
Transcriptional Signatures of Aerobic Exercise-Induced Muscle Adaptations in Humans



 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
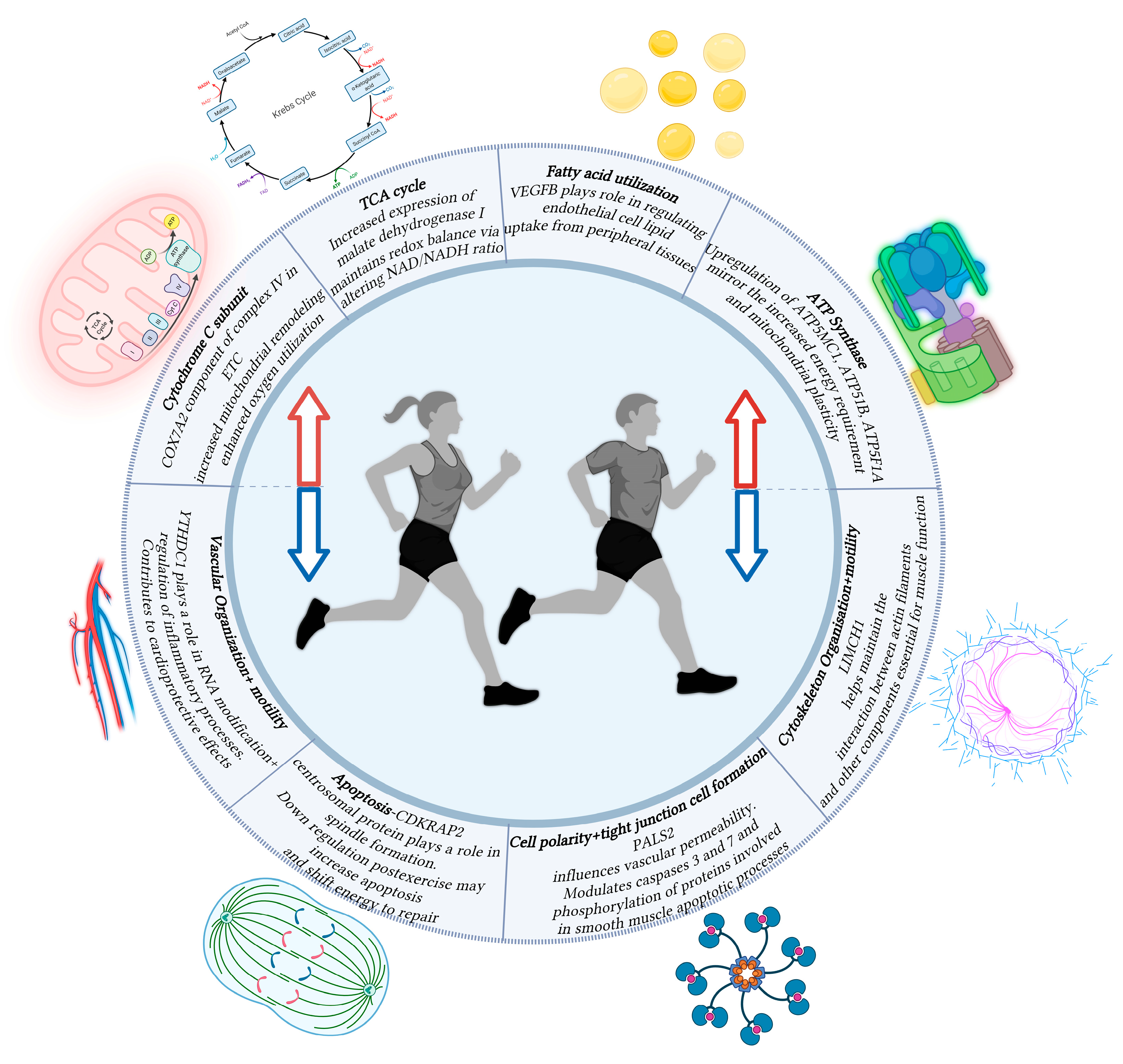
3. Results and Discussion
3.1. Elevated Gene Expression Involved in Cellular Respiration Following Aerobic Exercise
3.2. Aerobic Exercise Alter Genes Involved in Vascular and Cellular Organization
3.3. Regulation of Apoptosis Signaling Pathways Post-Exercise
3.4. Increased Fatty Acid Utilization Post-Exercise Through VEGFB Regulation
3.5. Exercise Modulates Novel Genes (LIMCH1, CMYA5, FOXJ3) Involved in Muscle Physiology
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Myers, J. Exercise and cardiovascular health. Circulation 2003, 107, e2–e5. [Google Scholar] [CrossRef] [PubMed]
- Fleg, J.L. Aerobic exercise in the elderly: A key to successful aging. Discov. Med. 2012, 13, 223–228. [Google Scholar] [PubMed]
- Egan, B.; Sharples, A.P. Molecular responses to acute exercise and their relevance for adaptations in skeletal muscle to exercise training. Physiol. Rev. 2023, 103, 2057–2170. [Google Scholar] [CrossRef] [PubMed]
- Hyldahl, R.D.; Xin, L.; Hubal, M.J.; Moeckel-Cole, S.; Chipkin, S.; Clarkson, P.M. Activation of nuclear factor-κPB following muscle eccentric contractions in humans is localized primarily to skeletal muscle-residing pericytes. FASEB J. 2011, 25, 2956–2966. [Google Scholar] [CrossRef] [PubMed]
- Kurz, E.; Hirsch, C.A.; Dalton, T.; Shadaloey, S.A.; Khodadadi-Jamayran, A.; Miller, G.; Pareek, S.; Rajaei, H.; Mohindroo, C.; Baydogan, S.; et al. Exercise-induced engagement of the IL-15/IL-15Rα axis promotes anti-tumor immunity in pancreatic cancer. Cancer Cell 2022, 40, 720–737.e5. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Saavedra, M.; De Repentigny, Y.; Yang, D.; O’meara, R.W.; Yan, K.; Hashem, L.E.; Racacho, L.; Ioshikhes, I.; Bulman, D.E.; Parks, R.J.; et al. Voluntary running triggers VGF-mediated oligodendrogenesis to prolong the lifespan of Snf2h-null ataxic mice. Cell Rep. 2016, 17, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.C.; Ellefsen, S.; Baar, K. Adaptations to endurance and strength training. Cold Spring Harb. Perspect. Med. 2018, 8, a029769. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Hahn, K.R.; Kang, M.S.; Choi, J.H.; Moon, S.M.; Yoon, Y.S.; Hwang, I.K.; Kim, D.W. Tat-malate dehydrogenase fusion protein protects neurons from oxidative and ischemic damage by reduction of reactive oxygen species and modulation of glutathione redox system. Sci. Rep. 2023, 13, 5653. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ford, H.C.; Carroll, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Persistence of the mitochondrial permeability transition in the absence of subunit c of human ATP synthase. Proc. Natl. Acad. Sci. USA 2017, 114, 3409–3414. [Google Scholar] [CrossRef] [PubMed]
- Choong, C.-J.; Okuno, T.; Ikenaka, K.; Baba, K.; Hayakawa, H.; Koike, M.; Yokota, M.; Doi, J.; Kakuda, K.; Takeuchi, T.; et al. Alternative mitochondrial quality control mediated by extracellular release. Autophagy 2021, 17, 2962–2974. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zou, M.-H. AMPK, mitochondrial function, and cardiovascular disease. Int. J. Mol. Sci. 2020, 21, 4987. [Google Scholar] [CrossRef] [PubMed]
- Yubero, D.; Montero, R.; Martín, M.Á.; Montoya, J.; Ribes, A.; Grazina, M.; Trevisson, E.; Aguilera, J.C.R.; Hargreaves, I.P.; Salviati, L.; et al. Secondary coenzyme Q10 deficiencies in oxidative phosphorylation (OXPHOS) and non-OXPHOS disorders. Mitochondrion 2016, 30, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, L.; Cui, X.; Zheng, S.; Li, R. Roles and mechanisms of the m6A reader YTHDC1 in biological processes and diseases. Cell Death Discov. 2022, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chen, S.; Wang, X.; Yang, X.; Chen, L.; Huang, T.; Zheng, Y.; Zheng, X.; Wu, X.; Sun, Y.; et al. Exercise mitigates endothelial pyroptosis and atherosclerosis by downregulating NEAT1 through N6-methyladenosine modifications. Arterioscler. Thromb. Vasc. Biol. 2023, 43, 910–926. [Google Scholar] [CrossRef] [PubMed]
- Yeh, J.I.; Chinte, U.; Du, S. Structure of glycerol-3-phosphate dehydrogenase, an essential monotopic membrane enzyme involved in respiration and metabolism. Proc. Natl. Acad. Sci. USA 2008, 105, 3280–3285. [Google Scholar] [CrossRef] [PubMed]
- Langston, P.K.; Nambu, A.; Jung, J.; Shibata, M.; Aksoylar, H.I.; Lei, J.; Xu, P.; Doan, M.T.; Jiang, H.; MacArthur, M.R.; et al. Glycerol phosphate shuttle enzyme GPD2 regulates macrophage inflammatory responses. Nat. Immunol. 2019, 20, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wideranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, H.; Zhang, H. Abnormal expression of mitochondrial ribosomal proteins and their encoding genes with cell apoptosis and diseases. Int. J. Mol. Sci. 2020, 21, 8879. [Google Scholar] [CrossRef] [PubMed]
- Millet, G.P.; Debevec, T.; Brocherie, F.; Malatesta, D.; Girard, O. Therapeutic use of exercising in hypoxia: Promises and limitations. Front. Physiol. 2016, 7, 224. [Google Scholar] [CrossRef] [PubMed]
- Cheong, A.; Lingutla, R.; Mager, J. Expression analysis of mammalian mitochondrial ribosomal protein genes. Gene Expr. Patterns 2020, 38, 119147. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Rhee, K. Importance of the CEP215-pericentrin interaction for centrosome maturation during mitosis. PLoS ONE 2014, 9, e87016. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Yigit, G.; E Brown, K.; Kayserili, H.; Pohl, E.; Caliebe, A.; Zahnleiter, D.; Rosser, E.; Bögershausen, N.; Uyguner, Z.O.; Altunoglu, U.; et al. Mutations in CDK 5 RAP 2 cause Seckel syndrome. Mol. Genet. Genom. Med. 2015, 3, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Ching, Y.-P.; Qi, Z.; Wang, J.H. Cloning of three novel neuronal Cdk5 activator binding proteins. Gene 2000, 242, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Rosales, J.L.; Gao, X.; Lee, K.-Y. Centromeric chromatin integrity is compromised by loss of Cdk5rap2, a transcriptional activator of CENP-A. Biomed. Pharmacother. 2021, 138, 111463. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed]
- McVey, D.G.; Andreadi, C.; Gong, P.; Stanczyk, P.J.; Solomon, C.U.; Turner, L.; Yan, L.; Chen, R.; Cao, J.; Nelson, C.P.; et al. Genetic influence on vascular smooth muscle cell apoptosis. Cell Death Dis. 2024, 15, 402. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, B.; Pajusola, K.; Kaipainen, A.; Von Euler, G.; Joukov, V.; Saksela, O.; Orpana, A.; Pettersson, R.F.; Alitalo, K.; Eriksson, U. Vascular endothelial growth factor B, a novel growth factor for endothelial cells. Proc. Natl. Acad. Sci. USA 1996, 93, 2576–2581. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, J.-S.; Tamarat, R.; Ebrahimian, T.G.; Le-Roux, A.; Clergue, M.; Emmanuel, F.; Duriez, M.; Schwartz, B.; Branellec, D.; Lévy, B.I. Vascular endothelial growth factor-B promotes in vivo angiogenesis. Circ. Res. 2003, 93, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, C.E.; Falkevall, A.; Wang, X.; Larsson, E.; Huusko, J.; Nilsson, I.; van Meeteren, L.A.; Samen, E.; Lu, L.; Vanwildemeersch, M.; et al. Vascular endothelial growth factor B controls endothelial fatty acid uptake. Nature 2010, 464, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.B.; Petersen, K.F.; Shulman, G.I. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol. Rev. 2007, 87, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Randle, P.; Garland, P.; Hales, C.; Newsholme, E. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 281, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.; Quan, L.; Lai, X.; Lang, L.; Li, F.; Yang, X.; Fu, Y.; Feng, S.; Yi, X.; Zhu, C.; et al. VEGFB promotes myoblasts proliferation and differentiation through VEGFR1-PI3K/Akt signaling pathway. Int. J. Mol. Sci. 2021, 22, 13352. [Google Scholar] [CrossRef] [PubMed]
- Landgren, H.; Carlsson, P. FoxJ3, a novel mammalian forkhead gene expressed in neuroectoderm, neural crest, and myotome. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 231, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.S.; Shi, X.; Voelker, K.A.; Grange, R.W.; Garcia, J.A.; Hammer, R.E.; Garry, D.J. Foxj3 transcriptionally activates Mef2c and regulates adult skeletal muscle fiber type identity. Dev. Biol. 2010, 337, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Kadrmas, J.L.; Beckerle, M.C. The LIM domain: From the cytoskeleton to the nucleus. Nat. Rev. Mol. Cell Biol. 2004, 5, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Fiuza-Luces, C.; Santos-Lozano, A.; Llavero, F.; Campo, R.; Nogales-Gadea, G.; Díez-Bermejo, J.; Baladrón, C.; González-Murillo, Á.; Arenas, J.; Martín, M.A.; et al. Muscle molecular adaptations to endurance exercise training are conditioned by glycogen availability: A proteomics-based analysis in the McArdle mouse model. J. Physiol. 2018, 596, 1035–1061. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Ma, Q.; Xie, W.; Liou, C.L.; Zhang, D.; Sweat, M.E.; Jardin, B.D.; Naya, F.J.; Guo, Y.; Cheng, H.; et al. CMYA5 establishes cardiac dyad architecture and positioning. Nat. Commun. 2022, 13, 2185. [Google Scholar] [CrossRef] [PubMed]
- Sarparanta, J.; Blandin, G.; Charton, K.; Vihola, A.; Marchand, S.; Milic, A.; Hackman, P.; Ehler, E.; Richard, I.; Udd, B. Interactions with M-band titin and calpain 3 link myospryn (CMYA5) to tibial and limb-girdle muscular dystrophies. J. Biol. Chem. 2010, 285, 30304–30315. [Google Scholar] [CrossRef] [PubMed]
- Kielbasa, O.M.; Reynolds, J.G.; Wu, C.; Snyder, C.M.; Cho, M.Y.; Weiler, H.; Kandarian, S.; Naya, F.J. Myospryn is a calcineurin-interacting protein that negatively modulates slow-fiber-type transformation and skeletal muscle regeneration. FASEB J. 2011, 25, 2276–2286. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Gene (Symbol) | GSE151066 FoldChange | GSE151066 padj | GSE221894 FoldChange | GSE221894 padj |
|---|---|---|---|---|
| MRPL41 | 2.409945 | 1.55E-35 | 1.518872 | 0.04875 |
| GATD3 | 2.03637 | 1.52E-27 | 1.314032 | 0.03834 |
| LOC102724023 | 2.022304 | 5.44E-32 | 1.312211 | 0.03763 |
| MDH1 | 1.952064 | 2.66E-26 | 1.443929 | 0.00106 |
| ATP5MC1 | 1.785094 | 3.31E-20 | 1.480413 | 0.02837 |
| ATP5F1B | 1.745935 | 1.59E-24 | 1.316767 | 0.03834 |
| VEGFB | 1.723092 | 1.80E-31 | 1.412254 | 0.02354 |
| COX7A2 | 1.64376 | 7.12E-23 | 1.509426 | 0.04875 |
| COQ10A | 1.640346 | 5.11E-28 | 1.482467 | 0.00106 |
| ATP5F1A | 1.636938 | 1.43E-20 | 1.265757 | 0.0299 |
| YTHDC1 | 0.772175 | 1.47E-24 | 0.831046 | 0.04875 |
| CDK5RAP2 | 0.758384 | 9.56E-20 | 0.744323 | 0.04875 |
| FOXJ3 | 0.756808 | 1.19E-18 | 0.833931 | 0.04875 |
| SGCB | 0.657016 | 1.60E-20 | 0.699308 | 0.04875 |
| CMYA5 | 0.635957 | 3.98E-19 | 0.78404 | 0.04875 |
| LIMCH1 | 0.594604 | 2.26E-37 | 0.789494 | 0.04875 |
| GPD2 | 0.559419 | 1.97E-20 | 0.694959 | 0.04875 |
| PALS2 | 0.495858 | 6.11E-19 | 0.619854 | 0.04002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iyer, P.; Asante, D.M.; Vyavahare, S.; Jin, L.T.; Ahluwalia, P.; Kolhe, R.; Kashyap, H.; Isales, C.; Fulzele, S. Transcriptional Signatures of Aerobic Exercise-Induced Muscle Adaptations in Humans. J. Funct. Morphol. Kinesiol. 2025, 10, 281. https://doi.org/10.3390/jfmk10030281
Iyer P, Asante DM, Vyavahare S, Jin LT, Ahluwalia P, Kolhe R, Kashyap H, Isales C, Fulzele S. Transcriptional Signatures of Aerobic Exercise-Induced Muscle Adaptations in Humans. Journal of Functional Morphology and Kinesiology. 2025; 10(3):281. https://doi.org/10.3390/jfmk10030281
Chicago/Turabian StyleIyer, Pranav, Diana M. Asante, Sagar Vyavahare, Lee Tae Jin, Pankaj Ahluwalia, Ravindra Kolhe, Hari Kashyap, Carlos Isales, and Sadanand Fulzele. 2025. "Transcriptional Signatures of Aerobic Exercise-Induced Muscle Adaptations in Humans" Journal of Functional Morphology and Kinesiology 10, no. 3: 281. https://doi.org/10.3390/jfmk10030281
APA StyleIyer, P., Asante, D. M., Vyavahare, S., Jin, L. T., Ahluwalia, P., Kolhe, R., Kashyap, H., Isales, C., & Fulzele, S. (2025). Transcriptional Signatures of Aerobic Exercise-Induced Muscle Adaptations in Humans. Journal of Functional Morphology and Kinesiology, 10(3), 281. https://doi.org/10.3390/jfmk10030281