The Limitations of Reward Effects on Saccade Latencies: An Exploration of Task-Specificity and Strength



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Apparatus
2.3. Stimuli
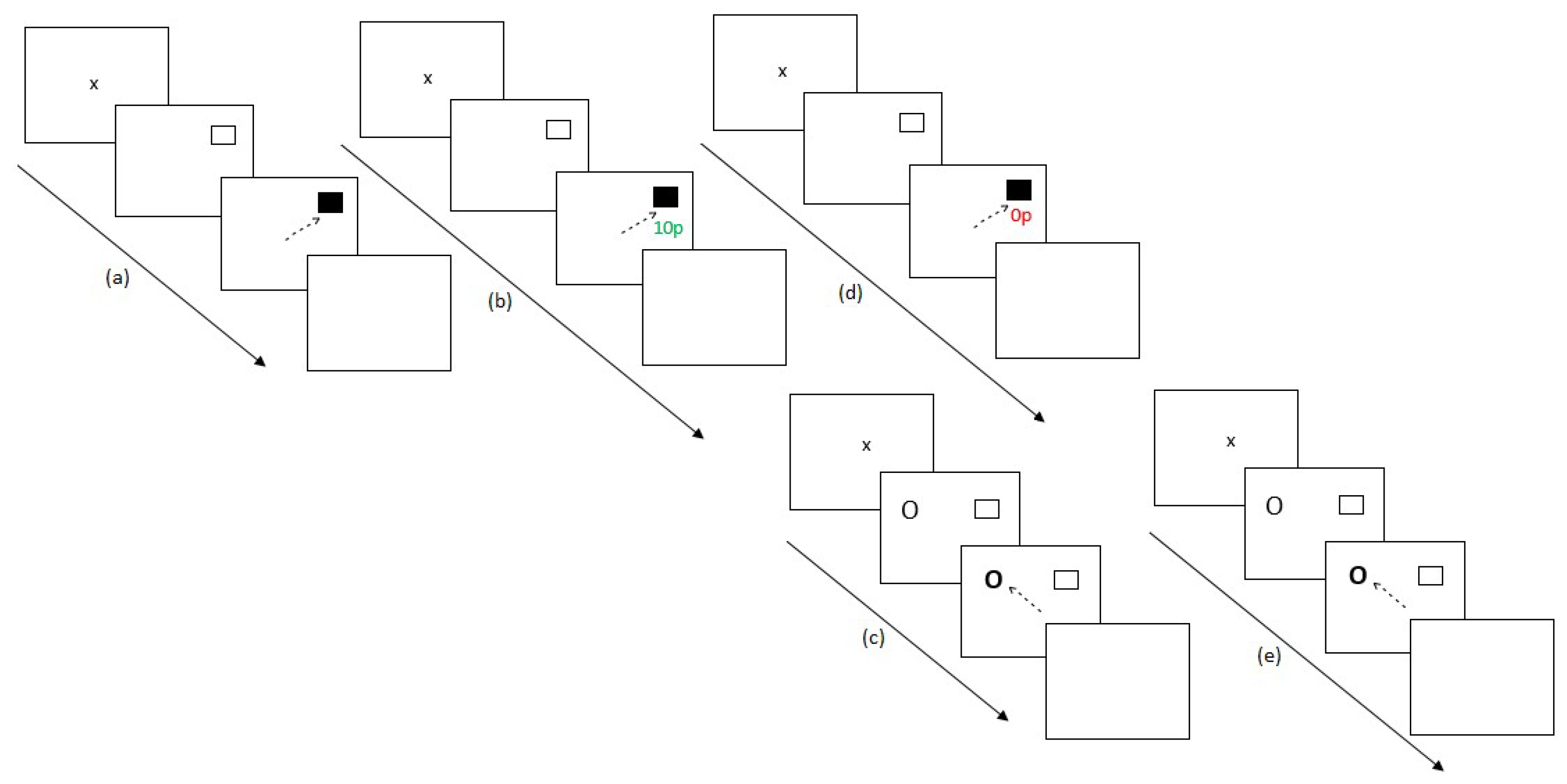
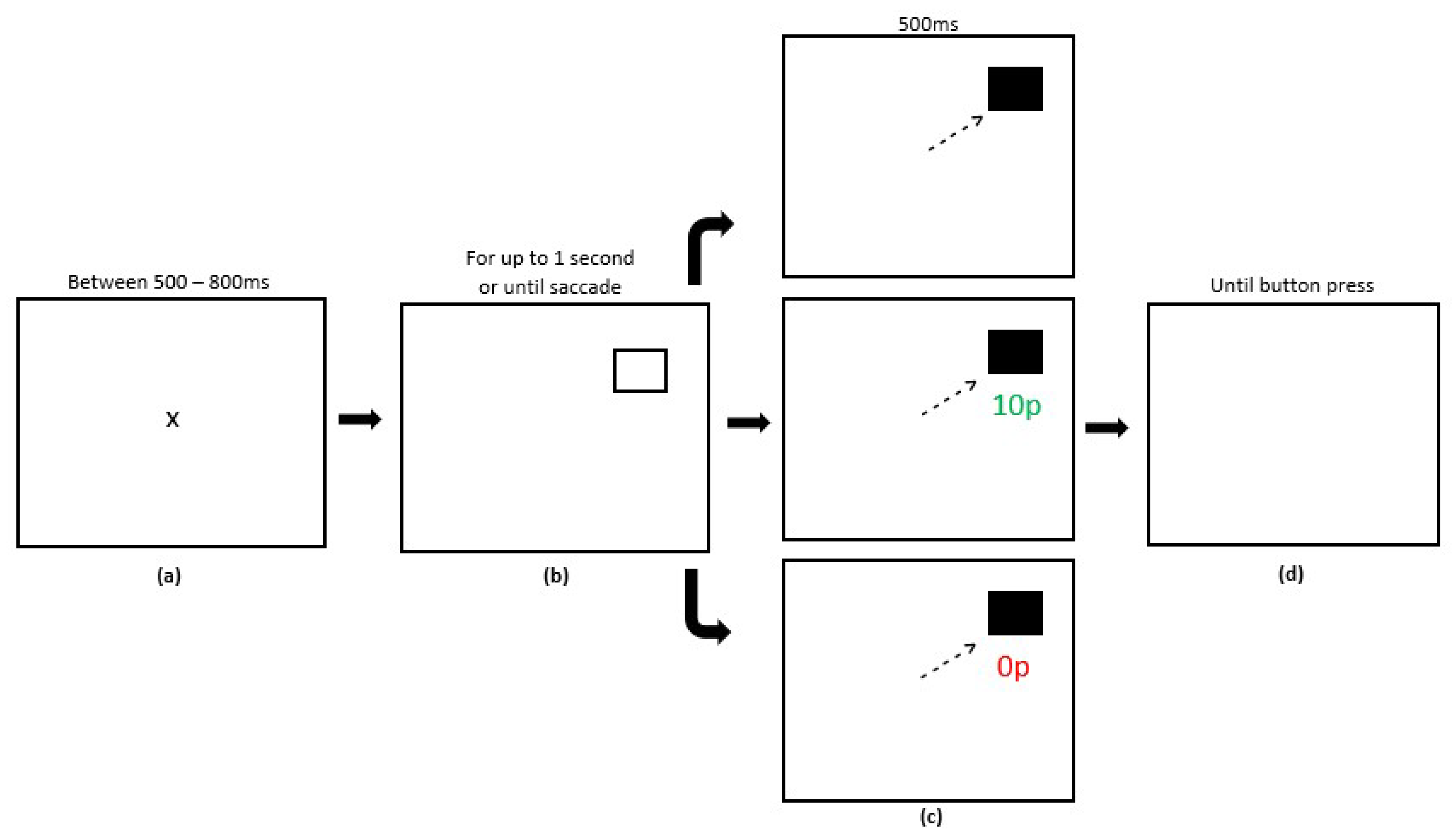
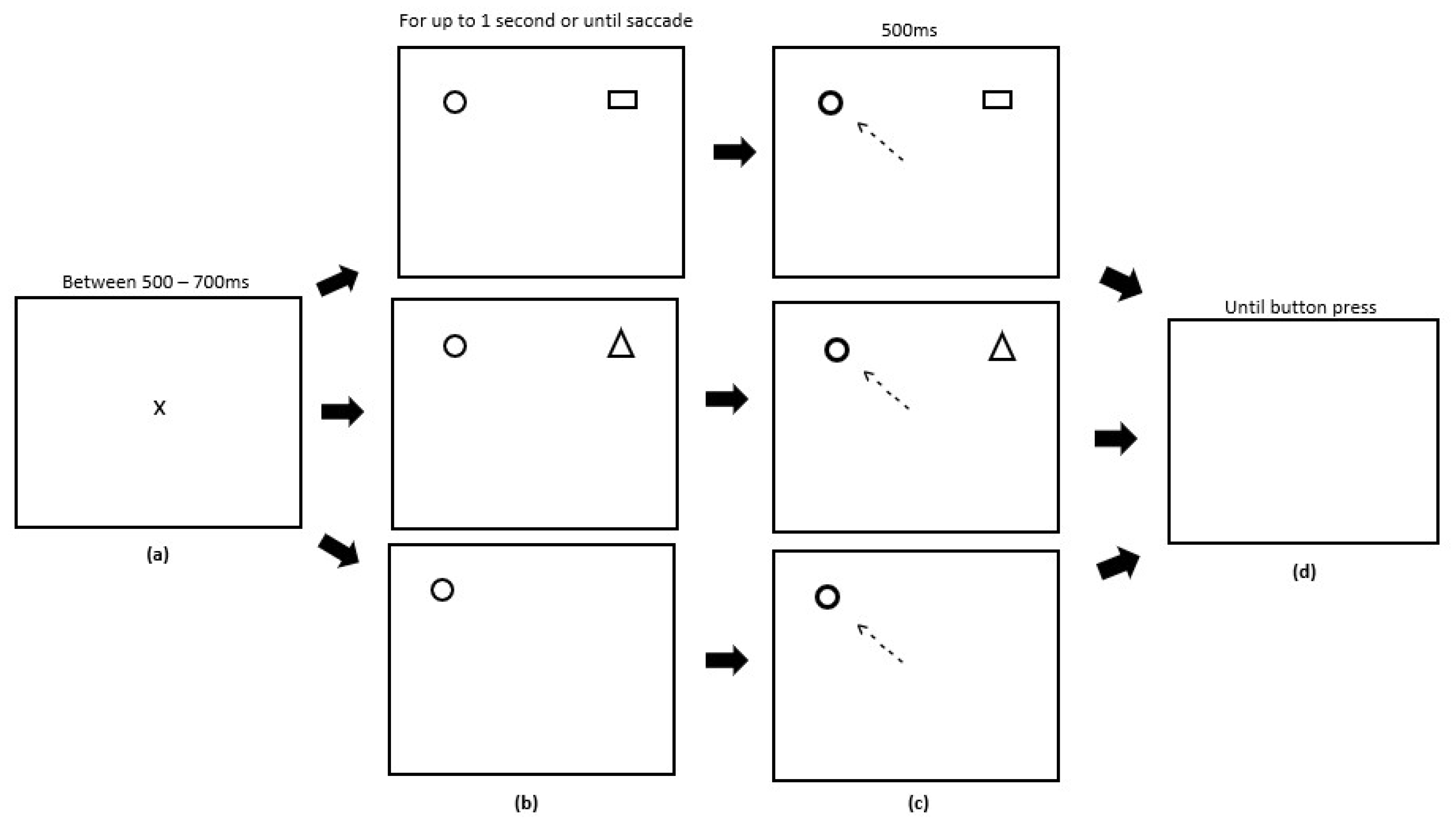
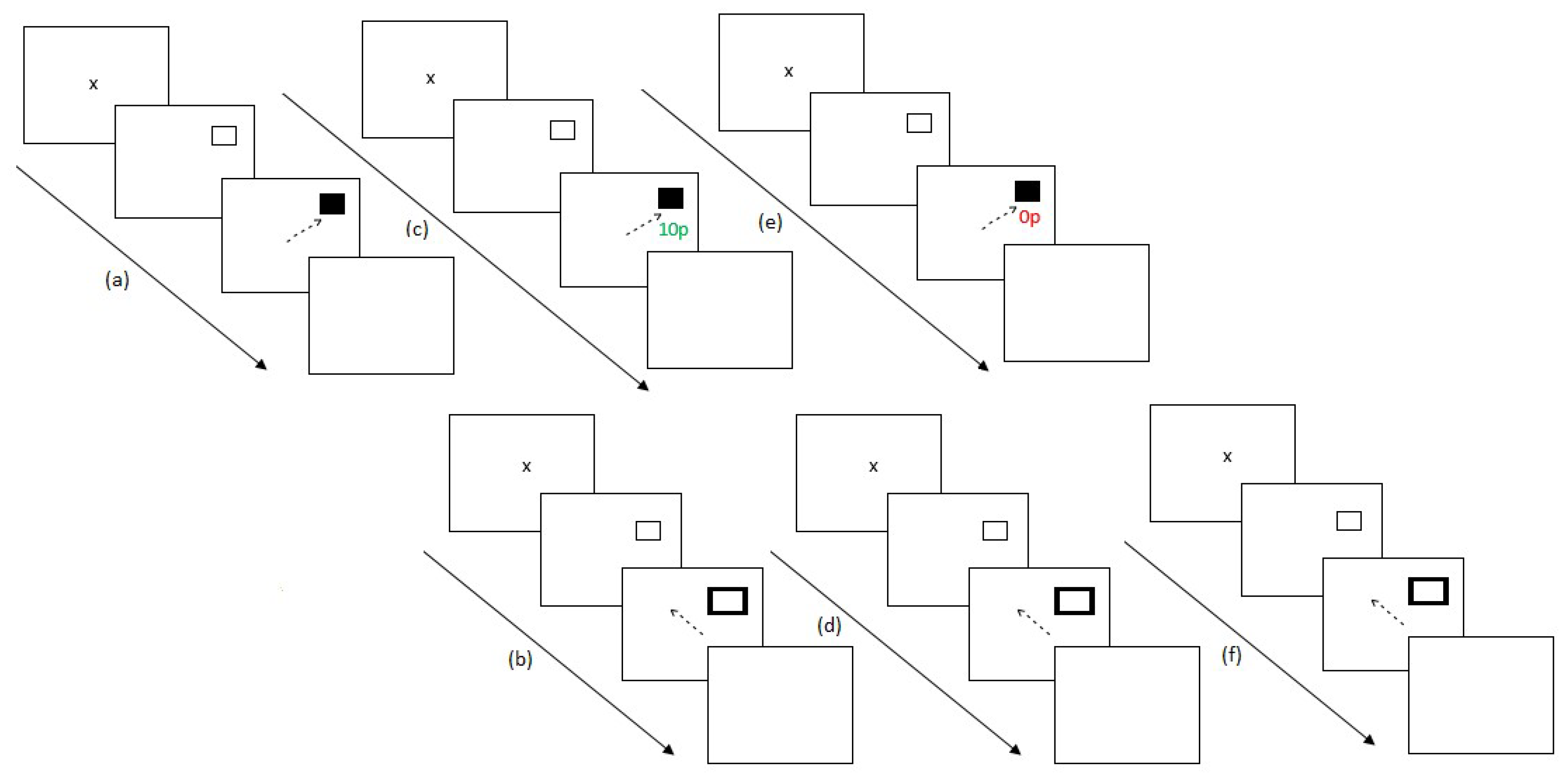
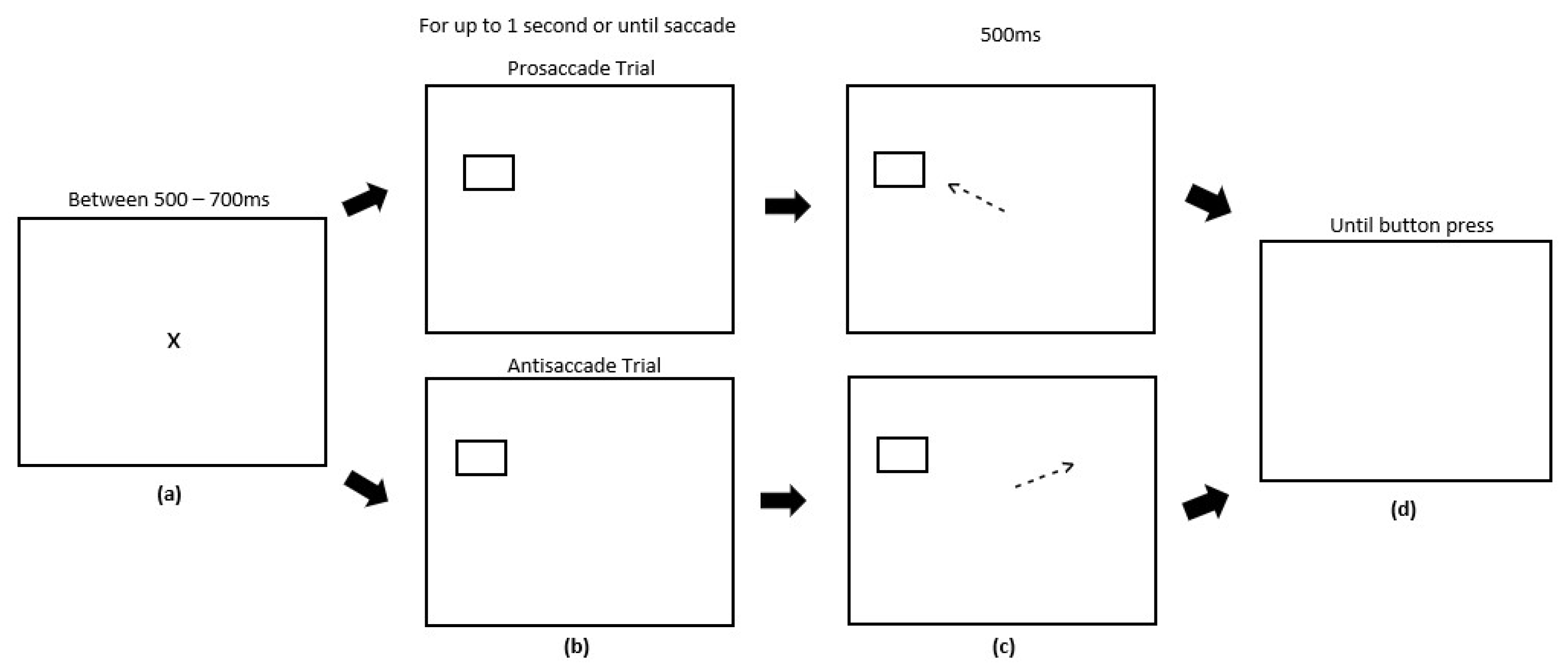
2.4. Procedure
2.4.1. Experiment 1
2.4.2. Experiment 2
2.5. Saccade Analysis
2.5.1. Experiment 1
2.5.2. Experiment 2
3. Results
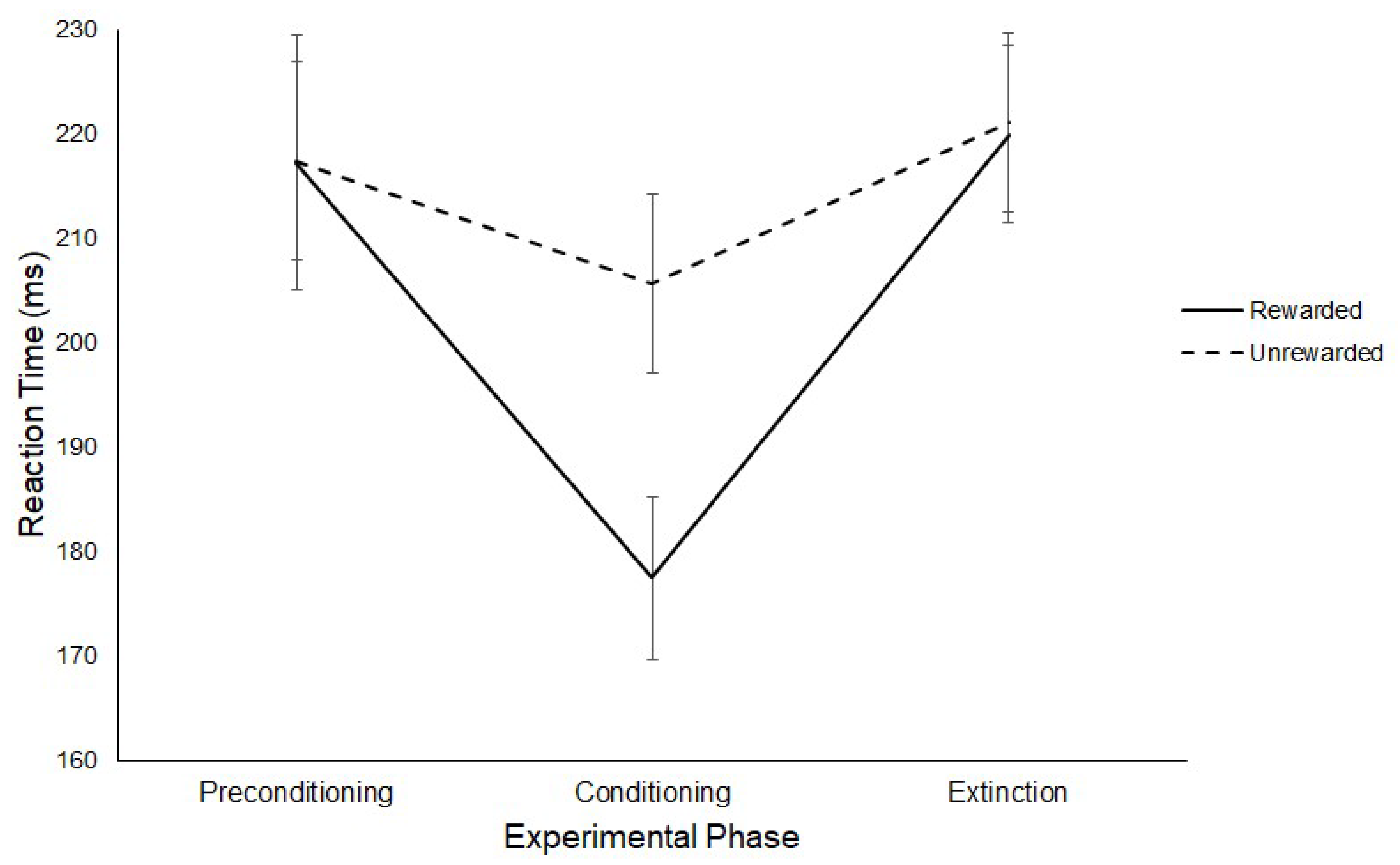
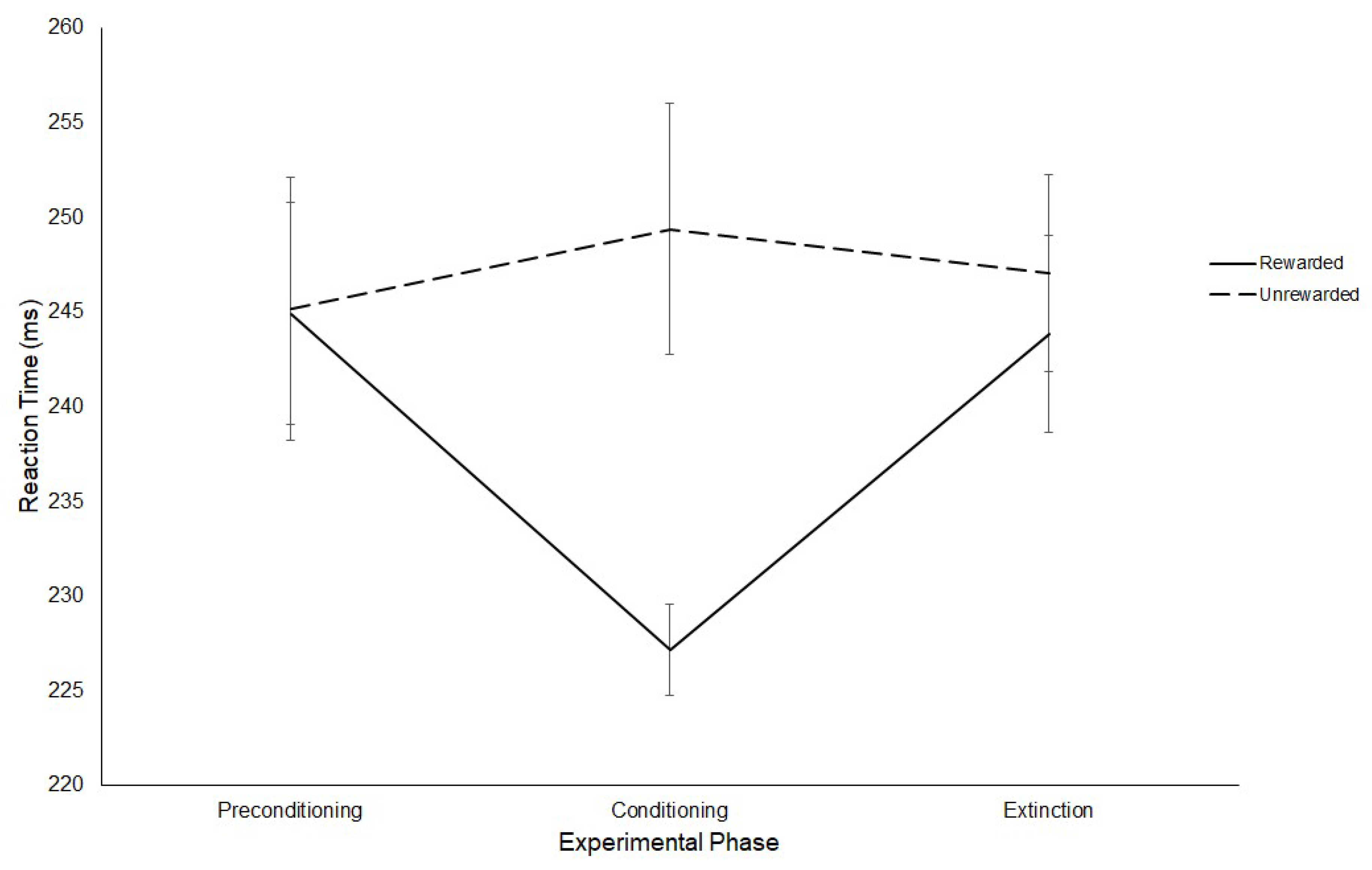
3.1. Latency
3.1.1. Experiment 1
Reward Paradigm
RD Task
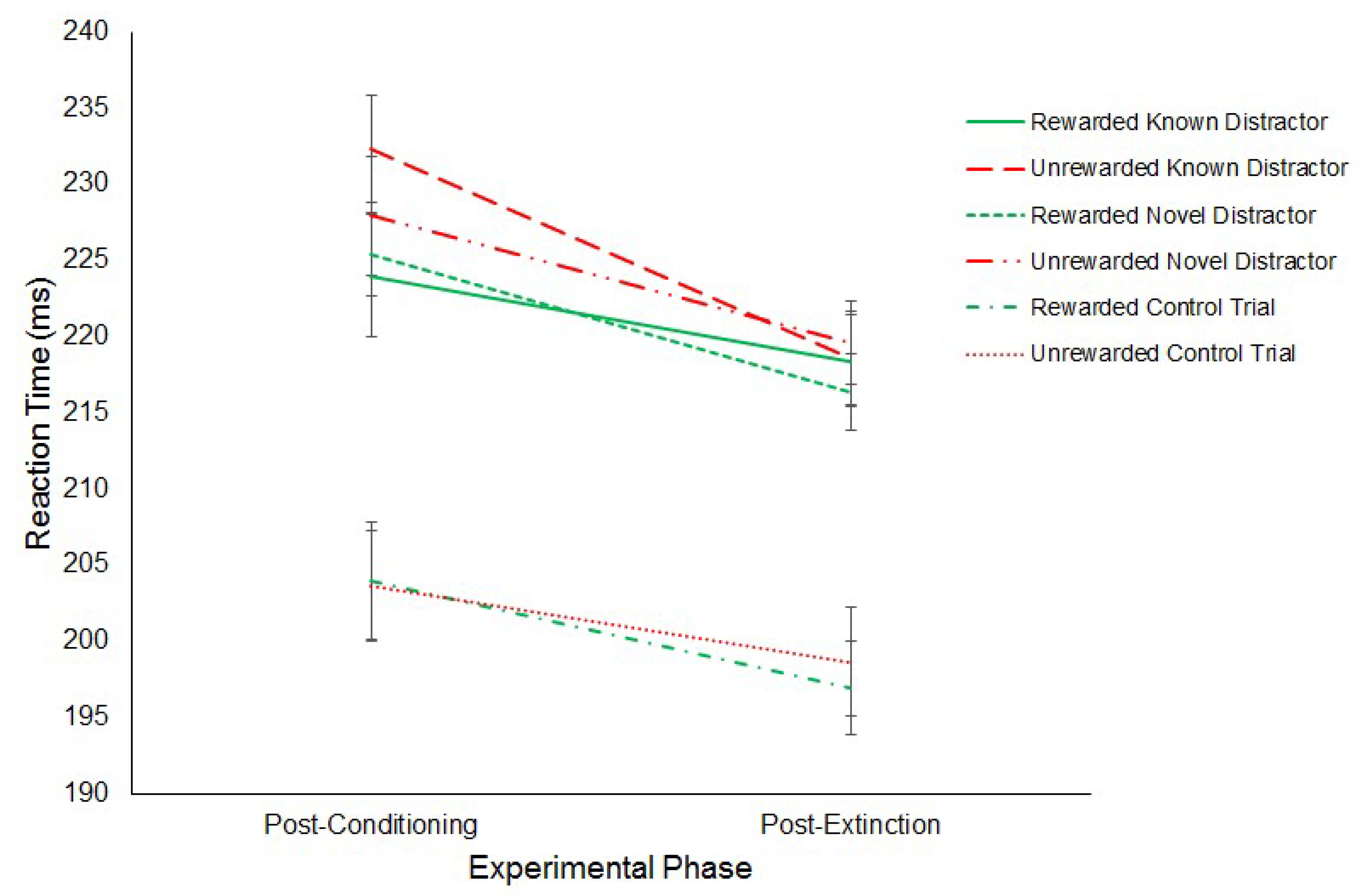
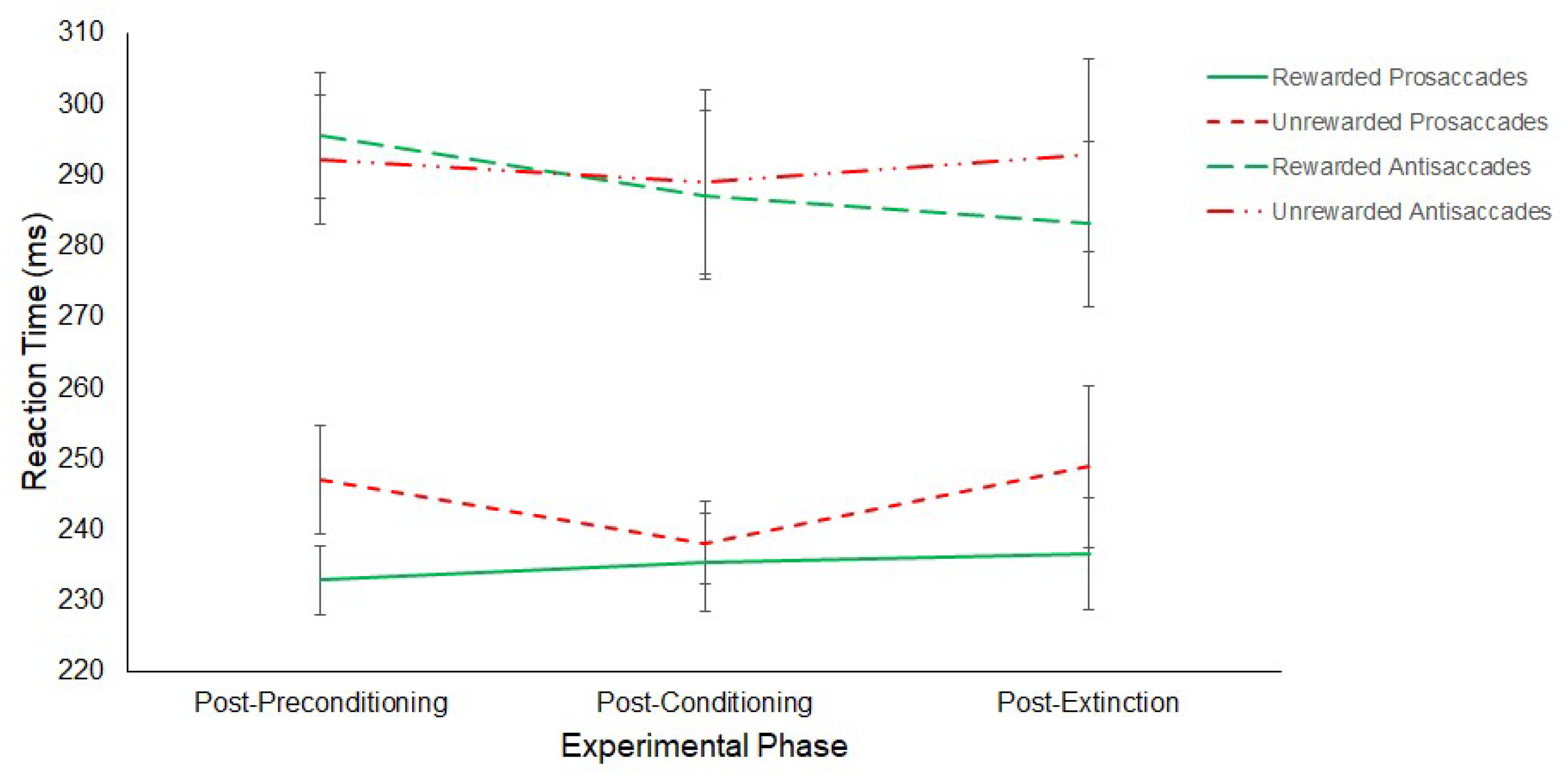
3.1.2. Experiment 2
Reward Paradigm
Antisaccade Task
3.2. Saccadic Error
3.2.1. Experiment 1
Reward Paradigm
RD Task
3.2.2. Experiment 2
Reward Paradigm
Antisaccade Task
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ludwig, C.J.; Gilchrist, I.D.; McSorley, E. The influence of spatial frequency and contrast on saccade latencies. Vis. Res. 2004, 44, 2597–2604. [Google Scholar] [CrossRef]
- Walker, R.; Deubel, H.; Schneider, W.X.; Findlay, J.M. Effect of remote distractors on saccade programming: Evidence for an extended fixation zone. J. Neurophysiol. 1997, 78, 1108–1119. [Google Scholar] [CrossRef]
- Walker, R.; Kentridge, R.W.; Findlay, J.M. Independent contributions of the orienting of attention, fixation offset and bilateral stimulation on human saccadic latencies. Exp. Brain Res. 1995, 103, 294–310. [Google Scholar] [CrossRef]
- Thorndike, E.L. The law of effect. Am. J. Psychol. 1927, 39, 212–222. [Google Scholar] [CrossRef]
- Ikeda, T.; Hikosaka, O. Reward-dependent gain and bias of visual responses in primate superior colliculus. Neuron 2003, 39, 693–700. [Google Scholar] [CrossRef]
- Lauwereyns, J.; Watanabe, K.; Coe, B.; Hikosaka, O. A neural correlate of response bias in monkey caudate nucleus. Nature 2002, 418, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Madelain, L.; Champrenaut, L.; Chauvin, A. Control of sensorimotor variability by consequences. J. Neurophysiol. 2007, 98, 2255–2265. [Google Scholar] [CrossRef] [PubMed]
- Milstein, D.M.; Dorris, M.C. The influence of expected value on saccadic preparation. J. Neurosci. 2007, 27, 4810–4818. [Google Scholar]
- Takikawa, Y.; Kawagoe, R.; Itoh, H.; Nakahara, H.; Hikosaka, O. Modulation of saccadic eye movements by predicted reward outcome. Exp. Brain Res. 2002, 142, 284–291. [Google Scholar] [CrossRef]
- Watanabe, K.; Lauwereyns, J.; Hikosaka, O. Neural correlates of rewarded and unrewarded eye movements in the primate caudate nucleus. J. Neurosci. 2003, 23, 10052–10057. [Google Scholar] [CrossRef]
- Brielmann, A.A.; Spering, M. Effects of reward on the accuracy and dynamics of smooth pursuit eye movements. J. Exp. Psychol. 2015, 41, 917. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, K.; Dalmaijer, E.; Manohar, S.; Husain, M. Voluntary modulation of saccadic peak velocity associated with individual differences in motivation. Cortex 2018. [Google Scholar] [CrossRef]
- Anderson, B.A.; Yantis, S. Value-driven attentional and oculomotor capture during goal-directed, unconstrained viewing. Atten. Percept. Psychophy. 2012, 74, 1644–1653. [Google Scholar] [CrossRef]
- Hickey, C.; Chelazzi, L.; Theeuwes, J. Reward changes salience in human vision via the anterior cingulate. J. Neurosci. 2010, 30, 11096–11103. [Google Scholar] [CrossRef]
- Hickey, C.; van Zoest, W. Reward creates oculomotor salience. Curr. Biol. 2012, 22, R219–R220. [Google Scholar] [CrossRef] [PubMed]
- Theeuwes, J.; Belopolsky, A.V. Reward grabs the eye: Oculomotor capture by rewarding stimuli. Vis. Res. 2012, 74, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Mccoy, B.; Theeuwes, J. Effects of reward on oculomotor control. J. Neurophysiol. 2016, 116, 2453–2466. [Google Scholar] [CrossRef]
- Duka, J.; Lupp, A. The effects of incentive on antisaccades: Is a dopaminergic mechanism involved? Behav. Pharmacol. 1997, 8, 373–382. [Google Scholar] [CrossRef]
- Jazbec, S.; Hardin, M.G.; Schroth, E.; Mcclure, E.; Pine, D.S.; Ernst, M. Age-related influence of contingencies on a saccade task. Exp. Brain Res. 2009, 174, 754–762. [Google Scholar] [CrossRef]
- Ross, M.; Lanyon, L.J.; Viswanathan, J.; Manoach, D.S.; Barton, J.J.S. Human prosaccades and antisaccades under risk: Effects of penalties and rewards on visual selection and the value of actions. Neuroscience 2011, 196, 168–177. [Google Scholar] [CrossRef]
- Anderson, B.A.; Laurent, P.A.; & Yantis, S. Learned Value Magnifies Salience-Based Attentional Capture. PLoS ONE 2011, 6, e27926. [Google Scholar] [CrossRef]
- Della Libera, C.; Chelazzi, L. Learning to attend and to ignore is a matter of gains and losses. Psychol. Sci. 2009, 20, 778–784. [Google Scholar] [CrossRef]
- Failing, M.; Theeuwes, J. Don’t let it distract you: How information about the availability of reward affects attentional selection. Atten. Percept. Psychophy. 2017, 1, 2275–2298. [Google Scholar] [CrossRef]
- Chelazzi, L.; Perlato, A.; Santandrea, E.; Della Libera, C. Rewards teach visual selective attention. Vis. Res. 2013, 85, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Knight, H.C.; Smith, D.T.; Knight, D.C.; Ellison, A. Altering attentional control settings causes persistent biases of visual attention. Q. J. Exp. Psychol. 2016, 69, 129–149. [Google Scholar] [CrossRef]
- Malhotra, P.A.; Soto, D.; Li, K.; Russell, C. Reward modulates spatial neglect. J. Neurol. Neurosurg. Psychiatry 2013, 84, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Kawagoe, R.; Takikawa, Y.; Hikosaka, O. Expectation of reward modulates cognitive signals in the basal ganglia. Nat. Neurosci. 1998, 1, 411–416. [Google Scholar] [CrossRef]
- Kojima, Y.; Soetedjo, R. Selective Reward Affects the Rate of Saccade Adaptation. Neuroscience 2017, 355, 113–125. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kim, H.F.; Hikosaka, O. Reward Value-Contingent Changes of Visual Responses in the Primate Caudate Tail Associated with a Visuomotor Skill. J. Neurosci. 2013, 33, 11227–11238. [Google Scholar] [CrossRef]
- Bendiksby, M.S.; Platt, M.L. Neural correlates of reward and attention in macaque area LIP. Neuropsychologia 2006, 44, 2411–2420. [Google Scholar] [CrossRef] [PubMed]
- Dunne, S.; Ellison, A.; Smith, D.T. Rewards modulate saccade latency but not exogenous spatial attention. Front Psychol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Heuer, A.; Schubö, A.; Schütz, A.C. The necessity to choose causes the effects of reward on saccade preparation. Sci Rep. 2017, 7, 16966. [Google Scholar] [CrossRef]
- Casteau, S.; Smith, D.T. Covert attention beyond the range of eye-movements: Evidence for a dissociation between exogenous and endogenous orienting. Cortex 2018. [Google Scholar] [CrossRef]
- Smith, D.T.; Schenk, T. The Premotor theory of attention: Time to move on? Neuropsychologia 2012, 50, 1104–1114. [Google Scholar] [CrossRef]
- Manohar, S.G.; Chong, T.T.J.; Apps, M.A.; Batla, A.; Stamelou, M.; Jarman, P.R.; Bhatia, K.P.; Husain, M. Reward pays the cost of noise reduction in motor and cognitive control. Curr. Biol. 2015, 25, 1707–1716. [Google Scholar] [CrossRef]
- Blaukopf, C.L.; DiGirolamo, G.J. Reward, context, and human behaviour. Sci. World J. 2007, 7, 626–640. [Google Scholar] [CrossRef]
- Theeuwes, J.A.N. Exogenous and endogenous control of attention: The effect of visual onsets and offsets. Percept. Psychophy. 1991, 49, 83–90. [Google Scholar] [CrossRef]
- Theeuwes, J.A.N. Perceptual selectivity for color and form. Percept. Psychophy. 1992, 51, 599–606. [Google Scholar] [CrossRef]
- Anderson, B.A. A value-driven mechanism of attentional selection stimulus value. J. Vis. 2013, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Failing, M.F.; Theeuwes, J. Exogenous visual orienting by reward. J. Vis. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Failing, M.F.; Theeuwes, J. Nonspatial attentional capture by previously rewarded scene semantics. Vis. Cogn. 2015, 23, 82–104. [Google Scholar] [CrossRef]
- Schütz, A.C.; Trommershäuser, J.; Gegenfurtner, K.R. Dynamic integration of information about salience and value for saccadic eye movements. Proc. Natl. Acad. Sci. USA 2012, 109, 7547–7552. [Google Scholar] [CrossRef] [PubMed]
- Lucas, N.; Schwartz, S.; Leroy, R.; Pavin, S.; Diserens, K.; Vuilleumier, P. Gambling against neglect: Unconscious spatial biases induced by reward reinforcement in healthy people and brain-damaged patients. Cortex 2013, 49, 2616–2627. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunne, S.; Ellison, A.; Smith, D.T. The Limitations of Reward Effects on Saccade Latencies: An Exploration of Task-Specificity and Strength. Vision 2019, 3, 20. https://doi.org/10.3390/vision3020020
Dunne S, Ellison A, Smith DT. The Limitations of Reward Effects on Saccade Latencies: An Exploration of Task-Specificity and Strength. Vision. 2019; 3(2):20. https://doi.org/10.3390/vision3020020
Chicago/Turabian StyleDunne, Stephen, Amanda Ellison, and Daniel T. Smith. 2019. "The Limitations of Reward Effects on Saccade Latencies: An Exploration of Task-Specificity and Strength" Vision 3, no. 2: 20. https://doi.org/10.3390/vision3020020
APA StyleDunne, S., Ellison, A., & Smith, D. T. (2019). The Limitations of Reward Effects on Saccade Latencies: An Exploration of Task-Specificity and Strength. Vision, 3(2), 20. https://doi.org/10.3390/vision3020020