1. Introduction
The Chinese sturgeon (
Acipenser sinensis) is critically listed on the IUCN Red List of Threatened Species [
1]. Recognized as a flagship species for conservation, it plays a pivotal role in indicating the ecological health of the Yangtze River [
2]. Its unique ecological significance extends to stabilizing aquatic ecosystems and supporting the abundance of numerous commercially important species [
3]. However, over the past five decades, the wild population of the Chinese sturgeon has experienced a drastic decline [
4]. In the 1970s, following intensive fishing activities, the breeding stock plummeted to a mere 1727 individuals, further decreasing to a range of 203–257 individuals between 2005 and 2007, and subsequently dwindling to only a few dozen by 2010 [
5].
Chinese sturgeons can live up to 30 years. Male and female sturgeons reach sexual maturity at the ages of 8 and 10, respectively, and they engage in reproductive migration every 2 to 3 years [
5]. The long reproductive cycle makes the natural reproductive capacity of this population especially vulnerable in the current environment of the Yangtze River. In recent years, various anthropogenic activities such as inadvertent fishing, shipping operations, pollution, abnormal temperature fluctuations [
6], endocrine disruptors [
7], noise pollution, and other environmental disturbances have emerged as significant threats to the survival and reproductive behaviors of the wild Chinese sturgeon population. These factors not only inflict physical harm upon Chinese sturgeon but also disrupt their ecological equilibrium, exacerbating conservation challenges for this species. The wild Chinese sturgeon population has experienced a rapid decline since the 20th century, particularly following the completion of the Gezhouba Dam in 1982 [
8]. As the wild population dwindles, the number of Chinese sturgeons engaging in natural spawning has significantly decreased, with no instances of spawning observed in the Gezhouba spawning area in 2013, 2015, and 2017. This alarming trend suggests that the cessation of natural reproduction for the Chinese sturgeon is imminent [
1]. Over the past four decades, approximately 5 million Chinese sturgeon larvae have been reintroduced into their native habitats as part of conservation efforts led by local governments and enterprises. However, subsequent studies and evaluations have revealed that these released larvae only account for around 10% of the total larval population in their natural environment [
9,
10], yielding minimal impact on the recovery of wild Chinese sturgeon populations. Thus, the sustenance of the wild Chinese sturgeon population still heavily relies on the natural reproductive capacities of wild adults, and presently, their survival primarily depends on the natural stocks inherited from the period preceding the closure of the Gezhouba Dam. Recent data reveals a noteworthy surge in the female-to-male ratio within these populations since the 1990s, underscoring a growing scarcity of adult male Chinese sturgeons, which typically exhibit shorter lifespans in their natural habitat [
11].
Overall, the Chinese sturgeon represents a long-lived anadromous species confronted with a dwindling population and numerous external threats. To safeguard the wild Chinese sturgeon population effectively, prioritizing the protection of existing aging individuals is imperative. However, our understanding of the physiological characteristics of Chinese sturgeon remains relatively incomplete. Hence, there is a pressing need to investigate the physiological traits of Chinese sturgeon across different age groups to inform conservation efforts effectively.
Blood parameters play a crucial role in animals’ circulatory systems, serving essential functions such as material transport, physiological regulation, and defense mechanisms [
12]. They act as parameters of species characteristics and physiological status, reflecting animal health and nutritional levels [
13], and provide valuable insights for research in animal reproductive physiology and pathology. However, there remains a significant research gap regarding the blood biochemical indices of Chinese sturgeon, particularly those relevant to rescue operations. Zhang et al. (2019) investigated the impact of temperature on the growth, hematology, and immune responses of subadult Chinese sturgeon under varied ammonia nitrogen conditions in a recirculating aquaculture system. They found that the total antioxidant capacity (T-AOC), alanine aminotransferase (ALT), and lysozyme (LZM) activities peaked (
p < 0.05) at 18 °C [
14]. As the temperature rose, oxygen consumption rates and ammonia excretion rates significantly increased. Another study assessing the effects of artificial feed versus natural food on Chinese sturgeon’s growth and blood biochemistry revealed that nearly half of the examined biochemical parameters exhibited significant seasonal fluctuations, highlighting the pronounced influence of seasons on Chinese sturgeon [
15]. Shi et al. (2006) conducted a comparative analysis of blood physiological parameters between Chinese sturgeon and Amur sturgeon, revealing profound differences between the two species [
16]. Chinese sturgeon’s blood-chemistry parameters are distinct, making it challenging to directly apply blood physiological characteristics from other sturgeon species as references.
In recent years, a series of studies have been conducted on the physiology, behavior, and growth of the Chinese sturgeon. However, a significant research gap still exists regarding the blood parameters of Chinese sturgeons across different age groups, and the scientific data necessary for the protection and rescue of the Chinese sturgeon remains incomplete. Due to interspecific and individual differences, reference values obtained from other sturgeon species or small samples are inadequate to support existing conservation efforts. This study conducted statistical analysis based on a substantial amount of sample data to establish a reference system for the blood-borne pathway health parameters of Chinese sturgeon under artificial conditions. This is of great significance for the subsequent conservation efforts aimed at wild Chinese sturgeon.
Additionally, the rescue efforts for Chinese sturgeon encounter several challenges: a low survival rate of released fry, weak reproductive capacity of artificially cultured individuals, severe gender imbalance among wild individuals, and decreasing numbers of migratory individuals [
1,
8,
9,
10,
17]. This study investigates the changes in biochemical parameters of Chinese sturgeon blood with age, assuming that the individual’s resilience to the environment is positively correlated with age. We aim to offer new perspectives for addressing the current conservation issues facing Chinese sturgeon.
2. Materials and Methods
2.1. Study Site and Experimental Fish
The study was conducted at the Jiading Temporary Base of the Shanghai Aquatic Wildlife Protection Research Centre. A total of 3017 experimental Chinese sturgeons, aged between 2 and 15 years, were individually cultured in temporary ponds. These ponds were constructed as rectangular cement slope protection soil pools measuring 35 m × 25 m × 2.5 m deep and were equipped with mechanical filters and biological filtration systems. Aquaculture practices included using impounded water drained from the Yangtze River, with 1/4–1/3 water exchange every 3 days, and oxygen supplementation via water-wheel aerator aeration. Water pre-treatment facilities encompassed filtration, sedimentation, disinfection, and temperature and salinity regulation, ensuring the stability of the physical and chemical properties of sturgeon aquaculture water. The water temperature was maintained within the optimal range for Chinese sturgeon, with sudden changes limited to a temperature difference of ±2 °C within the range of 14–26 °C. Throughout the experiment, the monthly mean water temperature ranged from 13.6 to 26.5 °C, and dissolved oxygen levels ranged from 7.42 to 8.93 mg/L.
The physical-chemical parameters of water in the aquaculture environment are listed in
Table 1. In particular, the mean water temperature of the indoor tank increased slowly from February (13.6 °C) to reach a peak in August (26.5 °C), and then decreased to November (18.1 °C) (
Figure 1).
The experimental sturgeons were fed three times a day with a commercial pellet diet by Ningbo Tianbang Feed Co., Ltd. (Ningbo, China), based on 1.0% body weight. These experimental fish were the first generation of Chinese sturgeon artificially bred between 1999 and 2008. The experiment was conducted in August 2014; sampling encompassed ages ranging from 2 to 15 years, with the number of samples collected in each age group varying from 88 to 415 individuals. The culture density was controlled to remain under 7.5 individuals per 100 m3.
2.2. Blood Sample Collection and Pretreatment
Capture: The feeding of the experimental subject was stopped for more than 24 h before sampling to avoid the interference of feeding activities on physiological and biochemical parameters. The capture of Chinese sturgeon was based on the principle of minimizing the time spent leaving the water, reducing stress, and avoiding injury. After the Chinese sturgeon was captured on a stretcher and stabilized, blood samples were collected. Covering the eyes of Chinese sturgeon with wet towels helps Chinese sturgeon enter a stable state quickly.
Sampling: Based on the structural characteristics of the circulatory system in the Chinese sturgeon, arterial blood sampling was performed beneath the base of the pelvic fin near the caudal peduncle. During sampling, the Chinese sturgeon was maintained in a dorsal position with its head submerged in water to ensure normal respiration, while the tail was slightly elevated above the water’s surface. A sterile disposable syringe was used to vertically extract blood along the ventral midline direction beneath the base of the pelvic fin near the caudal peduncle. Blood collection should be performed at a steady rate to avoid aspiration of air and mucus, which may lead to hemolysis. Blood sampling should be performed immediately after capturing the fish. In this method, the damage to the fish during sample collection was minimized; hence, anesthesia was not required during blood sampling.
2.3. Sample Storage
The collected blood samples were immediately transferred into 0.5 mL EDTA anticoagulant centrifuge tubes (containing 3% EDTA-2K anticoagulant solution, with 0.2 mL added to a 5 mL centrifuge tube) and dried under temperatures ranging from 37 to 50 °C. Each treatment group had at least 9 replicates, and the samples were stored in darkness to prevent light-induced degradation for blood cell classification counting. The remaining blood samples were divided into clean 2 mL centrifuge tubes and left at room temperature without exposure to light for 4 h. After clotting and stratification, the blood was centrifuged at 4 °C and 4000 rpm for 10 min using a tabletop micro refrigerated centrifuge (Microfuge 22R, Beckman, Brea, CA, USA). The supernatant was then transferred to a clean centrifuge tube using a pipette gun. Samples intended for routine biochemical index detection could be stored in a 4 °C storage box and analyzed within 24 h, while the remaining samples were stored in a −80 °C freezer for subsequent testing, to be completed within 1 week.
In this experiment, the MS9-5 automatic hematology analyzer, Melet Schloesing Laboratoires, France, was used for blood cell classification and counting, and the reagents used were the reagent kits accompanying the instrument. Each individual was counted twice, and the average value was taken.
The Vito Selectra_E-type automatic biochemical analyzer (Vital Scientific, Dieren, The Netherlands) was utilized to assess various biochemical parameters of the samples at 37 °C, including alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), lactate dehydrogenase (LDH), total protein (TP), albumin (ALB), total bilirubin (TBIL), total bile acid (TBA), glucose (GLU), urea (UREA), creatinine (CREA), triglyceride (TG), high-density lipoprotein (HDL), low-density lipoprotein (LDL), amylase (AMY), uric acid (UARE), C-reactive protein (CRP), and total cholesterol (CHOL). The detection reagents used were the Hitachi 7020 ready-to-use full liquid packaged biochemical kits manufactured by Fudan Zhangjiang Biotechnology Shanghai Co., Ltd., (Shanghai, China) calibrated with Landau cal2350 composite calibration serum, and quality controlled with Landau quality control products. Serum potassium (K+), sodium (Na+), chlorine (Cl−), calcium (Ca2+), and pH were analyzed using the Xd685 electrolyte analyzer (Schindler, Shanghai, China), while serum osmolality (Osmol) was measured with the Vapro 5520 dew point osmometer (Wescor, Logan, UT, USA).
Samples designated for sex steroid hormone detection were flash-frozen in liquid nitrogen and stored in a −80 °C freezer. The contents of T and E2 were determined by competitive radioimmunoassay (RIA). The principle was that the serum antigen (E2 or T) and the quantitative radioactive antigen (125I-E2 or 125I-T) were competitively combined with a limited amount of antibody to form an antigen-antibody complex. The radioactivity of the complex was determined, and the content of E2 and T in serum was calculated by drawing a standard curve. The content of 11-KT was determined by a competitive enzyme-linked immunosorbent assay (Elisa). The principle was that the serum antigen (11-KT) and enzyme-labeled antigen (11-KTAChe) were combined with specific antibodies to form an antigen-antibody complex. The absorbance (412 nm) of the complex was determined by substrate color reaction, and the content of 11-KT in serum was calculated by drawing a standard curve.
2.4. Statistical Analysis
The experimental data were organized, plotted using Excel 2010 software, and statistically analyzed using SPSS 25.0 and Origin 2023 software. Results were presented as mean ± standard deviation. Differences between different age groups were assessed using a one-way ANOVA, with significant differences determined by a post hoc Duncan multiple comparison test (
p < 0.05). Prior to analysis, abnormal individual data (including diseased individuals, abnormal individuals, individuals with ingested foreign bodies in the stomach, and individuals who died within 1 year after sampling) were excluded. The mean was normalized (the formula is as follows), and relevant physiological parameters were grouped and visualized as line graphs in Excel. Parameters were examined for correlation with age, excluding those with insignificant correlation (>0.001), and those with highly significant correlation (<0.001) were subjected to Principal Component Analysis (PCA) using Origin 2023.
In order to integrate the data as much as possible and make the graphs concise, we used data normalization for the graphs, which makes the original data significance as well as the error no longer meaningful. See the original data plots in the
Supplementary Material for specific data significance.
4. Discussion
Transaminases, which are widely distributed in organisms, play a crucial role in catalyzing the exchange of ketone groups and amino groups of amino acids, representing a vital enzyme reaction for intermediate metabolism in fish. Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) serve as parameters reflecting liver and gallbladder function to a certain extent, thereby holding significance in diagnosing the overall health status of the body [
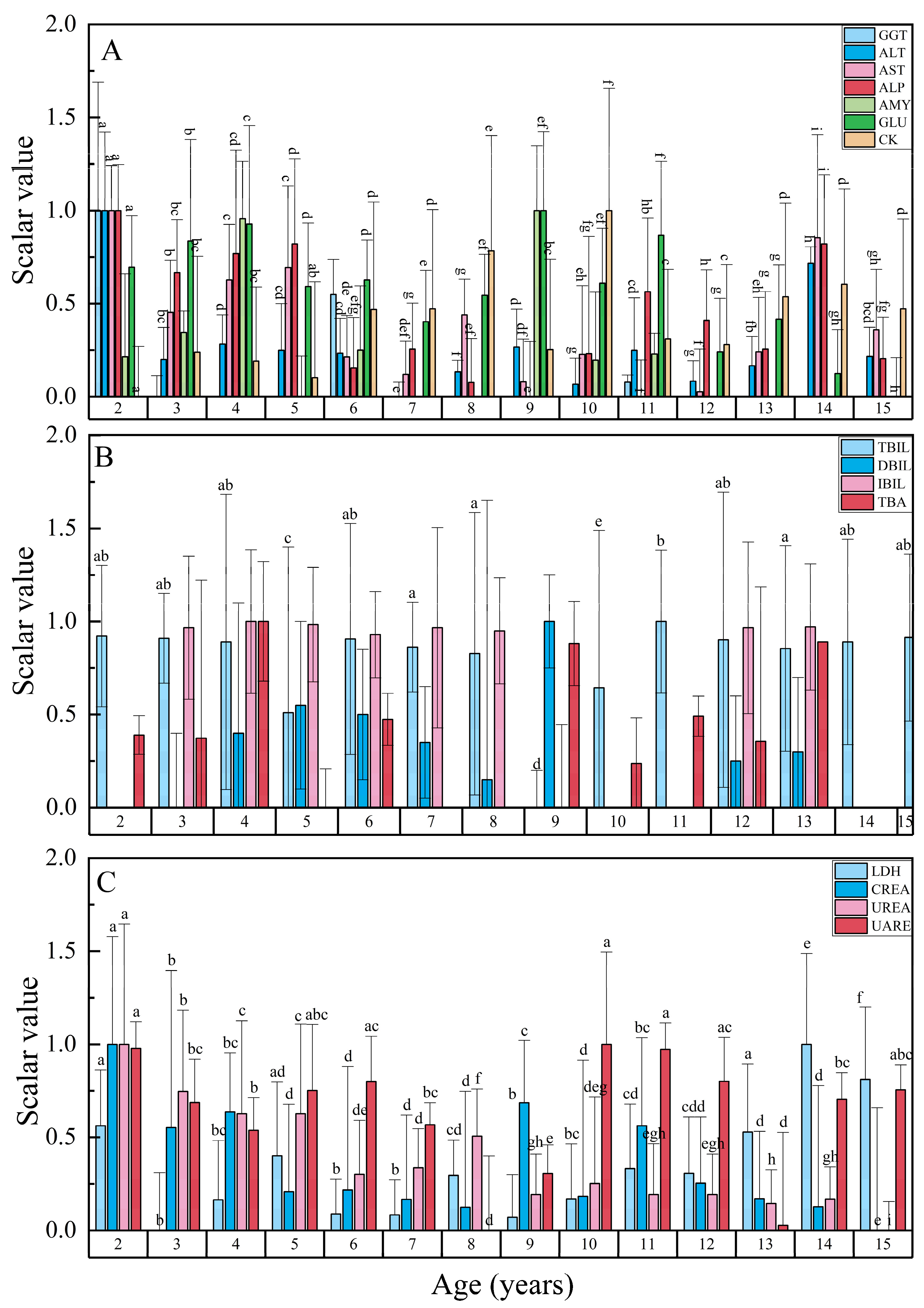
18]. Regarding liver function and bile-related parameters, ALT, AST, alkaline phosphatase (ALP), and gamma-glutamyl transpeptidase (GGT) are indicative of liver cell damage, typically exhibiting elevated levels upon liver cell injury. Notably, significant differences in ALT and AST levels were observed among different age groups of Chinese sturgeon, with the highest concentrations found in 2-year-old individuals compared to other age groups (
p < 0.05). Furthermore, total bilirubin (TBIL), direct bilirubin (DBIL), and indirect bilirubin (IBIL) reflect liver secretion and excretion function, with abnormalities in these parameters often signaling hepatobiliary system obstruction or impaired liver function. Within the bile-related parameter study, the concentrations of TBIL, DBIL, IBIL, and total bile acid (TBA) remained relatively stable across age groups except for age 9, where notable fluctuations were observed. This anomaly coincides with the age at which male Chinese sturgeons first reach gonadal maturity and initiate migration. Therefore, apart from potential environmental influences, abnormalities in these parameters at age 9 may suggest physiological disturbances stemming from inadequate culture conditions in preparation for migration.
In the kidney function-related group, the UREA levels in 15-year-old Chinese sturgeons were notably lower compared to other age groups (
p < 0.05), with a consistent decreasing trend observed in UREA levels across successive years. Moreover, the cholesterol (CHOL) levels in 14-year-old Chinese sturgeons were significantly higher than in other age groups (
p < 0.05). Conversely, no significant difference in cholesterol (CHOL) levels was observed among Chinese sturgeons aged 2–11 years (
p > 0.05). Notably, cholesterol levels exhibited a gradual increase in Chinese sturgeon aged 10–14 years over successive years. Zhang et al. (2004) previously reported significant differences in blood urea nitrogen, cholesterol, aspartate aminotransferase, and other biochemical parameters between juvenile and adult
Epinephalus coioides [
19], likely attributed to variations in hormone and metabolic levels between juvenile and adult fish.
For the immune-related group, total protein comprises albumin and globulin, reflecting liver synthesis and reserve function. Plasma total protein (TP), albumin (ALB), globulin (GLO), and the albumin-to-globulin ratio (A/G) are indicative of hepatic synthesis capacity. ALB is mainly synthesized and released by hepatic parenchymal cells, while GLO is produced by the hepatic endothelial system. A decline in hepatic synthesis function leads to alterations in TP, ALB, and GLO levels, with the extent of change correlating with the degree of impairment [
20]. ALB, a multifunctional protein, regulates osmotic pressure and acts as a transporter and storage agent. It plays a vital role in maintaining plasma osmotic pressure and fluid distribution within the body. Conversely, GLO serves an immune function, reflecting the body’s resistance. Healthy fish typically maintain normal levels of blood globulin. Various stressors, such as chronic starvation, parasitic infections, and exposure to toxic compounds, have been shown to adversely affect fish globulin [
18,
21,
22]. In the context of immune-related studies, all parameters exhibit an age-related increase, suggesting that the Chinese sturgeon’s immune capacity and visceral synthesis and storage functions improve with age. This indicates an enhancement in the organism’s resilience as it ages.
Meng et al. (2009) conducted a comparative study on the muscle composition and blood indexes of
Salvelinus malma at different ages [
23]. Significant differences were observed in various blood physiological and biochemical indexes of
Salvelinus malma at different ages, except for the serum total protein (TP), albumin (ALB), and globulin (GLO) contents (
p > 0.05). In our study, the total protein content in 14-year-old Chinese sturgeon was notably higher than in other age groups, exceeding 40 g/L (
p < 0.05), while in 2-year-old Chinese sturgeons, it was significantly lower than in other age groups (
p < 0.05). Total protein levels exhibited an increasing trend with age. The albumin content of 14-year-old Chinese sturgeons was significantly higher compared to other age groups (
p < 0.05), with a gradual increase observed in 2 to 5-year-old sturgeons over the years. Moreover, the globulin content of 14-year-old Chinese sturgeons surpassed that of other age groups, exceeding 20 g/L (
p < 0.05), while 9–12-year-old sturgeons showed a yearly increase in globulin levels. Overall, the total protein, albumin, and globulin levels in
Acipenser sinensis increased with age, indicating a gradual enhancement in immune capability. This suggests that the internal environment of the experimental
Acipenser sinensis remained relatively stable under artificial culture conditions, consistent with the majority of fish species. Simultaneously, it implies that the parameters of the artificial culture environment are reasonably set.
Serum electrolytes are vital for diagnosing fish health, with potassium (K
+), sodium (Na
+), chloride (Cl
−), and calcium (Ca
2+) playing pivotal roles in maintaining electrolyte and osmotic pressure balance. When fish metabolism is normal, ion concentrations in the body remain stable at optimal levels. Any deviations indicate metabolic abnormalities, potentially affecting fish well-being [
24]. Results showed non-significant variations in Na
+, Cl
−, and Ca
2+ levels across different ages of Chinese sturgeon, suggesting maintenance of electrolyte balance and normal metabolism during growth under artificial culture conditions. This aligns with Zhu et al.’s (1985) findings on consistent serum electrolyte and urea nitrogen changes in
Ctenopharyngodon idella [
25], emphasizing the importance of stable ion concentrations under normal metabolic conditions.
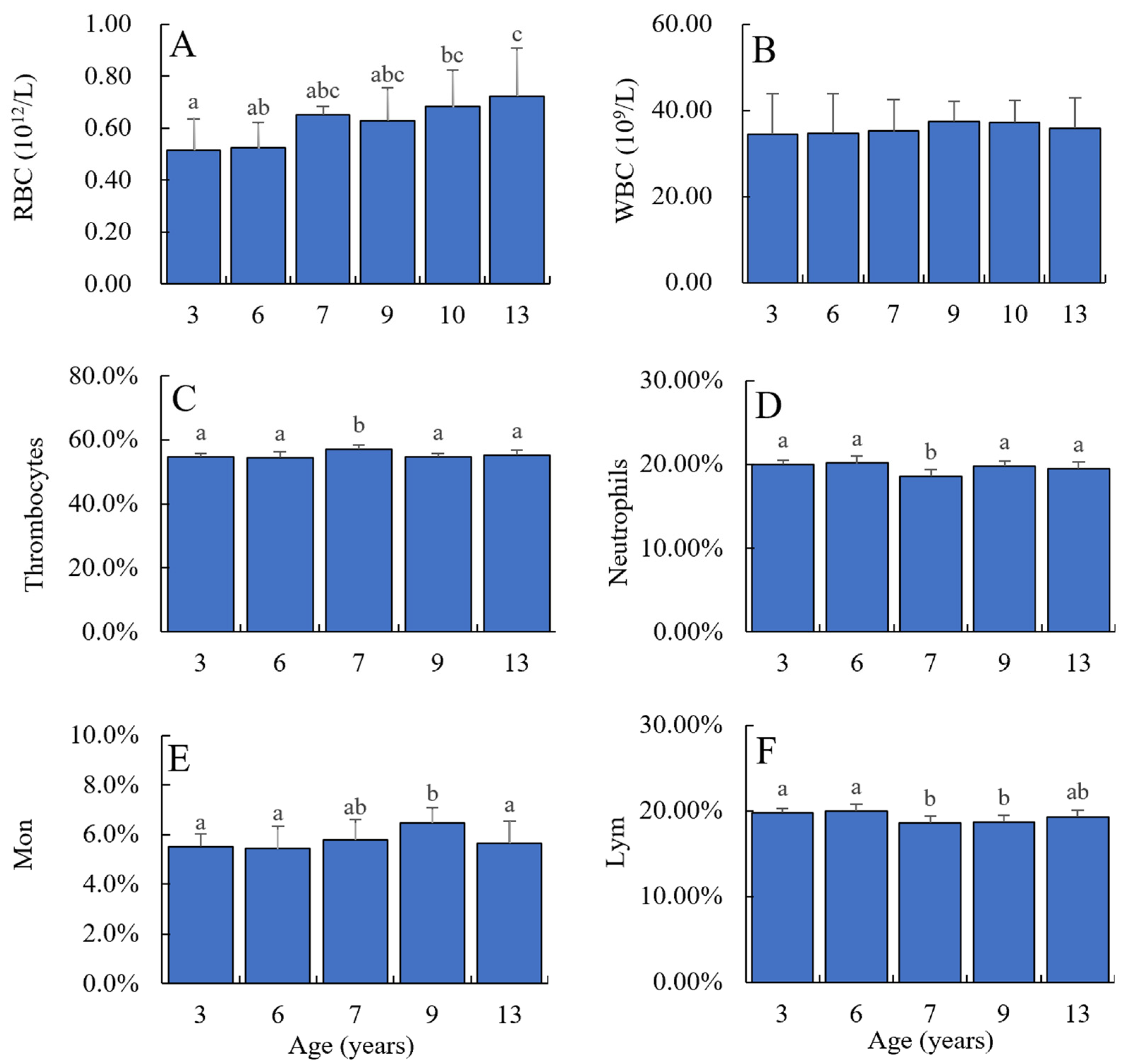
Changes in erythrocyte and leukocyte counts in the blood have been utilized as parameters of pollution and physiological dysfunction in environmental and aquaculture studies [
26]. Differences in blood cell parameters among different age groups of Chinese sturgeon serve as an index for assessing their health status. In this study, the red blood cell counts of Chinese sturgeon increased with age, with no significant difference observed between Chinese sturgeon aged 7 and 13 years. Zhu et al. (1985) demonstrated that the number of red blood cells in fish increases with age [
25]. As fish mature, the number of red blood cells stabilizes due to factors such as incomplete development of blood circulation organs in young fish and increased oxygen demand with growth, prompting an increase in red blood cell count to meet oxygen requirements [
25]. Conversely, there was no significant difference in the counts of leukocytes, thrombocytes, neutrophils, monocytes, lymphocytes, eosinophils, and basophils across all age groups of Chinese sturgeon. Changes in leukocyte count reflect alterations in fish immune function. The experimental findings indicate that the leukocyte count remains relatively stable with increasing age in Chinese sturgeon. This deviation from trends observed in other fish species may warrant further investigation and could potentially be attributed to the small sample size.
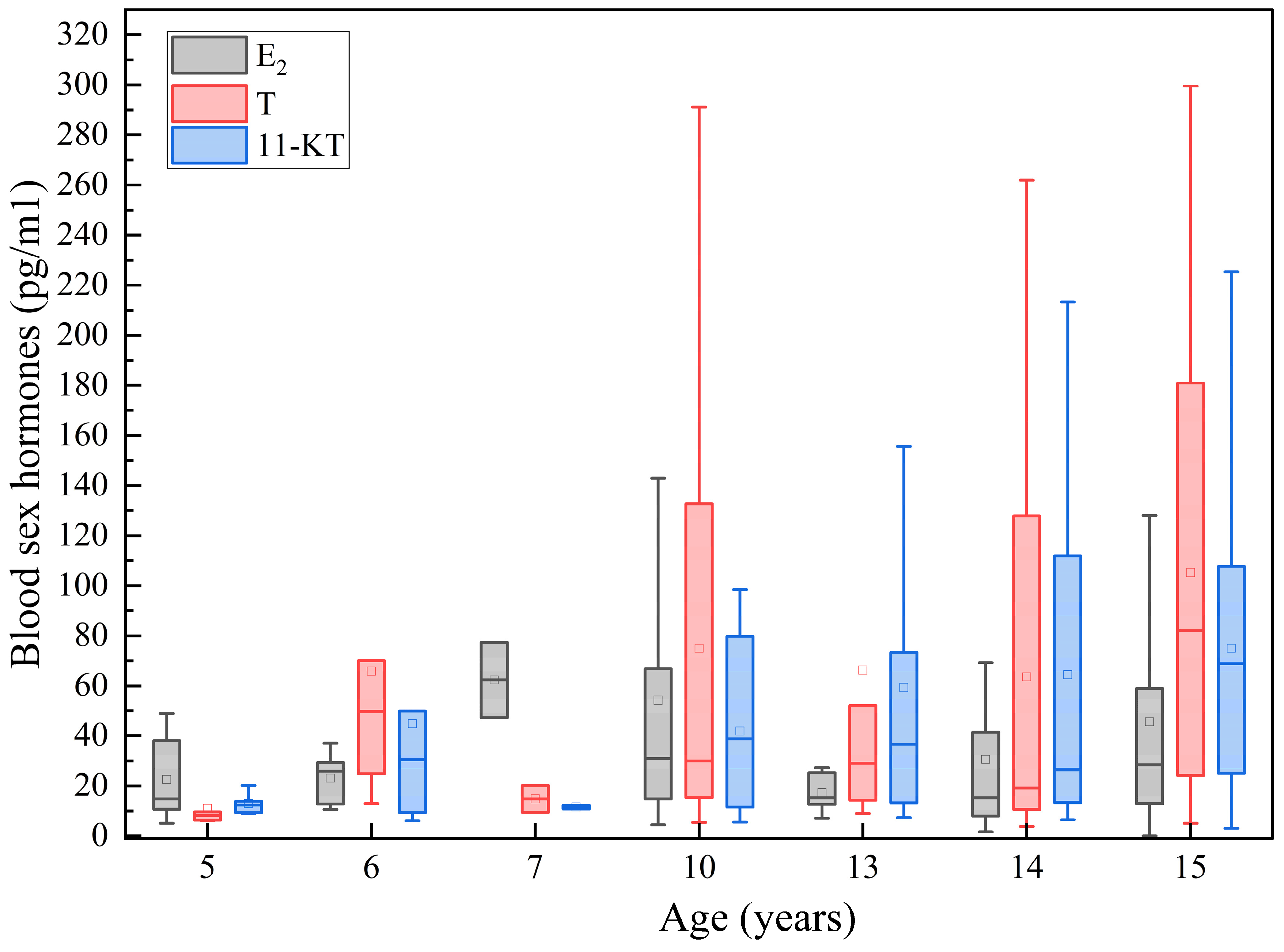
Sex steroid hormones play a pivotal role in regulating physiological processes such as individual development, gonadal maturation, and reproduction. The equilibrium of serum neutral steroid synthesis, degradation, metabolism, and excretion is crucial for maintaining homeostasis [
27]. These hormones encompass androgens, including testosterone and 11-ketotestosterone, with the former exhibiting substantial secretion and activity while the latter primarily contributes to testicular development. Additionally, estrogens like estradiol and estrone, with estradiol being the most potent, are also integral components [
28]. A radioimmunoassay was employed to track fluctuations in 17β-estradiol (E
2) and testosterone (T) levels during early Chinese sturgeon development. The results revealed minor fluctuations in E
2 levels during embryonic development, with T levels displaying a downward trajectory except for an initial increase in early embryonic stages. Following larval hatching, E
2 levels gradually rose, while T levels exhibited a temporary decline followed by a gradual increase [
29]. Previous research by Zhang and Qin demonstrated the promotion of gonadal development by testosterone and 11-ketotestosterone in sturgeon [
30,
31]. Consistently, this study found significant age-related increases in both hormones. Notably, the testosterone content was notably lower in 5-year-old sturgeon compared to 10-year-old and 15-year-old specimens, while 11-ketotestosterone levels peaked in the latter group. Conversely, estradiol levels did not significantly vary across different age groups of Chinese sturgeon. Zhang’s observations on gonadal development and related physiological parameters in cultured Chinese sturgeon further support these findings. Radioimmunoassay measurements of serum neutral steroid hormones in cultured and wild mature Chinese sturgeons revealed substantial fluctuations in E
2 levels among different ages, with a less discernible pattern, while T levels increased with age, consistent with the current study’s results. Nonetheless, widening confidence intervals with age (
Figure 5) suggest that some captive-bred individuals, despite being healthy and devoid of anomalies, exhibit underdeveloped gonads post-sexual maturity, likely due to the absence of natural conditions conducive to gonadal maturation in captivity.
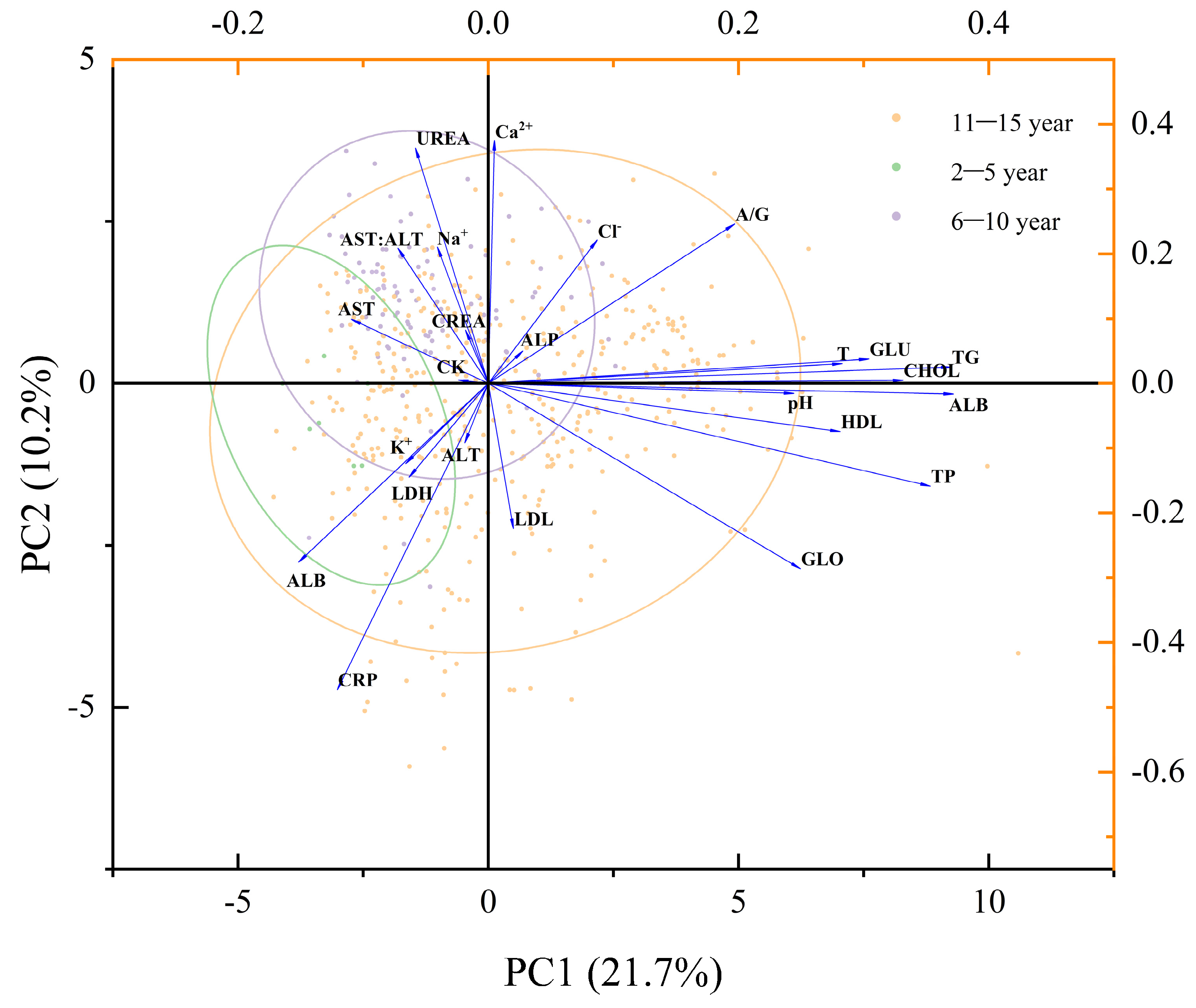
In this study, Chinese sturgeons of various ages were categorized into the juvenile stage (2–5 years of age), subadult stage (6–9 years of age), and adult stage (10–15 years of age), and their blood physiological indexes were subjected to principal component analysis (PCA). However, the analysis results revealed that the distinctions among the three age groups were not significant, with multiple components influencing the parameters (PC1 + PC2 = 31.7) (
Figure 6). This suggests that, generally, the disparities in blood biochemical indices among sturgeons of different ages are not substantial. In alignment with the outcomes of specific investigations, the disparities between the diverse age groups primarily manifest in certain parameters linked to growth and reproduction.
The serum chemistry profiles of fish can be influenced by a multitude of factors, both biological and environmental, including age, sex, water temperature, seasonal variations, and feeding conditions [
31]. Alterations in observed values of biochemical and physiological parameters often reflect the responses of organisms to changes in their surroundings, thereby serving as valuable parameters for a comprehensive assessment of organismal health across species. Baseline values may offer insights into how organisms have adapted to their local environments over evolutionary timescales.
The findings of this study revealed that the fundamental physicochemical parameters (Osm, K
+, Na
+, Cl
−, Ca
2+, pH) in the blood of 2–15-year-old Chinese sturgeons remained relatively stable under captive conditions. Consistent blood cell counts (WBC, thrombocytes, Neu, Lym, and Mon) further suggest robust sample health. The progressive increase in blood globulin content among captive Chinese sturgeon with age, spanning the 2 to 15-year age range, implies an augmentation of fish immunity over time. Notably, ALT and AST are recognized markers of liver impairment [
32], while UREA and CREA serve as parameters of gill or kidney dysfunction [
32,
33]. The significantly higher transaminase levels observed in the blood of 2-year-old sturgeons, coupled with the progressively declining UREA and CREA levels with age, may signify a transition from a more delicate and susceptible developmental stage to a more resilient physiological stage as Chinese sturgeon mature. This suggests that the age of 2 may represent a pivotal physiological developmental milestone in the life cycle of the Chinese sturgeon, underscoring the need for further investigation in future research endeavors.
The Chinese sturgeon is an anadromous species, meaning it migrates from freshwater to the sea for spawning. After hatching, young Chinese sturgeons born in the upper reaches of the Yangtze River need to swim downstream to the sea for further growth and development, a journey that typically takes about a year. The age at which Chinese sturgeons reach sexual maturity varies, with females maturing between 14 and 26 years (averaging at 18 years) and males between 9 and 18 years (averaging at 12 years). During this migration and growth process, the mortality rate among juvenile sturgeons is notably high. According to records, since the successful artificial breeding of Chinese sturgeon began in 1983, a total of 7,669,200 offspring have been released into the wild. However, the majority of these releases consisted of fry, with only 1,448,000 juveniles and larger specimens (TL > 10 cm) being released [
34]. From 1997 to 2000, the impact of releasing 100,000 juvenile sturgeons into the Yangtze River estuary was evaluated using physical and genetic markers, accounting for approximately 1–5% of the natural reproductive population replenishment that year [
34]. This assessment highlighted the inefficiency of the longstanding practice of artificially stocking Chinese sturgeons.
However, merely releasing larger juvenile individuals does not suffice as a solution to the current conservation challenges facing the Chinese sturgeon. A longstanding oversight in the artificial breeding of Chinese sturgeon has been the neglect of seawater processes. Research has revealed that adult Chinese sturgeon raised in freshwater exhibit distinct drawbacks, including noticeably smaller size (typically less than half the weight of wild brood stock), delayed sexual maturity, a reduced proportion of sexually mature individuals, and diminished reproductive quality (characterized by a smaller number of mature eggs and decreased sperm motility). These factors contribute to low spawning, fertilization, and hatching rates, as well as low fry survival rates [
35].
Based on our experimental findings, the physiological parameters of blood in Chinese sturgeon aged 2–15 years gradually stabilized with increasing age. This indicates enhancements in their blood’s oxygen transport capacity (increasing RBC), immune function (Increasing GLO, ALB, and A/G), organ development, and overall organismal resilience over time. This also confirms our hypothesis: the individual’s resilience to the environment is positively correlated with age. These findings bear significant implications for the enhanced survival of juvenile fish within the Yangtze River basin and its estuarine environment, where various detrimental factors such as noise pollution, shipping activities, and pollutant discharge prevail [
6,
7]. Hence, adjusting the current release strategy to prioritize larger juvenile Chinese sturgeon (aged 2 years or older) for release post-seawater acclimatization can be an effective approach to more efficiently protecting the Chinese sturgeon population.
Further research on the basic physiological characteristics of Chinese sturgeon, especially the basic physiology of breeding age sturgeon under captive conditions, and the resistance, adaptability, and immune capability of Chinese sturgeon juveniles reared in artificial ponds and subjected to seawater acclimatization and diverse feeding, can provide a more in-depth theoretical basis for the breeding and conservation efforts of Chinese sturgeon.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}