
Multi-Scale Coral Reef and Seascape Habitat Variables Combine to Influence Reef Fish Assemblages †

Abstract
1. Introduction
2. Materials and Methods
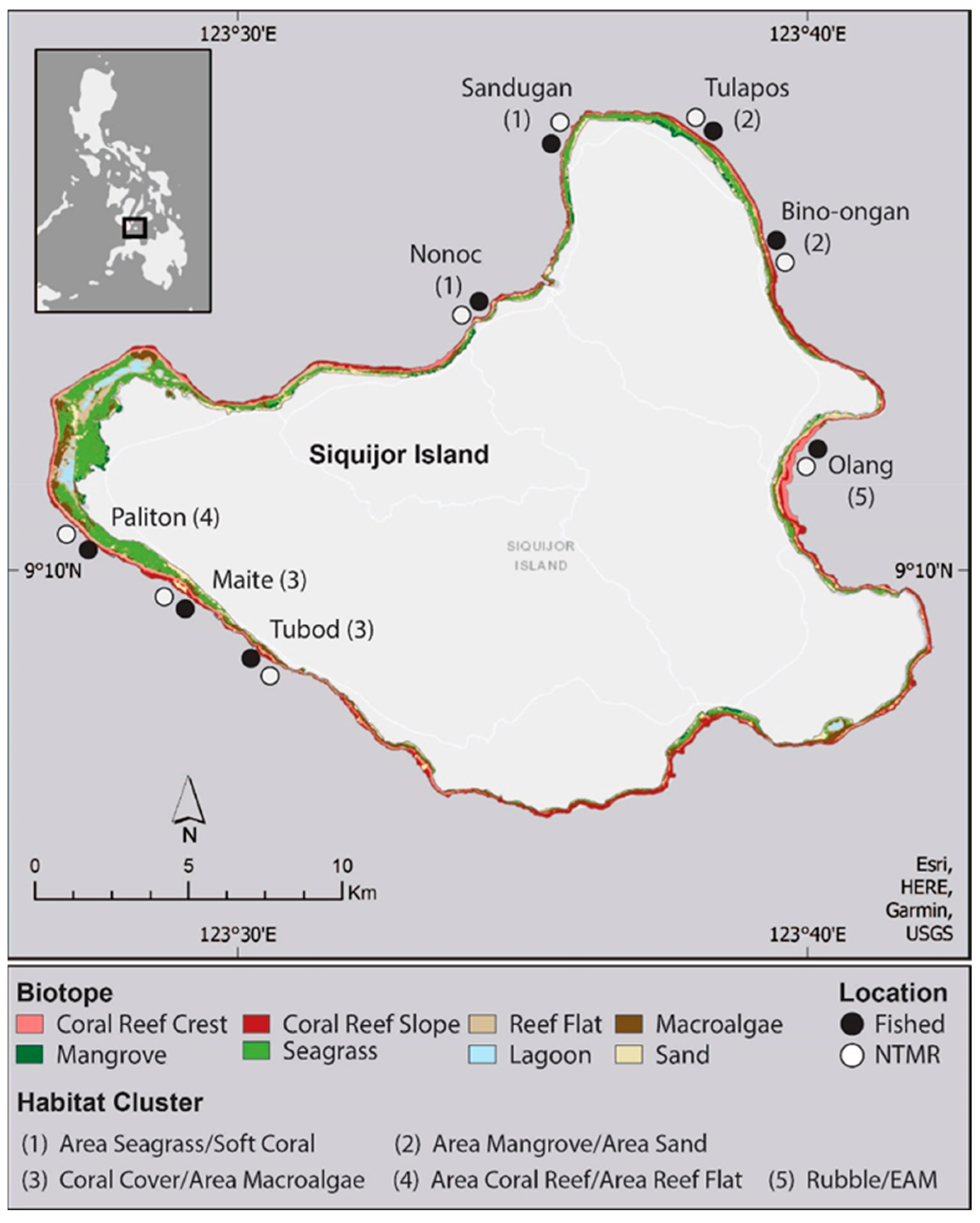
2.1. Study Area and Coral Reef Surveys
2.2. Coral Reef Surveys
2.3. Seascape-Level Spatial Analysis
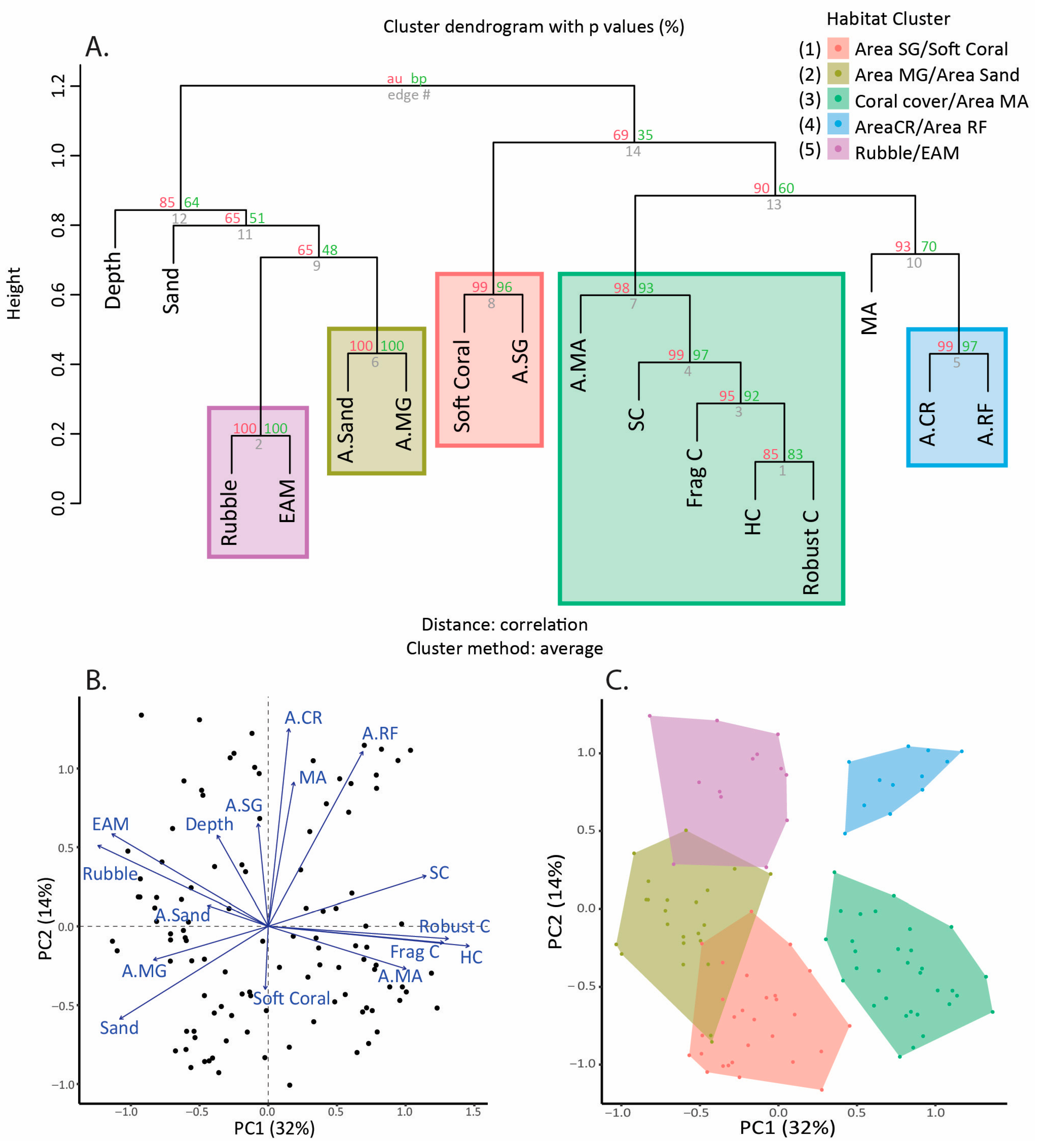
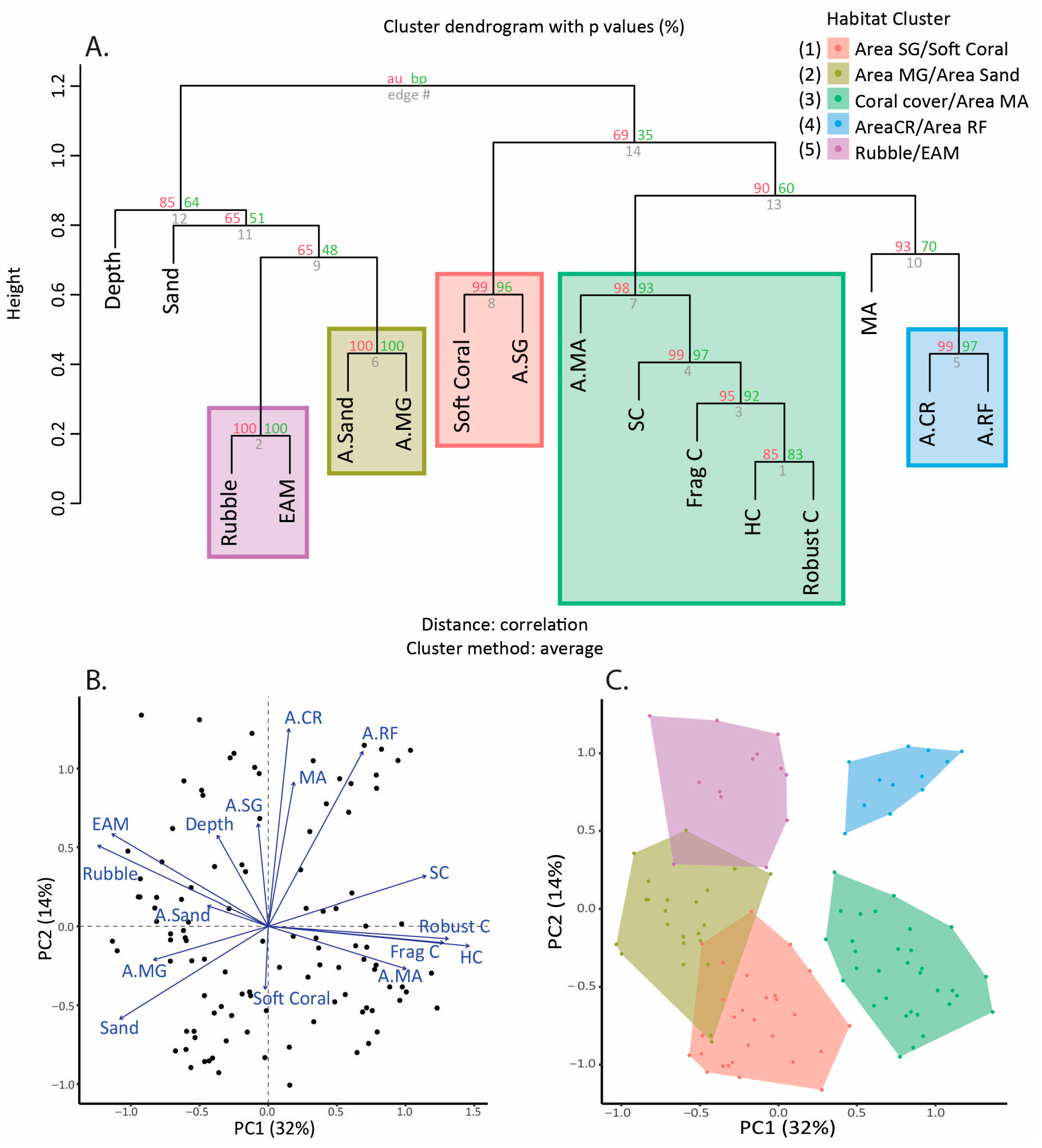
2.4. Statistical Analysis
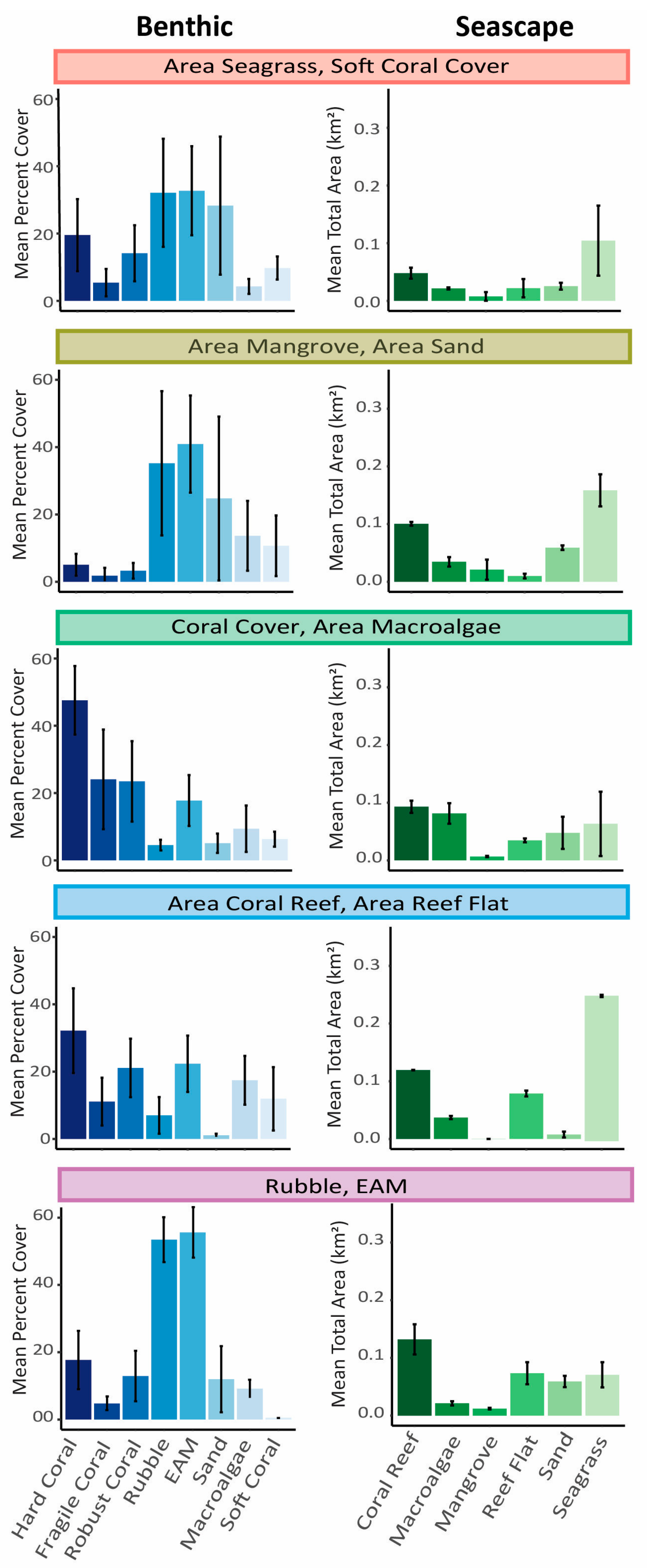
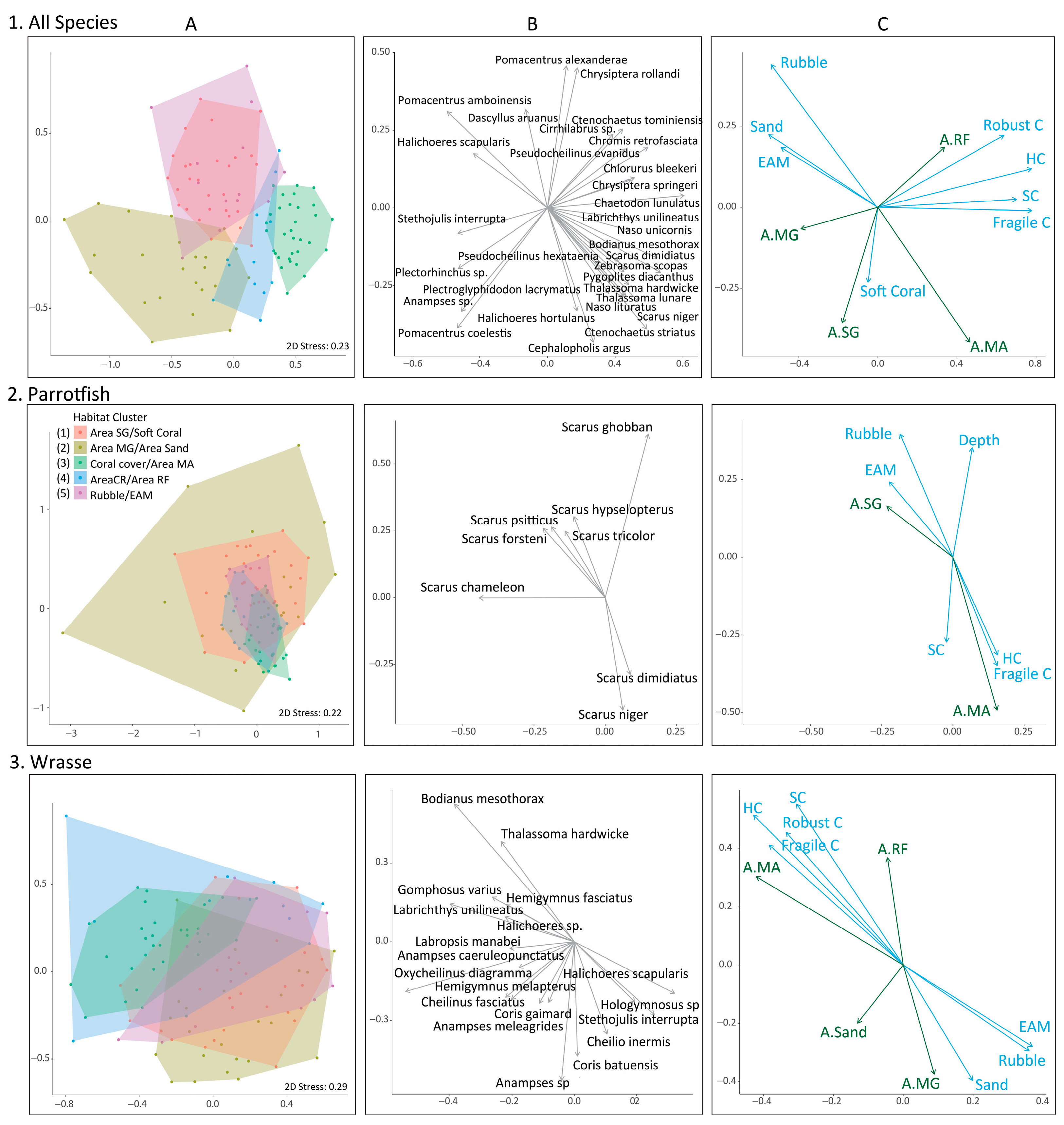
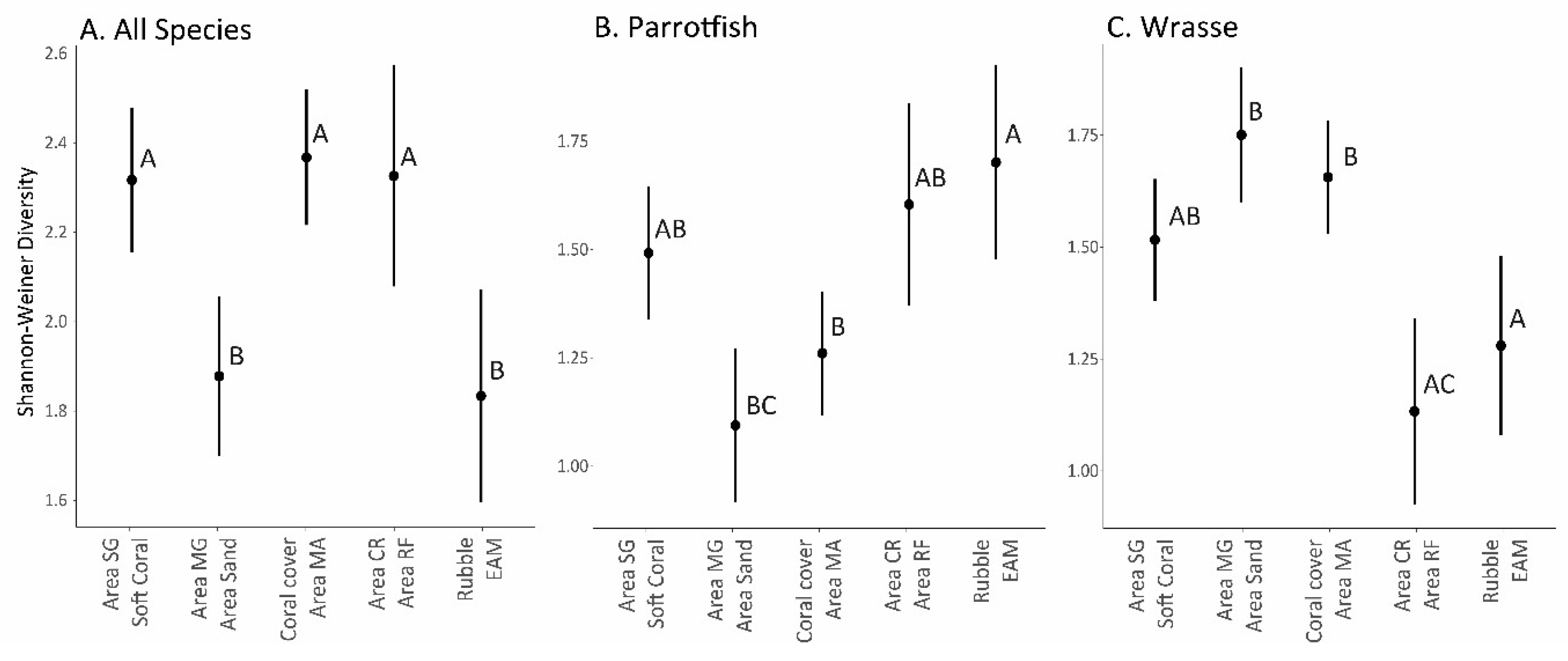
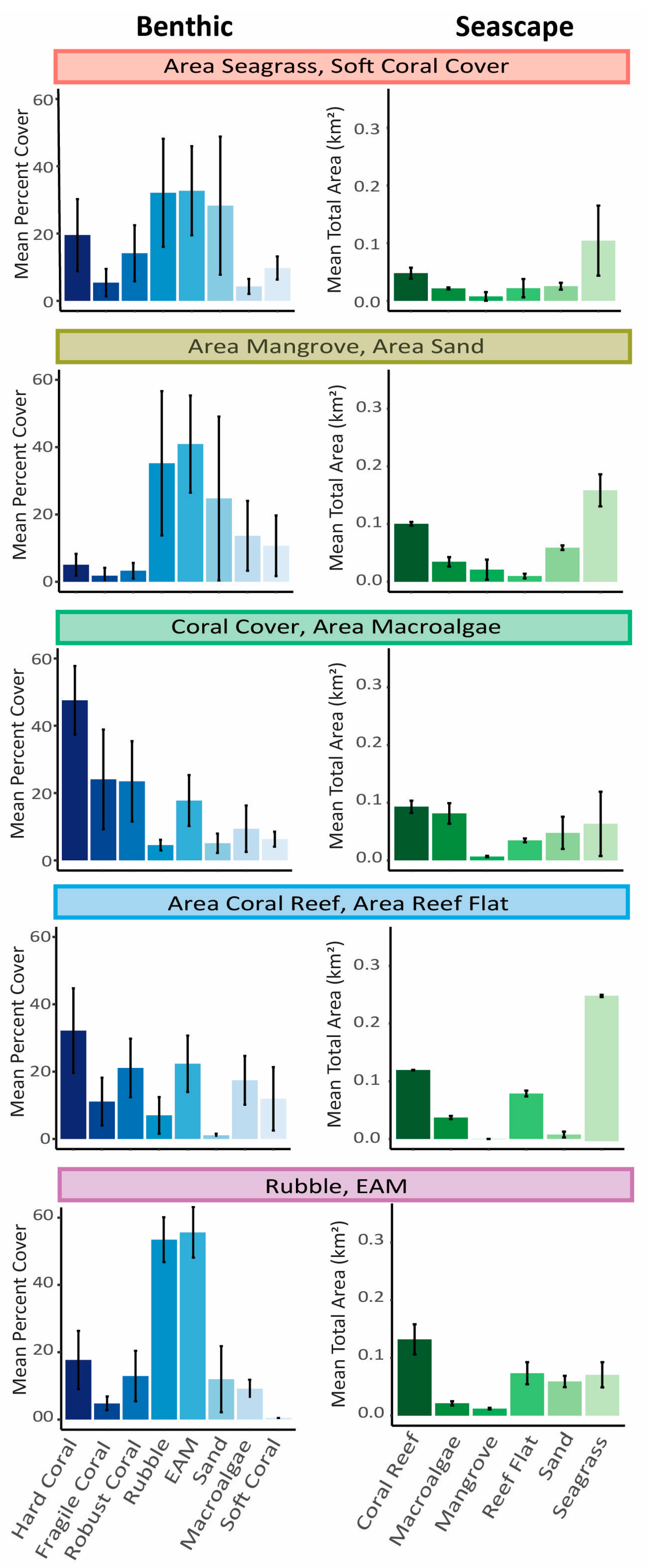
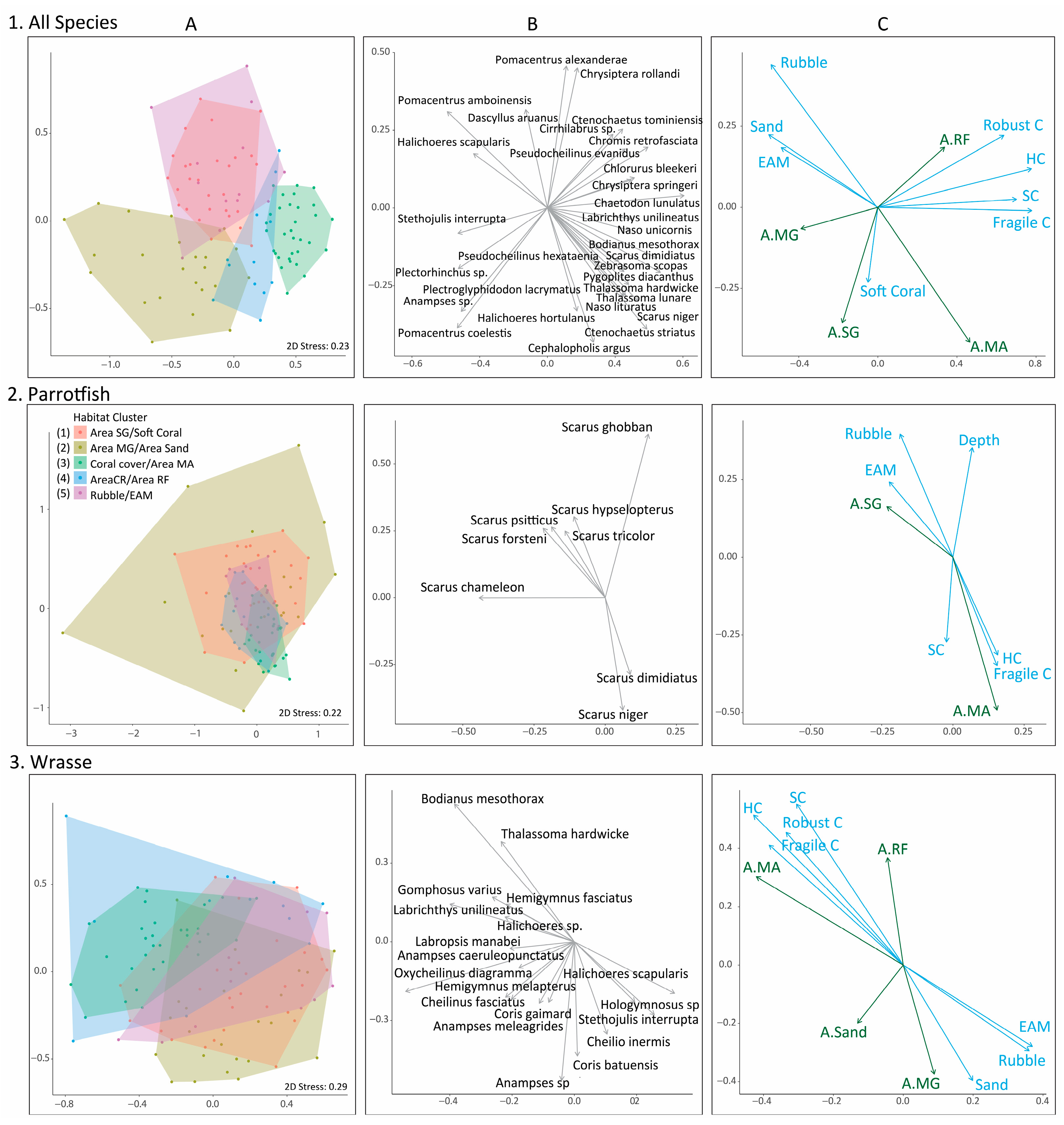
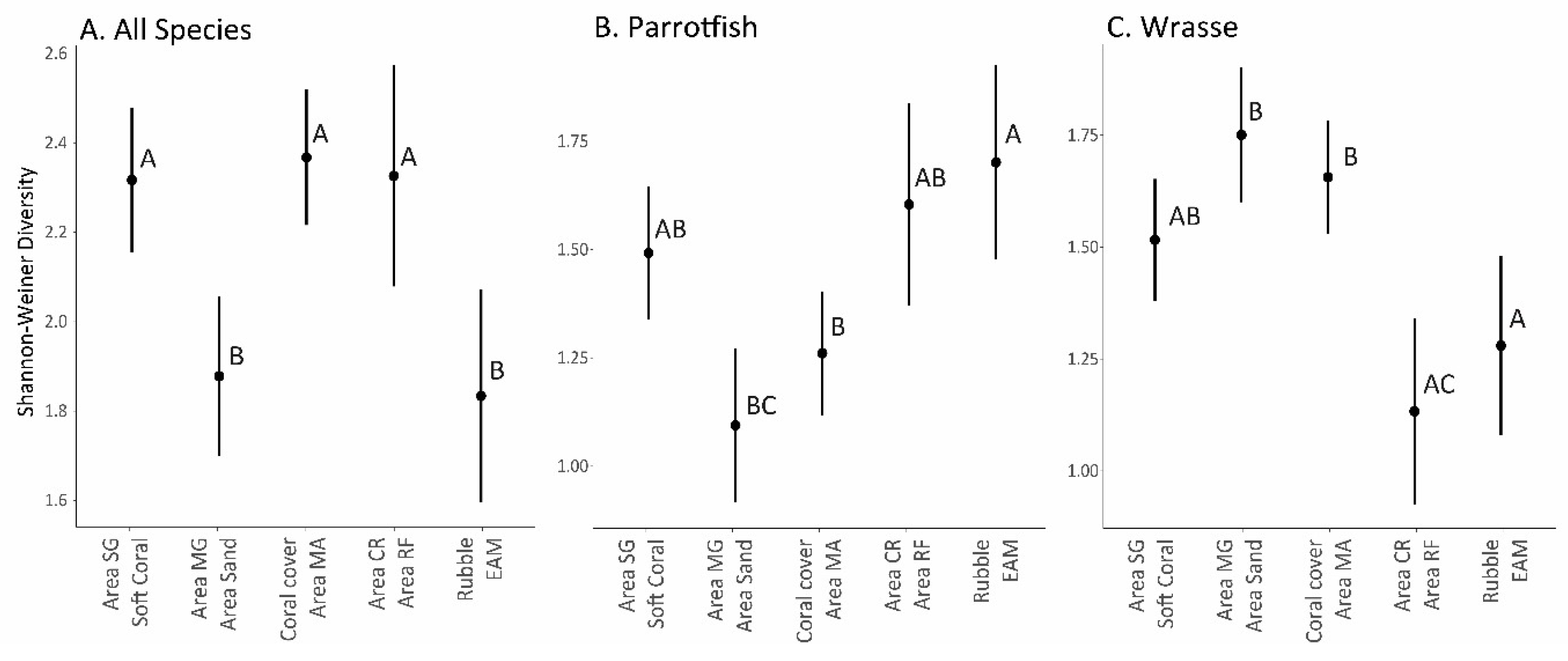
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Moilanen, A.; Nieminen, M. Simple Connectivity Measures in Spatial Ecology. Ecology 2002, 83, 1131–1145. [Google Scholar] [CrossRef]
- Tischendorf, L.; Bender, D.J.; Fahrig, L. Evaluation of Patch Isolation Metrics in Mosaic Landscapes for Specialist vs. Generalist Dispersers. Landsc. Ecol. 2003, 18, 41–50. [Google Scholar] [CrossRef]
- Ault, T.R.; Johnson, C.R. Spatially and Temporally Predictable Fish Communities on Coral Reefs. Ecol. Monogr. 1998, 68, 25–50. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Ogden, J.C.; Buckman, N.S. Movements Foraging Groups and Diurnal Migrations of the Striped Parrot Fish Scarus-Croicensis Scaridae. Ecology 1973, 54, 589–596. [Google Scholar] [CrossRef]
- Shulman, M.J.; Ogden, J.C. What Controls Tropical Reef Fish Populations: Recruitment or Benthic Mortality? An Example in the Caribbean Reef Fish Haemulon flavolineatum. Mar. Ecol. Prog. Ser. 1987, 39, 233–242. [Google Scholar] [CrossRef]
- Nagelkerken, I.; van der Velde, G.; Gorissen, M.W.; Meijer, G.J.; van’t Hof, T.; den Hartog, C. Importance of Mangroves, Seagrass Beds and the Shallow Coral Reef as a Nursery for Important Coral Reef Fishes, Using a Visual Census Technique. Estuar. Coast. Shelf Sci. 2000, 51, 31–44. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Dorenbosch, M.; Verberk, W.C.E.P.; De Morinière, E.C.; van der Velde, G. Importance of Shallow-Water Biotopes of a Caribbean Bay for Juvenile Coral Reef Fishes: Patterns in Biotope Association, Community Structure and Spatial Distribution. Mar. Ecol. Prog. Ser. 2000, 202, 175–192. [Google Scholar] [CrossRef]
- Grober-Dunsmore, R.; Pittman, S.J.; Caldow, C.; Kendall, M.S.; Frazer, T.K. A Landscape Ecology Approach for the Study of Ecological Connectivity across Tropical Marine Seascape. In Ecological Connectivity among Tropical Coastal Ecosystems; Nagelkerken, I., Ed.; Springer Science: Berlin/Heidelberg, Germany, 2009; pp. 493–530. ISBN 978-90-481-2405-3. [Google Scholar]
- Wedding, L.M.; Lepczyk, C.A.; Pittman, S.J.; Friedlander, A.M.; Jorgensen, S. Quantifying Seascape Structure: Extending Terrestrial Spatial Pattern Metrics to the Marine Realm. Mar. Ecol. Prog. Ser. 2011, 427, 219–232. [Google Scholar] [CrossRef]
- Pittman, S.J.; McAlpine, C.A.; Pittman, K.M. Linking Fish and Prawns to Their Environment: A Hierarchical Landscape Approach. Mar. Ecol. Prog. Ser. 2004, 283, 233–254. [Google Scholar] [CrossRef]
- Boström, C.; Pittman, S.J.; Simenstad, C.; Kneib, R.T. Seascape Ecology of Coastal Biogenic Habitats: Advances, Gaps, and Challenges. Mar. Ecol. Prog. Ser. 2011, 427, 191–217. [Google Scholar] [CrossRef]
- Chittaro, P.M.; Usseglio, P.; Sale, P.F. Variation in Fish Density, Assemblage Composition and Relative Rates of Predation among Mangrove, Seagrass and Coral Reef Habitats. Environ. Biol. Fishes 2005, 72, 175–187. [Google Scholar] [CrossRef]
- Mumby, P.J.; Hastings, A. The Impact of Ecosystem Connectivity on Coral Reef Resilience. J. Appl. Ecol. 2008, 45, 854–862. [Google Scholar] [CrossRef]
- Olds, A.D.; Connolly, R.M.; Pitt, K.A.; Maxwell, P.S. Primacy of Seascape Connectivity Effects in Structuring Coral Reef Fish Assemblages. Mar. Ecol. Prog. Ser. 2012, 462, 191–203. [Google Scholar] [CrossRef]
- Berkström, C.; Jörgensen, T.L.; Hellström, M. Ecological Connectivity and Niche Differentiation between Two Closely Related Fish Species in the Mangrove-Seagrass-Coral Reef Continuum. Mar. Ecol. Prog. Ser. 2013, 477, 201–215. [Google Scholar] [CrossRef]
- Pittman, S.J.; Olds, A.D. Seascape Ecology of Fishes on Coral Reefs. In Ecology of Fishes on Coral Reefs; Mora, C., Ed.; Cambridge University Press: Cambridge, UK, 2015; pp. 274–282. ISBN 9781316105412. [Google Scholar]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K.; Messmer, V.; Graham, N.A.J. Changes in Biodiversity and Functioning of Reef Fish Assemblages Following Coral Bleaching and Coral Loss. Diversity 2011, 3, 424–452. [Google Scholar] [CrossRef]
- Coker, D.J.; Wilson, S.K.; Pratchett, M.S. Importance of Live Coral Habitat for Reef Fishes. Rev. Fish Biol. Fish. 2014, 24, 89–126. [Google Scholar] [CrossRef]
- Russ, G.R.; Miller, K.I.; Rizzari, J.R.; Alcala, A.C. Long-Term No-Take Marine Reserve and Benthic Habitat Effects on Coral Reef Fishes. Mar. Ecol. Prog. Ser. 2015, 529, 233–248. [Google Scholar] [CrossRef]
- Igulu, M.M.; Nagelkerken, I.; Dorenbosch, M.; Grol, M.G.G.; Harborne, A.R.; Kimirei, I.A.; Mumby, P.J.; Olds, A.D.; Mgaya, Y.D. Mangrove Habitat Use by Juvenile Reef Fish: Meta-Analysis Reveals That Tidal Regime Matters More than Biogeographic Region. PLoS ONE 2014, 9, e114715. [Google Scholar] [CrossRef] [PubMed]
- Hemingson, C.R.; Bellwood, D.R. Biogeographic Patterns in Major Marine Realms: Function Not Taxonomy Unites Fish Assemblages in Reef, Seagrass and Mangrove Systems. Ecography 2018, 41, 174–182. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking Patch Size and Isolation Effects: The Habitat Amount Hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Berkström, C.; Gullström, M.; Lindborg, R.; Mwandya, A.W.; Yahya, S.A.S.; Kautsky, N.; Nyström, M. Exploring ‘Knowns’ and ‘Unknowns’ in Tropical Seascape Connectivity with Insights from East African Coral Reefs. Estuar. Coast. Shelf Sci. 2012, 107, 1–21. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. What Size Is a Biologically Relevant Landscape? Landsc. Ecol. 2012, 27, 929–941. [Google Scholar] [CrossRef]
- Pittman, S.J.; Brown, K.A. Multi-Scale Approach for Predicting Fish Species Distributions across Coral Reef Seascapes. PLoS ONE 2011, 6, e20583. [Google Scholar] [CrossRef]
- Johnson, A.F.; Jenkins, S.R.; Hiddink, J.G.; Hinz, H. Linking Temperate Demersal Fish Species to Habitat: Scales, Patterns and Future Directions. Fish Fish. 2013, 14, 256–280. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Dorenbosch, M.; Verberk, W.C.E.P.; Cocheret De La Morinière, E.; Van Der Velde, G. Day-Night Shifts of Fishes between Shallow-Water Biotopes of a Caribbean Bay, with Emphasis on the Nocturnal Feeding of Haemulidae and Lutjanidae. Mar. Ecol. Prog. Ser. 2000, 194, 55–64. [Google Scholar] [CrossRef]
- Appeldoorn, R.S.; Aguilar-Perera, A.; Bouwmeester, B.L.K.; Dennis, G.D.; Hill, R.L.; Merten, W.; Recksiek, C.W.; Williams, S.J. Movement of Fishes (Grunts: Haemulidae) across the Coral Reef Seascape: A Review of Scales, Patterns and Processes. Caribb. J. Sci. 2009, 45, 304–316. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Sheaves, M.; Baker, R.; Connolly, R.M. The Seascape Nursery: A Novel Spatial Approach to Identify and Manage Nurseries for Coastal Marine Fauna. Fish Fish. 2015, 16, 362–371. [Google Scholar] [CrossRef]
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The Identification, Conservation, and Management of Estuarine and Marine Nurseries for Fish and Invertebrates. Bioscience 2001, 51, 633–641. [Google Scholar] [CrossRef]
- Adams, A.J.; Ebersole, J.P. Use of Back-Reef and Lagoon Habitats by Coral Reef Fishes. Mar. Ecol. Prog. Ser. 2002, 228, 213–226. [Google Scholar] [CrossRef]
- Ogden, J.C.; Quinn, T.P. Migration of Coral Reef Fishes: Ecological Significance and Orientation Mechanisms. In Mechanisms of Migration in Fishes; Springer: Boston, MA, USA, 1984; pp. 293–308. [Google Scholar]
- Meyer, J.L.; Schultz, E.T. Migrating Haemulid Fishes as a Source of Nutrients and Organic Matter on Coral Reefs. Limnol. Oceanogr. 1985, 30, 146–156. [Google Scholar] [CrossRef]
- Harborne, A.R.; Nagelkerken, I.; Wolff, N.H.; Bozec, Y.M.; Dorenbosch, M.; Grol, M.G.G.; Mumby, P.J. Direct and Indirect Effects of Nursery Habitats on Coral-Reef Fish Assemblages, Grazing Pressure and Benthic Dynamics. Oikos 2016, 125, 957–967. [Google Scholar] [CrossRef]
- Nakamura, Y.; Horinouchi, M.; Shibuno, T.; Tanaka, Y.; Miyajima, T.; Koike, I.; Kurokura, H.; Sano, M. Evidence of Ontogenetic Migration from Mangroves to Coral Reefs by Black-Tail Snapper Lutjanus Fulvus: Stable Isotope Approach. Mar. Ecol. Prog. Ser. 2008, 355, 257–266. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Grol, M.G.G.; Mumby, P.J. Effects of Marine Reserves versus Nursery Habitat Availability on Structure of Reef Fish Communities. PLoS ONE 2012, 7, e36906. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Hughes, B.B.; Johnson, A.J.; Pfirrman, B.W.; Rasher, D.B.; Smyth, A.R.; Williams, B.L.; Beck, M.W.; Orth, R.J. Are Coastal Habitats Important Nurseries? A Meta-Analysis. Conserv. Lett. 2019, 12, e12645. [Google Scholar] [CrossRef]
- Abu El-Regal, M.A.; Ibrahim, N.K. Role of Mangroves as a Nursery Ground for Juvenile Reef Fishes in the Southern Egyptian Red Sea. Egypt. J. Aquat. Res. 2014, 40, 71–78. [Google Scholar] [CrossRef]
- Kimirei, I.A.; Nagelkerken, I.; Slooter, N.; Gonzalez, E.T.; Huijbers, C.M.; Mgaya, Y.D.; Rypel, A.L. Demography of Fish Populations Reveals New Challenges in Appraising Juvenile Habitat Values. Mar. Ecol. Prog. Ser. 2015, 518, 225–237. [Google Scholar] [CrossRef]
- Nakamura, Y.; Horinouchi, M.; Nakai, T.; Sano, M. Food Habits of Fishes in a Seagrass Bed on a Fringing Coral Reef at Iriomote Island, Southern Japan. Ichthyol. Res. 2003, 50, 15–22. [Google Scholar] [CrossRef]
- Dorenbosch, M.; Grol, M.G.G.; Christianen, M.J.A.; Nagelkerken, I.; van Der Velde, G. Indo-Pacific Seagrass Beds and Mangroves Contribute to Fish Density and Diversity on Adjacent Coral Reefs. Mar. Ecol. Prog. Ser. 2005, 302, 63–76. [Google Scholar] [CrossRef]
- Grober-Dunsmore, R.; Frazer, T.K.; Lindberg, W.J.; Beets, J. Reef Fish and Habitat Relationships in a Caribbean Seascape: The Importance of Reef Context. Coral Reefs 2007, 26, 201–216. [Google Scholar] [CrossRef]
- Davis, J.P.; Pitt, K.A.; Fry, B.; Olds, A.D.; Connolly, R.M. Seascape-Scale Trophic Links for Fish on Inshore Coral Reefs. Coral Reefs 2014, 33, 897–907. [Google Scholar] [CrossRef]
- Berkström, C.; Eggertsen, L.; Goodell, W.; Cordeiro, C.A.M.M.; Lucena, M.B.; Gustafsson, R.; Bandeira, S.; Jiddawi, N.; Ferreira, C.E.L. Thresholds in Seascape Connectivity: The Spatial Arrangement of Nursery Habitats Structure Fish Communities on Nearby Reefs. Ecography 2020, 43, 882–896. [Google Scholar] [CrossRef]
- Fulton, C.J.; Abesamis, R.A.; Berkström, C.; Depczynski, M.; Graham, N.A.J.; Holmes, T.H.; Kulbicki, M.; Noble, M.M.; Radford, B.T.; Tano, S.; et al. Form and Function of Tropical Macroalgal Reefs in the Anthropocene. Funct. Ecol. 2019, 33, 989–999. [Google Scholar] [CrossRef]
- Fulton, C.J.; Berkström, C.; Wilson, S.K.; Abesamis, R.A.; Bradley, M.; Åkerlund, C.; Barrett, L.T.; Bucol, A.A.; Chacin, D.H.; Chong-Seng, K.M.; et al. Macroalgal Meadow Habitats Support Fish and Fisheries in Diverse Tropical Seascapes. Fish Fish. 2020, 21, 700–717. [Google Scholar] [CrossRef]
- Sambrook, K.; Hoey, A.S.; Andréfouët, S.; Cumming, G.S.; Duce, S.; Bonin, M.C. Beyond the Reef: The Widespread Use of Non-Reef Habitats by Coral Reef Fishes. Fish Fish. 2019, 20, 903–920. [Google Scholar] [CrossRef]
- Sievers, K.T.; McClure, E.C.; Abesamis, R.A.; Russ, G.R. Non-Reef Habitats in a Tropical Seascape Affect Density and Biomass of Fishes on Coral Reefs. Ecol. Evol. 2020, 10, 13673–13686. [Google Scholar] [CrossRef] [PubMed]
- Eggertsen, L.; Ferreira, C.E.L.; Fontoura, L.; Kautsky, N.; Gullström, M.; Berkström, C. Seaweed Beds Support More Juvenile Reef Fish than Seagrass Beds in a South-Western Atlantic Tropical Seascape. Estuar. Coast. Shelf Sci. 2017, 196, 97–108. [Google Scholar] [CrossRef]
- Sambrook, K.; Bonin, M.C.; Bradley, M.; Cumming, G.S.; Duce, S.; Andréfouët, S.; Hoey, A.S. Broadening Our Horizons: Seascape Use by Coral Reef-Associated Fishes in Kavieng, Papua New Guinea, Is Common and Diverse. Coral Reefs 2020, 39, 1187–1197. [Google Scholar] [CrossRef]
- Huntington, B.E.; Karnauskas, M.; Babcock, E.A.; Lirman, D. Untangling Natural Seascape Variation from Marine Reserve Effects Using a Landscape Approach. PLoS ONE 2010, 5, e12327. [Google Scholar] [CrossRef] [PubMed]
- Olds, A.D.; Connolly, R.M.; Pitt, K.A.; Maxwell, P.S. Habitat Connectivity Improves Reserve Performance. Conserv. Lett. 2012, 5, 56–63. [Google Scholar] [CrossRef]
- Olds, A.D.; Albert, S.; Maxwell, P.S.; Pitt, K.A.; Connolly, R.M. Mangrove-Reef Connectivity Promotes the Effectiveness of Marine Reserves across the Western Pacific. Glob. Ecol. Biogeogr. 2013, 22, 1040–1049. [Google Scholar] [CrossRef]
- Verweij, M.C.; Nagelkerken, I.; Wartenbergh, S.L.J.; Pen, I.R.; van Der Velde, G. Caribbean Mangroves and Seagrass Beds as Daytime Feeding Habitats for Juvenile French Grunts, Haemulon flavolineatum. Mar. Biol. 2006, 149, 1291–1299. [Google Scholar] [CrossRef]
- Yabsley, N.A.; Olds, A.D.; Connolly, R.M.; Martin, T.S.H.; Gilby, B.L.; Maxwell, P.S.; Huijbers, C.M.; Schoeman, D.S.; Schlacher, T.A. Resource Type Influences the Effects of Reserves and Connectivity on Ecological Functions. J. Anim. Ecol. 2016, 85, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.S.H.; Olds, A.D.; Olalde, A.B.H.; Berkström, C.; Gilby, B.L.; Schlacher, T.A.; Butler, I.R.; Yabsley, N.A.; Zann, M.; Connolly, R.M. Habitat Proximity Exerts Opposing Effects on Key Ecological Functions. Landsc. Ecol. 2018, 33, 1273–1286. [Google Scholar] [CrossRef]
- Henderson, C.J.; Gilby, B.L.; Lee, S.Y.; Stevens, T. Contrasting Effects of Habitat Complexity and Connectivity on Biodiversity in Seagrass Meadows. Mar. Biol. 2017, 164, 117. [Google Scholar] [CrossRef]
- Sievers, K.T.; Abesamis, R.A.; Bucol, A.A.; Russ, G.R. Unravelling Seascape Patterns of Cryptic Life Stages: Non-Reef Habitat Use in Juvenile Parrotfishes. Diversity 2020, 12, 376. [Google Scholar] [CrossRef]
- Vanderklift, M.A.; How, J.; Wernberg, T.; MacArthur, L.D.; Heck, K.L.; Valentine, J.F. Proximity to Reef Influences Density of Small Predatory Fishes, While Type of Seagrass Influences Intensity of Their Predation on Crabs. Mar. Ecol. Prog. Ser. 2007, 340, 235–243. [Google Scholar] [CrossRef]
- van Lier, J.R.; Wilson, S.K.; Depczynski, M.; Wenger, L.N.; Fulton, C.J. Habitat Connectivity and Complexity Underpin Fish Community Structure across a Seascape of Tropical Macroalgae Meadows. Landsc. Ecol. 2018, 33, 1287–1300. [Google Scholar] [CrossRef]
- Evans, R.D.; Wilson, S.K.; Field, S.N.; Moore, J.A.Y. Importance of Macroalgal Fields as Coral Reef Fish Nursery Habitat in North-West Australia. Mar. Biol. 2014, 161, 599–607. [Google Scholar] [CrossRef]
- McClure, E.C.; Sievers, K.T.; Abesamis, R.A.; Hoey, A.S.; Alcala, A.C.; Russ, G.R. Higher Fish Biomass inside than Outside Marine Protected Areas despite Typhoon Impacts in a Complex Reefscape. Biol. Conserv. 2020, 241, 108354. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Polunin, N.V.C. Appraisal of Visual Assessments of Habitat Complexity and Benthic Composition on Coral Reefs. Mar. Biol. 2007, 151, 1069–1076. [Google Scholar] [CrossRef]
- Suzuki, R.; Shimodaira, H. R Package, version 1.2-2; Hierarchical Clustering with P-Values via Multiscale Bootstrap Resampling; RStudio: Boston, MA, USA, 2011. [Google Scholar]
- Jouffray, J.B.; Wedding, L.M.; Norström, A.V.; Donovan, M.K.; Williams, G.J.; Crowder, L.B.; Erickson, A.L.; Friedlander, A.M.; Graham, N.A.J.; Gove, J.M.; et al. Parsing Human and Biophysical Drivers of Coral Reef Regimes. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182544. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.; Singh, S. A Survey of Clustering Techniques. Int. J. Comput. Appl. 2010, 7, 1–5. [Google Scholar] [CrossRef]
- Gillanders, B.M.; Able, K.W.; Brown, J.A.; Eggleston, D.B.; Sheridan, P.F. Evidence of Connectivity between Juvenile and Adult Habitats for Mobile Marine Fauna: An Important Component of Nurseries. Mar. Ecol. Prog. Ser. 2003, 247, 281–295. [Google Scholar] [CrossRef]
- Kimirei, I.A.; Nagelkerken, I.; Griffioen, B.; Wagner, C.; Mgaya, Y.D. Ontogenetic Habitat Use by Mangrove/Seagrass-Associated Coral Reef Fishes Shows Flexibility in Time and Space. Estuar. Coast. Shelf Sci. 2011, 92, 47–58. [Google Scholar] [CrossRef]
- McClure, E.C.; Hoey, A.S.; Sievers, K.T.; Abesamis, R.A.; Russ, G.R. Relative Influence of Environmental Factors and Fishing on Coral Reef Fish Assemblages. Conserv. Biol. 2020, 35, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Russ, G.R.; Payne, C.S.; Bergseth, B.J.; Rizzari, J.R.; Abesamis, R.A.; Alcala, A.C. Decadal-Scale Response of Detritivorous Surgeonfishes (Family Acanthuridae) to No-Take Marine Reserve Protection and Changes in Benthic Habitat. J. Fish Biol. 2018, 93, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.M.; Russ, G.R. Response of Herbivorous Fishes to Crown-of-Thorns Starfish Acanthaster planci Outbreaks. III. Age, Growth, Mortality and Maturity Indices of Acanthurus nigrofuscus. Mar. Ecol. Prog. Ser. 1996, 136, 25–35. [Google Scholar] [CrossRef]
- Bouillon, S.; Connolly, R.M.; Lee, S.Y. Organic Matter Exchange and Cycling in Mangrove Ecosystems: Recent Insights from Stable Isotope Studies. J. Sea Res. 2008, 59, 44–58. [Google Scholar] [CrossRef]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; Mckee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological Role and Services of Tropical Mangrove Ecosystems: A Reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Randall, J.E. Acanthuridae. In Smith’s Sea Fishes; Smith, M.M., Heemstra, P.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 811–823. [Google Scholar]
- Russ, G.R.; Aller-Rojas, O.D.; Rizzari, J.R.; Alcala, A.C. Off-Reef Planktivorous Reef Fishes Respond Positively to Decadal-Scale No-Take Marine Reserve Protection and Negatively to Benthic Habitat Change. Mar. Ecol. 2017, 38, e12442. [Google Scholar] [CrossRef]
- Russ, G.R.; Alcala, A.C.; Maypa, A.P. Spillover from Marine Reserves: The Case of Naso vlamingii at Apo Island, the Philippines. Mar. Ecol. Prog. Ser. 2003, 264, 15–20. [Google Scholar] [CrossRef]
- Abesamis, R.A.; Russ, G.R. Density-Dependent Spillover from a Marine Reserve: Long-Term Evidence. Ecol. Appl. 2005, 15, 1798–1812. [Google Scholar] [CrossRef]
- Choat, J.H.; Clements, K.D.; Robbins, W.D. The Trophic Status of Herbivorous Fishes on Coral Reefs 1: Dietary Analyses. Mar. Biol. 2002, 140, 613–623. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-Shelf Variation in Browsing Intensity on the Great Barrier Reef. Coral Reefs 2010, 29, 499–508. [Google Scholar] [CrossRef]
- Bierwagen, S.L.; Price, D.K.; Pack, A.A.; Meyer, C.G. Bluespine Unicornfish (Naso unicornis) Are Both Natural Control Agents and Mobile Vectors for Invasive Algae in a Hawaiian Marine Reserve. Mar. Biol. 2017, 164, 25. [Google Scholar] [CrossRef]
- Mumby, P.J.; Edwards, A.J.; Arias-Gonazalez, E.J.; Lindeman, K.C.; Blackwell, P.G.; Gall, A.; Gorczynska, M.I.; Harborne, A.R.; Pescod, C.L.; Renken, H.; et al. Mangroves Enhance the Biomass of Coral Reef Fish Communities in the Caribbean. Nature 2004, 427, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Serafy, J.E.; Shideler, G.S.; Araújo, R.J.; Nagelkerken, I. Mangroves Enhance Reef Fish Abundance at the Caribbean Regional Scale. PLoS ONE 2015, 10, e0142022. [Google Scholar] [CrossRef] [PubMed]
- Lugendo, B.R.; Nagelkerken, I.; van der Velde, G.; Mgaya, Y.D. The Importance of Mangroves, Mud and Sand Flats, and Seagrass Beds as Feeding Areas for Juvenile Fishes in Chwaka Bay, Zanzibar: Gut Content and Stable Isotope Analyses. J. Fish Biol. 2006, 69, 1639–1661. [Google Scholar] [CrossRef]
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.; Choat, J.H. Integrating Ecological Roles and Trophic Diversification on Coral Reefs: Multiple Lines of Evidence Identify Parrotfishes as Microphages. Biol. J. Linn. Soc. 2017, 120, 729–751. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving Resource Partitioning in Parrotfishes (Scarini) Using Microhistology of Feeding Substrata. Coral Reefs 2020, 39, 1313–1327. [Google Scholar] [CrossRef]
- Russ, G.R.; Questel, S.L.A.; Rizzari, J.R.; Alcala, A.C. The Parrotfish–Coral Relationship: Refuting the Ubiquity of a Prevailing Paradigm. Mar. Biol. 2015, 162, 2029–2045. [Google Scholar] [CrossRef]
- Carlson, P.M.; Davis, K.; Warner, R.R.; Caselle, J.E. Fine-Scale Spatial Patterns of Parrotfish Herbivory Are Shaped by Resource Availability. Mar. Ecol. Prog. Ser. 2017, 577, 165–176. [Google Scholar] [CrossRef]
- Eggertsen, L.; Goodell, W.; Cordeiro, C.A.M.M.; Mendes, T.C.; Longo, G.O.; Ferreira, C.E.L.; Berkström, C. Seascape Configuration Leads to Spatially Uneven Delivery of Parrotfish Herbivory across a Western Indian Ocean Seascape. Diversity 2020, 12, 434. [Google Scholar] [CrossRef]
- Fulton, C.J.; Bellwood, D.R. Patterns of Foraging in Labrid Fishes. Mar. Ecol. Prog. Ser. 2002, 226, 135–142. [Google Scholar] [CrossRef]
- Wainwright, P.C.; Bellwood, D.R.; Westneat, M.W. Ecomorphology of Locomotion in Labrid Fishes. Environ. Biol. Fishes 2002, 65, 47–62. [Google Scholar] [CrossRef]
- Russ, G.R.; Lowe, J.R.; Rizzari, J.R.; Bergseth, B.J.; Alcala, A.C. Partitioning No-Take Marine Reserve (NTMR) and Benthic Habitat Effects on Density of Small and Large-Bodied Tropical Wrasses. PLoS ONE 2017, 12, e0188515. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.R.; Williamson, D.H.; Ceccarelli, D.M.; Evans, R.D.; Russ, G.R. Responses of Coral Reef Wrasse Assemblages to Disturbance and Marine Reserve Protection on the Great Barrier Reef. Mar. Biol. 2019, 166, 119. [Google Scholar] [CrossRef]
- Green, A.L. Spatial, Temporal and Ontogenetic Patterns of Habitat Use by Coral Reef Fishes (Family Labridae). Mar. Ecol. Prog. Ser. 1996, 133, 1–11. [Google Scholar] [CrossRef]
- Berkström, C.; Jones, G.P.; McCormick, M.I. Trade-Offs in the Ecological Versatility of Juvenile Wrasses: An Experimental Evaluation. J. Exp. Mar. Biol. Ecol. 2014, 453, 91–97. [Google Scholar] [CrossRef]
- Berkström, C.; Jones, G.P.; McCormick, M.I.; Srinivasan, M. Ecological Versatility and Its Importance for the Distribution and Abundance of Coral Reef Wrasses. Mar. Ecol. Prog. Ser. 2012, 461, 151–163. [Google Scholar] [CrossRef]
- Kramer, M.J.; Bellwood, O.; Bellwood, D.R. Foraging and Microhabitat Use by Crustacean-Feeding Wrasses on Coral Reefs. Mar. Ecol. Prog. Ser. 2016, 548, 277–282. [Google Scholar] [CrossRef]
- Shibuno, T.; Nakamura, Y.; Horinouchi, M.; Sano, M. Habitat Use Patterns of Fishes across the Mangrove-Seagrass-Coral Reef Seascape at Ishigaki Island, Southern Japan. Ichthyol. Res. 2008, 55, 218–237. [Google Scholar] [CrossRef]
- Gullström, M.; Berkström, C.; Öhman, M.C.; Bodin, M.; Dahlberg, M. Scale-Dependent Patterns of Variability of a Grazing Parrotfish (Leptoscarus vaigiensis) in a Tropical Seagrass-Dominated Seascape. Mar. Biol. 2011, 158, 1483–1495. [Google Scholar] [CrossRef]
- Tano, S.A.; Eggertsen, M.; Wikstrom, S.; Berkstrom, C.; Buriyo, A.; Halling, C. Tropical Seaweed Beds as Important Habitats for Juvenile Fish. Mar. Freshw. Res. 2017, 68, 1921–1934. [Google Scholar] [CrossRef]
- Wilson, S.K.; Depczynski, M.; Holmes, T.H.; Noble, M.M.; Radford, B.T.; Tinkler, P.; Fulton, C.J. Climatic Conditions and Nursery Habitat Quality Provide Indicators of Reef Fish Recruitment Strength. Limnol. Oceanogr. 2017, 62, 1868–1880. [Google Scholar] [CrossRef]
- Wilson, S.K.; Depczynski, M.; Fisher, R.; Holmes, T.H.; O’Leary, R.A.; Tinkler, P. Habitat Associations of Juvenile Fish at Ningaloo Reef, Western Australia: The Importance of Coral and Algae. PLoS ONE 2010, 5, e15185. [Google Scholar] [CrossRef] [PubMed]
- Olds, A.D.; Connolly, R.M.; Pitt, K.A.; Maxwell, P.S.; Aswani, S.; Albert, S. Incorporating Surrogate Species and Seascape Connectivity to Improve Marine Conservation Outcomes. Conserv. Biol. 2014, 28, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Grober-Dunsmore, R.; Frazer, T.K.; Beets, J.P.; Lindberg, W.J.; Zwick, P.; Funicelli, N.A. Influence of Landscape Structure on Reef Fish Assemblages. Landsc. Ecol. 2008, 23, 37–53. [Google Scholar] [CrossRef]
- Moustaka, M.; Evans, R.D.; Kendrick, G.A.; Hyndes, G.A.; Cuttler, M.V.W.; Bassett, T.J.; O’Leary, M.J.; Wilson, S.K. Local Habitat Composition and Complexity Outweigh Seascape Effects on Fish Distributions across a Tropical Seascape. Landsc. Ecol. 2024, 39, 28. [Google Scholar] [CrossRef]
- Alcala, A.C.; Russ, G.R. A Direct Test of the Effects of Protective Management on a Tropical Marine Reserve. ICES J. Mar. Sci. 1990, 46, 40–47. [Google Scholar] [CrossRef]
- Russ, G.R.; Alcala, A.C. Marine Reserves: Long-Term Protection Is Required for Full Recovery of Predatory Fish Populations. Oecologia 2004, 138, 622–627. [Google Scholar] [CrossRef]
- Russ, G.R.; Rizzari, J.R.; Abesamis, R.A.; Alcala, A.C. Coral Cover a Stronger Driver of Reef Fish Trophic Biomass than Fishing. Ecol. Appl. 2021, 31, e02224. [Google Scholar] [CrossRef] [PubMed]
- Verweij, M.C.; Nagelkerken, I. Short and Long-Term Movement and Site Fidelity of Juvenile Haemulidae in Back-Reef Habitats of a Caribbean Embayment. Hydrobiologia 2007, 592, 257–270. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Bothwell, J.; Nemeth, R.S.; Pitt, J.M.; van der Velde, G. Interlinkage between Caribbean Coral Reefs and Seagrass Beds through Feeding Migrations by Grunts (Haemulidae) Depends on Habitat Accessibility. Mar. Ecol. Prog. Ser. 2008, 368, 155–164. [Google Scholar] [CrossRef]
- Sekund, L.; Pittman, S. Explaining Island-wide Geographical Patterns of Caribbean Fish Diversity: A Multi-scale Seascape Ecology Approach. Mar. Ecol. 2017, 38, e12434. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. Are Ecologists Conducting Research at the Optimal Scale? Glob. Ecol. Biogeogr. 2015, 24, 52–63. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Name | Coral Reef Level Habitat Variables | Seascape-Level Habitat Variables | Location (see Figure 1) | Number of Transects |
|---|---|---|---|---|
| A.SG/Soft coral | Soft coral cover | Area of seagrass | 1 | 28 |
| A.MG/A.Sand | None | Area of mangrove; area of sand | 2 | 23 |
| Coral/A.MA | Hard coral cover | Area of macroalgae | 3 | 32 |
| A.CR/A.RF | None | Area of coral reef; area of reef flat | 4 | 12 |
| Rubble/EAM | Rubble cover, EAM cover | None | 5 | 13 |
| Habitat Cluster | All Species | Parrotfish | Wrasse |
|---|---|---|---|
| Area Seagrass | Chaetodon baronessa | Scarus tricolor | Choerodon spp. |
| Soft Coral | Scarus ghobban | Cheilio inermis | |
| Scarus rivulatus | Epibulus brevis | ||
| Cheilinus trilobatus | |||
| Area Mangrove | Ctenochaetus striatus | Chlorurus spp. | Anampses sp. |
| Area Sand | Acanthurus nigrofuscus | Scarus chameleon | Anampses meleagrides |
| Plectorhinchus sp. | Scarus hypselopterus | Cheilinus chlorurous | |
| Coral Cover | Naso lituratus | Scarus spp. | Bodianus mesothorax, |
| Area Macroalgae | Zebrasoma scopas | Scarus dimidiatus | Labrichthys unilineatus |
| Naso unicornis | Scarus niger | Cheilinus oxychephalus | |
| Chlorurus bleekeri | |||
| Area Coral Reef | Pterocaesio tile | Chlorurus microrhinos | Bodianus dictynna |
| Area Reef Flat | Caesio caerulaurea | Scarus psittacus | Coris gaimard |
| Caesio teres | Gomphosus varius | ||
| Naso vlamingii | |||
| Rubble | Ctenochaetus binotatus | Chlorurus bowersi | Coris batuensis |
| EAM | Naso minor | Chlorurus bleekeri | Hemigymnus melapterus |
| Novaculichthys taeniourus | |||
| Oxycheilinus celebicus | |||
| Cheilinus fasciatus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sievers, K.T.; McClure, E.C.; Abesamis, R.A.; Russ, G.R. Multi-Scale Coral Reef and Seascape Habitat Variables Combine to Influence Reef Fish Assemblages. Fishes 2024, 9, 137. https://doi.org/10.3390/fishes9040137
Sievers KT, McClure EC, Abesamis RA, Russ GR. Multi-Scale Coral Reef and Seascape Habitat Variables Combine to Influence Reef Fish Assemblages. Fishes. 2024; 9(4):137. https://doi.org/10.3390/fishes9040137
Chicago/Turabian StyleSievers, Katie T., Eva C. McClure, Rene A. Abesamis, and Garry R. Russ. 2024. "Multi-Scale Coral Reef and Seascape Habitat Variables Combine to Influence Reef Fish Assemblages" Fishes 9, no. 4: 137. https://doi.org/10.3390/fishes9040137
APA StyleSievers, K. T., McClure, E. C., Abesamis, R. A., & Russ, G. R. (2024). Multi-Scale Coral Reef and Seascape Habitat Variables Combine to Influence Reef Fish Assemblages. Fishes, 9(4), 137. https://doi.org/10.3390/fishes9040137