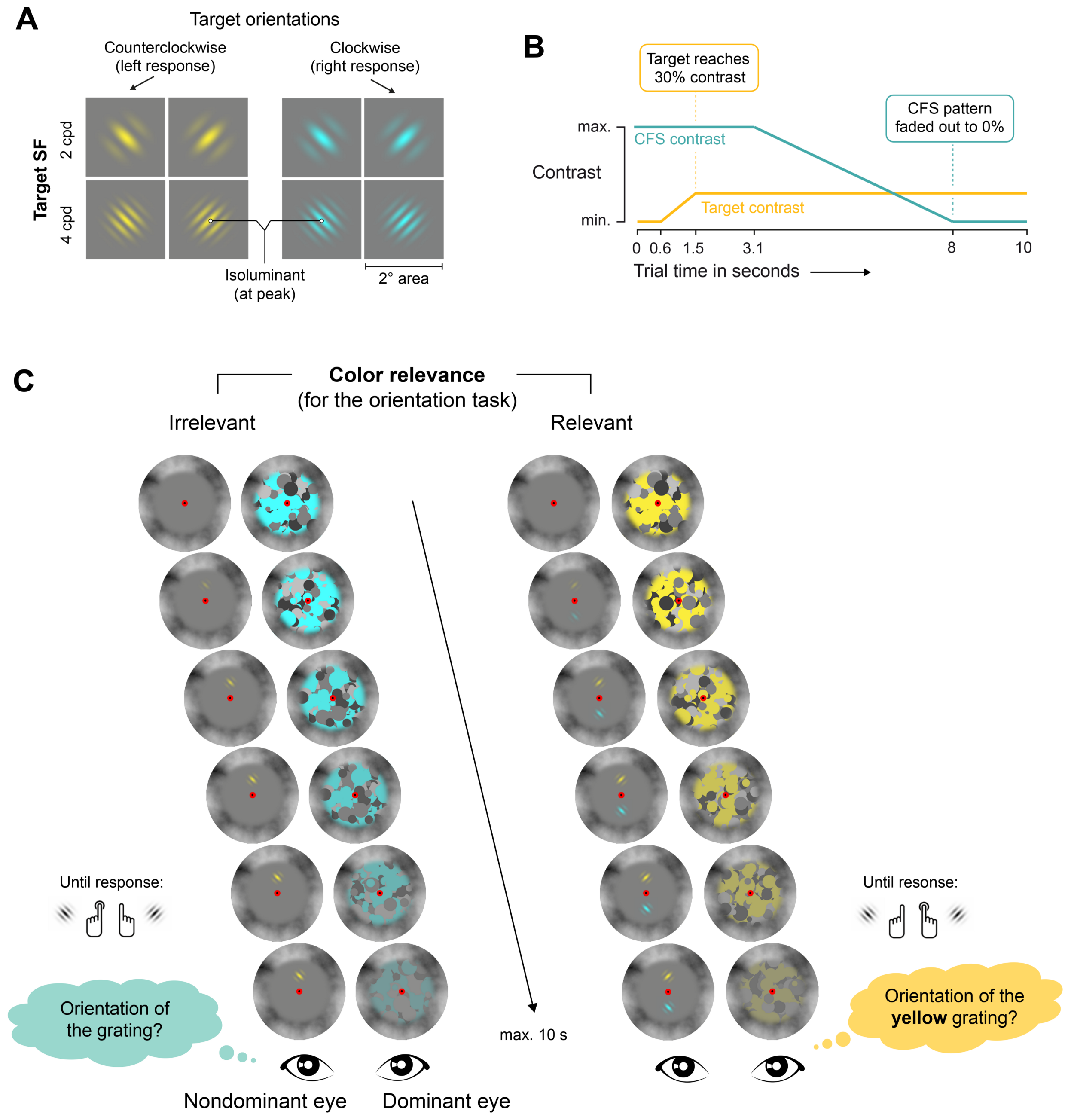
2.3. Stimuli
2.4. Procedure and Design
2.4.1. Eye Dominance Assessment
2.4.2. Trial Procedure
2.4.3. Task
2.4.4. Experimental Design
3. Results
3.1. Description of the Dataset
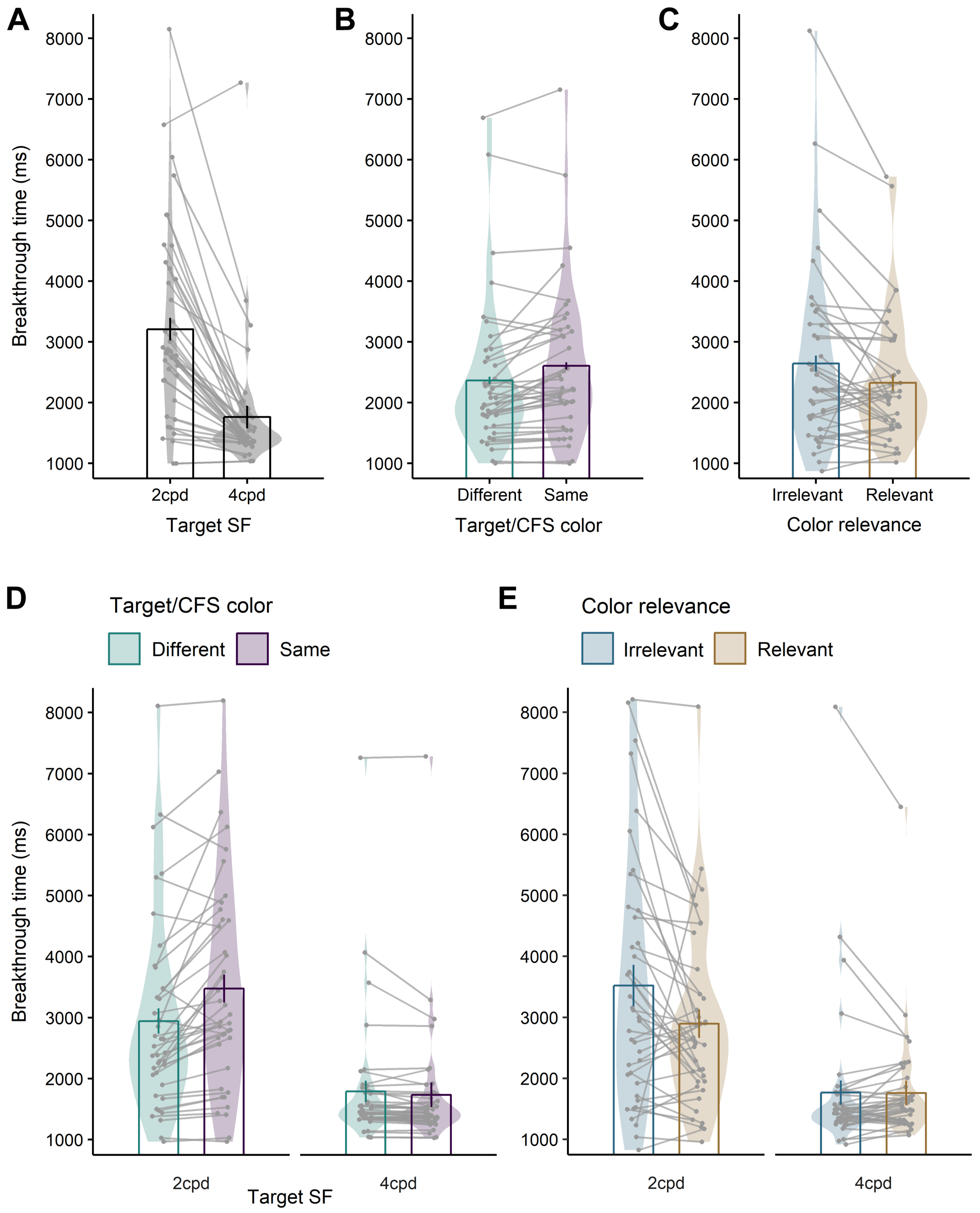
3.2. Breakthrough Time
3.2.1. Main Analysis of Experimental Effects
3.2.2. Role of the Distractor for the Task Effect
3.3. Error Rates
4. Discussion
4.1. The Bottom-Up Effect of Color Salience on Breakthrough Times
4.2. The Top-Down Effect of Task Relevance on Breakthrough Times
4.3. Limitations
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BR | Binocular Rivalry |
| CFS | Continuous Flash Suppression |
| cpd | Cycles per degree of visual angle |
| CRT | Cathode Ray Tube |
| HSF | High Spatial Frequency |
| LSF | Low Spatial Frequency |
| SF | Spatial Frequency |
| VWM | Visual Working Memory |
References
- Foster, D.H.; Nascimento, S.M. Relational colour constancy from invariant cone-excitation ratios. Proc. R. Soc. Ser. B Biol. Sci. 1994, 257, 115–121. [Google Scholar]
- Nascimento, S.M.; Ferreira, F.P.; Foster, D.H. Statistics of spatial cone-excitation ratios in natural scenes. J. Opt. Soc. Am. A 2002, 19, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Sumner, P.; Mollon, J.D. Catarrhine photopigments are optimized for detecting targets against a foliage background. J. Exp. Biol. 2000, 203, 1963–1986. [Google Scholar] [PubMed]
- Pflüger, L.S.; Valuch, C.; Gutleb, D.R.; Ansorge, U.; Wallner, B. Colour and contrast of female faces: Attraction of attention and its dependence on male hormone status in Macaca fuscata. Anim. Behav. 2014, 94, 61–71. [Google Scholar] [CrossRef]
- Einhäuser, W.; Spain, M.; Perona, P. Objects predict fixations better than early saliency. J. Vis. 2008, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Theeuwes, J. Perceptual selectivity for color and form. Percept. Psychophys. 1992, 51, 599–606. [Google Scholar] [CrossRef]
- Itti, L.; Koch, C. A saliency-based search mechanism for overt and covert shifts of visual attention. Vis. Res. 2000, 40, 1489–1506. [Google Scholar] [CrossRef]
- Duncan, J.; Humphreys, G.W. Visual search and stimulus similarity. Psychol. Rev. 1989, 96, 433. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.M.; Horowitz, T.S. What attributes guide the deployment of visual attention and how do they doit? Nat. Rev. Neurosci. 2004, 5, 495–501. [Google Scholar] [CrossRef]
- Gayet, S.; Douw, I.; Van der Burg, V.; Van der Stigchel, S.; Paffen, C.L. Hide and seek: Directing top-down attention is not sufficient for accelerating conscious access. Cortex 2020, 122, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Mack, A.; Rock, I. Inattentional Blindness; MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Dehaene, S.; Changeux, J.P.; Naccache, L.; Sackur, J.; Sergent, C. Conscious, preconscious, and subliminal processing: A testable taxonomy. Trends Cogn. Sci. 2006, 10, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Parise, C. Prior-entry: A review. Conscious. Cogn. 2010, 19, 364–379. [Google Scholar] [CrossRef]
- Koch, C.; Tsuchiya, T. Attention and consciousness: Related yet different. Trends Cogn. Sci. 2015, 16, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Lamy, D.; Alon, L.; Carmel, T.; Shalev, N. The role of conscious perception in attentional capture and object-file updating. Psychol. Sci. 2015, 26, 48–57. [Google Scholar] [CrossRef]
- Kim, C.Y.; Blake, R. Psychophysical magic: Rendering the visible “invisible”. Trends Cogn. Sci. 2005, 9, 381–388. [Google Scholar] [CrossRef]
- Dehaene, S.; Changeux, J.-P. Experimental and theoretical approaches to conscious processing. Neuron 2011, 70, 200–227. [Google Scholar] [CrossRef]
- Breitmeyer, B.G. Psychophysical “blinding” methods reveal a functional hierarchy of unconscious visual processing. Conscious. Cogn. 2015, 35, 234–250. [Google Scholar] [CrossRef]
- Tsuchiya, N.; Koch, C. Continuous flash suppression reduces negative afterimages. Nat. Neurosci. 2005, 8, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.; Hebart, M.N.; Sterzer, P. Breaking Continuous Flash Suppression: A New Measure of Unconscious Processing during Interocular Suppression? Front. Hum. Neurosci. 2011, 5, 167. [Google Scholar] [CrossRef] [PubMed]
- Valuch, C.; Mattler, U. Action priming is linked to visual perception in continuous flash suppression. J. Vis. 2019, 19, 13. [Google Scholar] [CrossRef]
- Levelt, W.J.M. On Binocular Rivalry; Van Gorcum: Assen, The Netherlands, 1965. [Google Scholar]
- Brascamp, J.W.; Baker, D.H. Psychophysics of binocular rivalry. In The Consitution of Visual Consciousness; Miller, S.M., Ed.; John Benjamins: Amsterdam, The Netherlands, 2013; pp. 109–139. [Google Scholar]
- Tsuchiya, N.; Koch, C.; Gilroy, L.A.; Blake, R. Depth of interocular suppression associated with continuous flash suppression, flash suppression, and binocular rivalry. J. Vis. 2006, 6, 6. [Google Scholar] [CrossRef]
- Valuch, C.; Kulke, L. Predictive context biases binocular rivalry in children and adults with no positive relation to two measures of social cognition. Sci. Rep. 2020, 10, 2059. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Blake, R. Deconstructing continuous flash suppression. J. Vis. 2012, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Lunghi, C.; Alais, D. The temporal frequency tuning of continuous flash suppression reveals peak suppression at very low frequencies. Sci. Rep. 2016, 6, 35723. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Blake, R.; McDonald, J.E. A new interocular suppression technique for measuring sensory eye dominance. Investig. Ophthalmol. Vis. Sci. 2010, 51, 588–593. [Google Scholar] [CrossRef]
- Ding, Y.; Naber, M.; Gayet, S.; Van der Stigchel, S.; Paffen, C.L.E. Assessing the generalizability of eye dominance across binocular rivalry, onset rivalry, and continuous flash suppression. J. Vis. 2018, 18, 6. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Loftus, G.R.; Masson, M.E. Using confidence intervals in within-subjects designs. Psychon. Bull. Rev. 1994, 1, 476–490. [Google Scholar] [CrossRef] [PubMed]
- Gayet, S.; Stein, T. Between-subject variability in the breaking continuous flash suppression paradigm: Potential causes, consequences, and solutions. Front. Psychol. 2017, 8, 437. [Google Scholar] [CrossRef][Green Version]
- Stuit, S.M.; Cass, J.; Paffen, C.L.; Alais, D. Orientation-tuned suppression in binocular rivalry reveals general and specific components of rivalry suppression. J. Vis. 2009, 9, 17. [Google Scholar] [CrossRef][Green Version]
- O’Shea, R.P.; Sims, A.J.; Govan, D.G. The effect of spatial frequency and field size on the spread of exclusive visibility in binocular rivalry. Vis. Res. 1997, 37, 175–183. [Google Scholar] [CrossRef]
- Moors, P.; Wagemans, J.; de-Wit, L. Moving stimuli are less effectively masked using traditional continuous flash suppression (CFS) compared to a moving mondrian mask (MMM): A test case for feature-selective suppression and retinotopic adaptation. PLoS ONE 2014, 9, e98298. [Google Scholar] [CrossRef] [PubMed]
- Drewes, J.; Zhu, W.; Melcher, D. The edge of awareness: Mask spatial density, but not color, determines optimal temporal frequency for continuous flash suppression. J. Vis. 2018, 18, 12. [Google Scholar] [CrossRef]
- Ding, Y.; Paffen, C.L.; Naber, M.; Van der Stigchel, S. Visual working memory and saliency independently influence the priority for access to visual awareness. J. Vis. 2019, 19, 9. [Google Scholar] [CrossRef]
- Ludwig, K.; Hesselmann, G. Weighing the evidence for a dorsal processing bias under continuous flash suppression. Conscious. Cogn. 2015, 35, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Gayet, S.; Stein, T.; Peelen, M.V. The danger of interpreting detection differences between image categories: A brief comment on “Mind the snake: Fear detection relies on low spatial frequencies”. (Gomes, Soares, Silva, & Silva, 2018). Emotion 2019, 19, 928–932. [Google Scholar]
- Niedenthal, P.M.; Wood, A. Does emotion influence visual perception? Depends on how you look at it. Cogn. Emot. 2019, 33, 77–84. [Google Scholar] [CrossRef]
- Gayet, S.; Paffen, C.L.; Belopolsky, A.V.; Theeuwes, J.; Van der Stigchel, S. Visual input signaling threat gains preferential access to awareness in a breaking continuous flash suppression paradigm. Cognition 2016, 149, 77–83. [Google Scholar] [CrossRef]
- Gayet, S.; Paffen, C.L.; Van der Stigchel, S. Information matching the content of visual working memory is prioritized for conscious access. Psychol. Sci. 2013, 24, 2472–2480. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Lin, B.; Zhao, Y.; Soto, D. Working memory biasing of visual perception without awareness. Atten. Percept. Psychophys. 2014, 76, 2051–2062. [Google Scholar] [CrossRef]
- Olivers, C.N.L.; Meijer, F.; Theeuwes, J. Feature-based memory-driven attentional capture: Visual working memory content affects visual attention. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 1243–1265. [Google Scholar] [CrossRef] [PubMed]
- Gayet, S.; Guggenmos, M.; Christophel, T.B.; Haynes, J.D.; Paffen, C.L.; Sterzer, P.; Van der Stigchel, S. No evidence for mnemonic modulation of interocularly suppressed visual input. NeuroImage 2020, 215, 116801. [Google Scholar] [CrossRef]
- Van Moorselaar, D.; Gayet, S.; Paffen, C.L.E.; Theeuwes, J.; Van der Stigchel, S.; Olivers, C.N.L. Competitive interactions in visual working memory drive access to awareness. Cortex 2018, 102, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Frătescu, M.; Van Moorselaar, D.; Mathôt, S. Can you have multiple attentional templates? Large-scale replications of Van Moorselaar, Theeuwes, and Olivers (2014) and Hollingworth and Beck (2016). Atten. Percept. Psychophys. 2019, 81, 2700–2709. [Google Scholar] [CrossRef]
- Huang, L.; Pashler, H. A Boolean map theory of visual attention. Psychol. Rev. 2007, 114, 599–631. [Google Scholar] [CrossRef]
- Olivers, C.N.; Peters, J.; Houtkamp, R.; Roelfsema, P.R. Different states in visual working memory: When it guides attention and when it does not. Trends Cogn. Sci. 2011, 15, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Büsel, C.; Pomper, U.; Ansorge, U. Capture of attention by target-similar cues during dual-color search reflects reactive control among top-down selected attentional control settings. Psychon. Bull. Rev. 2019, 26, 531–537. [Google Scholar] [CrossRef]
- Stein, T.; Peelen, M.V. Content-specific expectations enhance stimulus detectability by increasing perceptual sensitivity. J. Exp. Psychol. Gen. 2015, 144, 1089. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y.; van Gaal, S.; de Lange, F.P.; Lamme, V.A.; Seth, A.K. Expectations accelerate entry of visual stimuli into awareness. J. Vis. 2015, 15, 13. [Google Scholar] [CrossRef] [PubMed]
- Lanfranco, R.; Stein, T.; Rabagliati, H.; Carmel, D. Dissociating perceptual sensitivity from decisional factors in unconscious processing of face orientation and gaze direction. PsyArXiv 2020. [Google Scholar] [CrossRef]
- Green, D.M.; Swets, J.A. Signal Detection Theory and Psychophysics; Wiley: New York, NY, USA, 1966. [Google Scholar]
- Santhi, N.; Reeves, A. The roles of distractor noise and target certainty in search: A signal detection model. Vis. Res. 2004, 44, 1235–1256. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| LSF (2 cpd) | HSF (4 cpd) | |||
|---|---|---|---|---|
| Different | Same | Different | Same | |
| Breakthrough time (ms) | ||||
| Color relevant | 2675 (216.1) | 3117 (257.5) | 1778 (143.2) | 1738 (137.9) |
| Color irrelevant | 3209 (312.2) | 3832 (344.2) | 1801 (201.1) | 1731 (190.1) |
| Error Rate (%) | ||||
| Color relevant | 3.05 (0.62) | 4.35 (0.84) | 2.76 (0.41) | 3.78 (0.54) |
| Color irrelevant | 4.59 (1.04) | 4.32 (0.95) | 2.53 (0.57) | 2.21 (0.42) |
| Error Rate (arcsine transform) | ||||
| Color relevant | 0.13 (0.02) | 0.18 (0.02) | 0.14 (0.01) | 0.17 (0.01) |
| Color irrelevant | 0.17 (0.02) | 0.15 (0.02) | 0.13 (0.02) | 0.11 (0.02) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valuch, C. Color Can Shorten Breakthrough Times in Continuous Flash Suppression through Increased Salience and Task Relevance. Vision 2021, 5, 13. https://doi.org/10.3390/vision5010013
Valuch C. Color Can Shorten Breakthrough Times in Continuous Flash Suppression through Increased Salience and Task Relevance. Vision. 2021; 5(1):13. https://doi.org/10.3390/vision5010013
Chicago/Turabian StyleValuch, Christian. 2021. "Color Can Shorten Breakthrough Times in Continuous Flash Suppression through Increased Salience and Task Relevance" Vision 5, no. 1: 13. https://doi.org/10.3390/vision5010013
APA StyleValuch, C. (2021). Color Can Shorten Breakthrough Times in Continuous Flash Suppression through Increased Salience and Task Relevance. Vision, 5(1), 13. https://doi.org/10.3390/vision5010013