1. Introduction
Fish meal (FM) is widely recognized as the primary protein source in aquatic feeds, particularly for carnivorous fish, owing to its elevated digestible protein, well-balanced amino acids, and minor antinutritional factors [
1,
2]. In recent decades, the rapid growth of aquaculture has escalated the demand for aquatic feeds, exacerbating the imbalance between FM supply and demand [
2]. However, the excessive exploitation of marine fish resources has resulted in a significant reduction in fishing area, consequently leading to an ongoing decrease in FM production [
3]. Therefore, it is imperative to urgently explore alternative protein sources as substitutes for FM to reduce FM inclusion and dietary costs.
Replacing FM with plant protein sources, such as rapeseed meal [
4], peanut meal [
5], soybean protein meal [
6], etc., represents a prominent research direction in the field of fish nutrition. However, antinutritional factors such as gossypol, trypsin inhibitors, phytic acids, and tannin in traditional plant protein sources are not conducive to the digestion, absorption, and growth of fish [
7]. By improving the plant protein source through processing technology and fermentation technology, the antinutritional factors can be effectively removed, and the availability of plant protein source can be promoted [
8]. Cottonseed protein concentrate (CPC) is a product derived from common cottonseed meal through a low-temperature leaching and double-solvent extraction process, exhibiting notable characteristics such as reduced antinutritional factor content and elevated protein content [
9,
10]. CPC is considered a superior plant protein source and an ideal alternative to FM [
10]. Some studies demonstrated that the substitution of a portion of FM in the diet with CPC had no detrimental impact on the growth performance of largemouth bass (
Micropterus salmoides) [
11], rainbow trout (
Oncorhynchus mykiss) [
2], golden pompano (
Trachinotus ovatus) [
12], and pearl gentiana grouper (
Epinephelus fuscoguttatus ♀ ×
Epinephelus lanceolatu ♂) [
13]. Furthermore, the incorporation of CPC as a partial substitute for FM resulted in enhanced hepatic antioxidant capacity in rainbow trout [
2] and golden pompano [
12]. These findings indicate that it is possible to use CPC as a substitute for a portion of FM in the diets for carnivorous fish.
The hybrid F
1, resulting from a cross between
Erythroculter ilishaeformis (♀) and
Ancherythroculter nigrocauda (♂), exhibits remarkable traits including rapid growth, high stress resistance, delicious meat, cost-effective feeding, and robust disease resistance [
14,
15]. Currently, the cultivation of the hybrid F
1 is widespread across China. Previous studies have demonstrated that the hybrid culter exhibits a carnivorous feeding behavior, with its dietary composition typically comprising 40% FM [
16]. However, there are currently no reports documenting the replacement of FM with CPC in the diet of the hybrid culter. Therefore, to clarify the effects of the dietary CPC on growth and health, the growth performance, serum biochemical parameters, muscle amino acid composition, antioxidant properties, and intestinal morphology of hybrid culter were examined. The findings provide a theoretical foundation for the implementation of CPC in a low FM diet for carnivorous fish.
2. Materials and Methods
2.1. Experimental Diets
The formulation and proximate composition of the experimental diets are shown in
Table 1, and the amino acid compositions of diets are illustrated in
Table 2. Experimental basic diets were compounded with FM, soybean meal, and chicken meal as main protein sources (about 42% protein); fish oil and soybean oil as main lipid sources (about 11.4% lipid); and wheat flour as a carbohydrate source. Four experimental diets were prepared by replacing the same amount of FM with about 0, 6%, 12%, and 18% CPC in the basal diet, recorded as T0, T15, T30, and T45, respectively. All the large particles of ingredients were crushed first and then passed through a 60-mesh sieve. Dry ingredients listed in
Table 1 were thoroughly mixed in a mixer before the addition of oil and 40% water. The mixture was extruded through a single-screw extruder to form Φ1 mm strips, subsequently desiccated at 60 °C for 2 h, and finally crushed into pellet feed with a length of approximately 5 mm using a crusher, and then were stored at −20 °C until used.
2.2. Feeding and Management
The experimental fish were provided by the Fisheries Research Institute, Wuhan Academy of Agricultural Sciences. Fish were domesticated in a recirculating aquacultural system for 2 weeks. After being acclimatized for 2 weeks, A total of 360 fish (mean initial weight: 10.0 ± 0.5 g) with uniform size and health were randomly distributed to 12 tanks with 30 fish per tank. Each experimental diet was randomly assigned to triplicate tanks. Fish were hand-fed twice daily (08:00 and 16:30) at a daily feeding rate of 3% of body weight for 70 days. Uneaten feed and feces were removed and approximately 10% of the water in each tank was replenished. During the experiment, the water temperature was maintained at 26.5–27.5 °C, the dissolved oxygen was kept at more than 5 mg/L, and regular monitoring of ammonia nitrogen and nitrite concentrations was performed. The dissolved oxygen and temperature were measured using a YSI-Pro20 dissolved oxygen meter, while ammonia nitrogen and nitrite were analyzed using a HACH ultraviolet–visible spectrophotometer (DR6000) and the corresponding detection kits.
2.3. Sample Collection
After a 70-day feeding trial, all fish were fasting for 24 h and then were weighed and counted. Three fish were randomly selected from each tank, anesthetized with an overdose of MS-222, euthanized, and stored at −40 °C for determination of whole fish nutrient composition. Additionally, six randomly chosen fish from each tank were anesthetized (75 mg/L MS-222) to measure their body weight and length. Subsequently, blood samples from the tail vein of these fish were collected to separate serum for the analysis of biochemical indicators. Then, the dorsal muscles were collected and subsequently stored at −80 °C to facilitate the subsequent analysis of muscle amino acid composition. The visceral mass, liver, and foregut of six fish from each tank were extracted, while the visceral mass and liver underwent weighing to calculate hepatosomatic and viscerosomatic index, respectively. Then, foregut tissue along with part of the liver tissue was fixed with paraformaldehyde fixative for preparation of liver and intestinal sections; another portion of the liver tissue was stored at −40 °C for assessment of antioxidant indexes.
2.4. Chemical Composition and Enzyme Activity Analysis Methods
Proximate compositions of experimental diets and whole fish samples were measured according to the Association of Official Agricultural Chemists methods [
17]. Crude protein content was determined by measuring nitrogen (N × 6.25) levels using the Kjeldahl method following acid digestion with an auto Kjeldahl System (Kjelflex K-360; BUCHI Labortechnik AG, Flawil, Switzerland), and the crude lipid content was determined by Soxhlet extraction. Moisture content was measured by freeze-drying samples for 48 h in a vacuum freeze dryer (Christ Beta 2–4 LD plus LT, Marin Christ Corporation, Osterode, Germany). Ash level was examined through incineration at 550 °C for 24 h in a muffle furnace. The amino acid composition of diets and muscle samples were assessed using an automatic amino acid analyzer (Hitachi L-8900, Tokyo, Japan) after pretreatment with hydrochloric acid hydrolysis.
Serum glucose (GLU), serum albumin (ALB), triglyceride (TG), total protein (TP), and total cholesterol (TCHO), along with alkaline phosphatase (ALP), aminotransferase (AST), and alanine aminotransferase (ALT) were determined using an automatic biochemical analyzer (CHEMIX-800, Sysmex Corporation, Kobe, Japan) with commercial diagnostic reagent kits (Sysmex Wuxi Co., Ltd., Wuxi, China).
The activities of superoxide dismutase (SOD) and catalase (CAT) and the content of malondialdehyde (MDA) were detected with commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.5. Histological Analysis
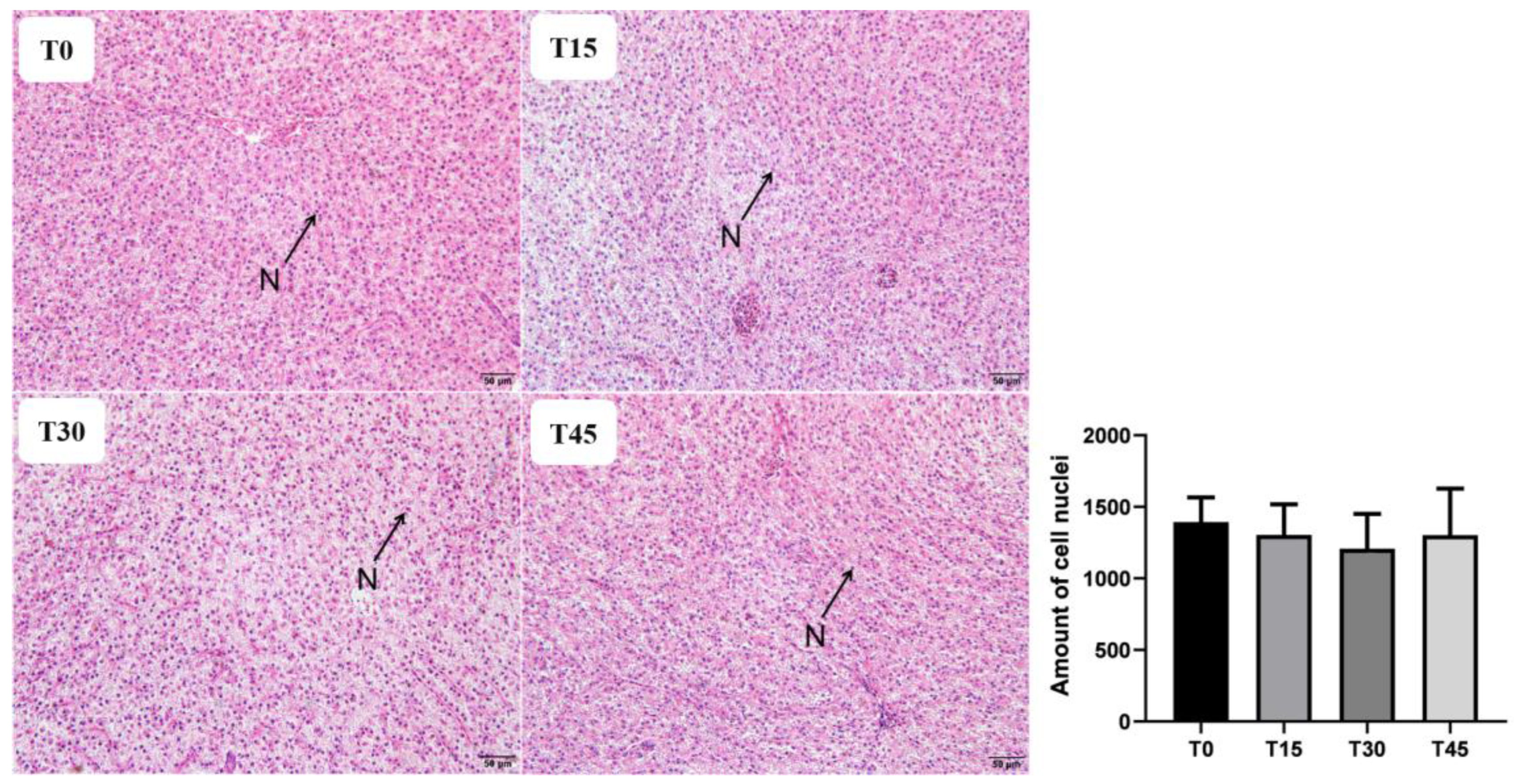
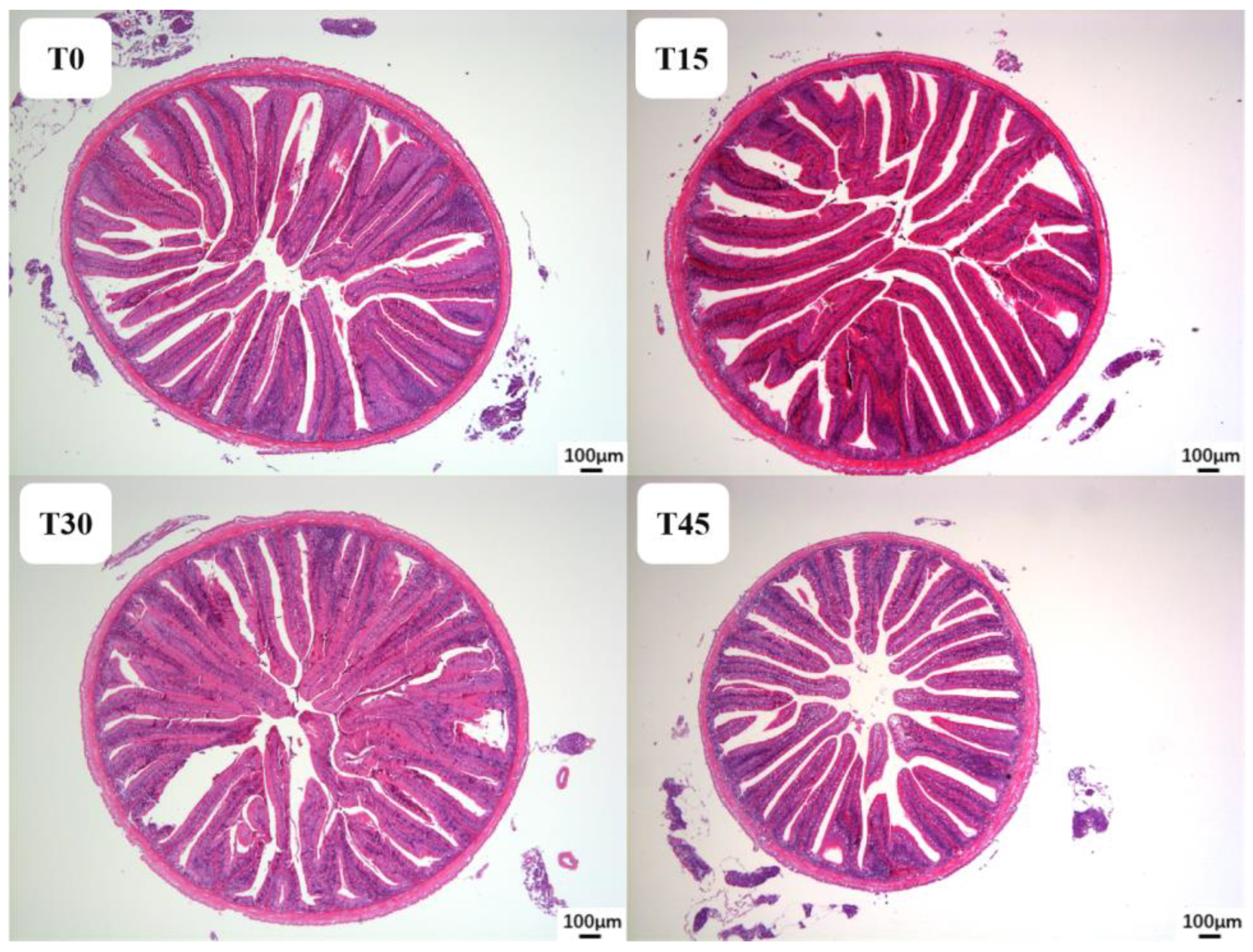
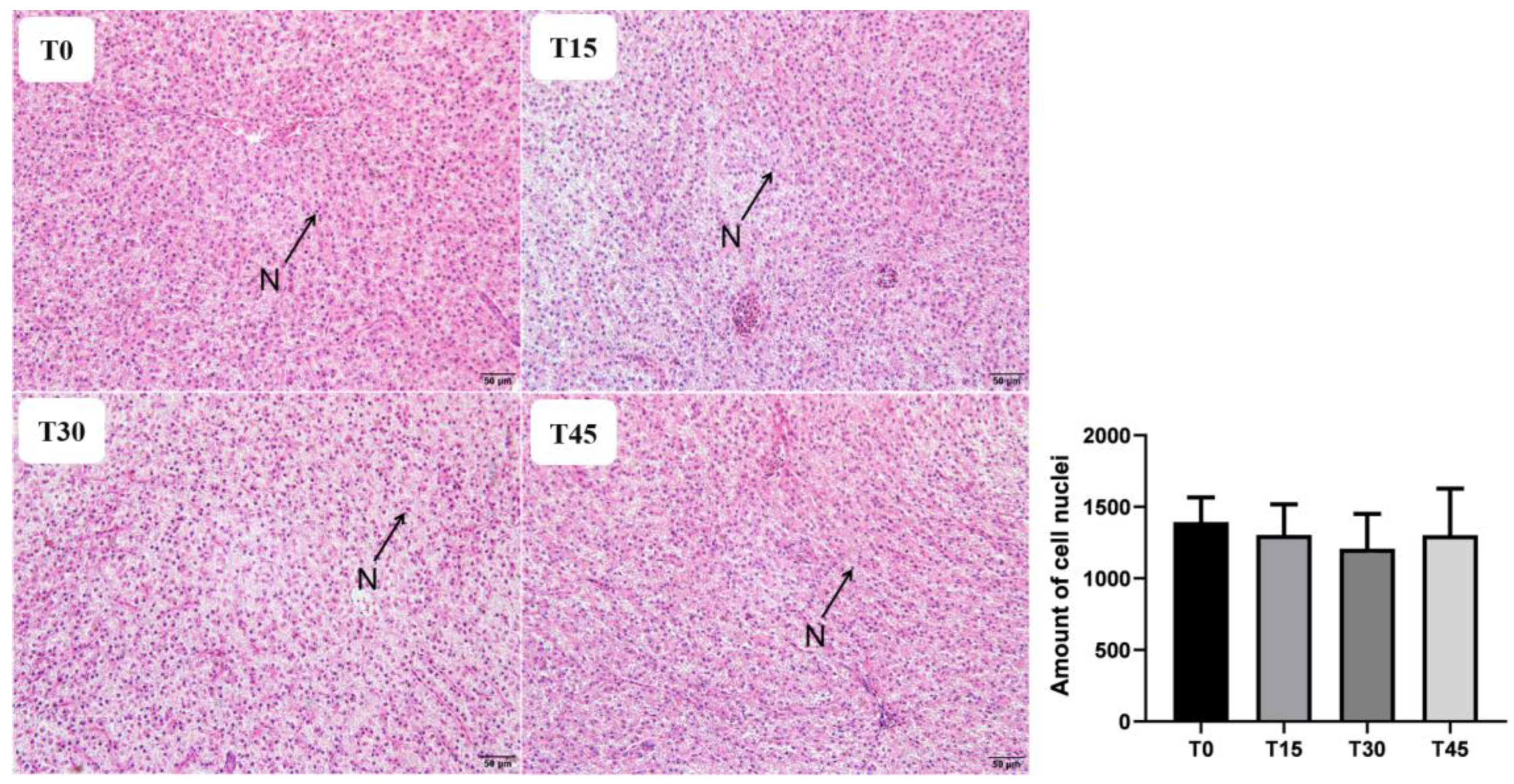
Liver and foregut tissues were dehydrated by ethanol, then permeabilized, soaked in wax and embedded in paraffin, cut into 5 μm slices, dried, and then stained with hematoxylin–eosin (H&E). Three slides were obtained from liver and intestinal sections of each group for quantification, with three randomly selected fields analyzed on each slide. The number of cell nuclei in liver and the villi height, width, and muscular layer thickness of foregut were measured using Image J (National Institutes of Health, Bethesda, MD, USA).
2.6. Statistical Analysis
All data were analyzed using the statistical software SPSS 20.0 (SPSS Inc., Chicago, IL, USA). Data were subjected to a homogeneity test and a normality test prior to analysis by the one-way analysis of variance (ANOVA) and Duncan’s multiple range test. The results were described as mean ± standard deviation (SD), where the threshold of statistical significance was p < 0.05.
4. Discussion
Previous studies revealed that partial replacement of FM with a plant protein source has the benefit of decreasing diet cost and without adverse effects on growth, but higher replacement levels caused its growth retardation [
18,
19]. In largemouth bass, the replacement of 30% FM by CPC had no significant impacts on growth, flesh composition, flavor, or muscular tissue structure, whereas high FM replacement (≥45%) decreased growth performance [
19]. Similar results were also reported in common carp (
Cyprinus carpio) [
20], hybrid grouper (♀
Epinephelus fuscoguttatus × ♂
Epinephelus lanceolatu) [
21], and cobia (
Rachycentron canadum) [
22]. Our study demonstrated a consistent decrease in WG with increasing dietary CPC level and decreasing dietary FM, and the WG in T30 and T45 groups was significantly lower than that in the control group. Although the CPC had undergone dephenolization, it still contained various antinutrient factors such as gossyphene, phytic acid, and tannin. These compounds form complexes with proteins, sugars, and various metal ions that are not easily digestible or absorbable by the body, thus adversely affecting the palatability, feed intake, and growth [
21,
23]. In this study, the hybrid culter group fed with T15 did not impact on growth performance and SR (≥90%). Higher dietary CPC (T30 and T45) significantly decreased the growth performance and food intake of hybrid culter. Several studies have shown that replacing FM with higher levels of CPC reduces the SR of largemouth bass [
10,
24] and rainbow trout [
2,
25]. Furthermore, there were no significant differences in FCR among all the groups, which is consistent with the results on rainbow trout [
25] and golden pompano [
26].
Studies have demonstrated that the utilization of phenol-free cottonseed meal and CPC as FM substitutes does not significantly impact the body composition of rainbow trout [
24], black sea bass (
Centropristis striata) [
27], and Chinese soft-shelled turtle (
Pelodiscus sinensis) [
28]. However, other studies have shown that replacing FM with CPC has a significant effect on the body composition [
10,
26]. The current study indicates an increasing trend in crude lipid content of whole fish as the CPC replacement proportion increases. Notably, the T45 group exhibits a significantly higher concentration of crude lipid compared to the T0 group, which aligns with previous results observed in largemouth bass [
1,
11] and southern flounder (
Paralichthys lethostigma) [
29]. Nevertheless, no significant differences were observed in terms of crude protein and ash contents between groups. These observations may be attributed to variations in cottonseed protein processing methods, levels of FM substitution, and experimental conditions [
24,
30].
Blood is the pivotal component of the circulatory system in the body and plays crucial roles in material transportation, physiological regulation, and defense mechanisms. In fish, serum biochemical indicators effectively reflect various metabolic processes, nutritional status, and health [
31]. Previous studies have demonstrated that gossypol consumption may result in liver damage and associated alterations in serum biochemistry [
32]. AST and ALT are two vital transaminase enzymes involved in amino acid metabolism, with increased activities typically indicating liver damage [
33]. Our findings revealed an elevation in serum ALT activity with increasing CPC content in the diet, where the T45 group exhibited the highest value compared to the other groups. The results indicated that a higher substitution of FM with CPC influences the hepatic function in hybrid culter. Similar observations have been reported in hybrid grouper [
21], Amur sturgeon (
Acipenser schrenckii) [
34], and rainbow trout [
35]. Serum TP and ALB levels serve as indicators for protein metabolic status and immune capacity in fish [
36]. Our study indicated a decreasing trend in serum TP and ALB contents with increasing proportions of CPC replacement. This could be attributed to nutritional deficiencies or abnormalities in hepatic protein metabolism [
37,
38]. Serum T-CHO and TG levels provide insights into lipid metabolism within the body [
39]. Our results demonstrated that a decreasing trend was observed in the T-CHO and TG contents in serum, and at the same time, the whole-fish lipid level increased significantly with the dietary CPC level. This observation implies that the TG and TCHO in serum may accelerate the transformation into lipids and deposition in fish, but the results of serum biochemical indicators (TG and TCHO) are not significant. Further investigation is warranted to elucidate the mechanism by which CPC affects lipid deposition in fish.
The nutritional composition and content of animal muscle are intricately linked to its quality. The nutritional value of muscle proteins is determined by the composition, types, and proportions of amino acids, which serve as the fundamental building blocks [
19]. The flavor quality of fish fillets mainly depends on alanine, glutamic acid, aspartic acid, and glycine, which contribute to the umami and sweetness of fish fillets [
40,
41]. In this study, 17 essential and nonessential amino acids in the diet and muscles were detected in the muscle of hybrid culter. No differences were found in the amino acid profiles under the replacement of FM by CPC. This indicates that CPC can replace up to 45% of FM in the diet without negatively impacting the composition of muscle amino acids. Our findings are consistent with previous research on Nile tilapia (
Oreochromis niloticus) [
42] and large yellow croaker (
Larimichthys crocea) [
43], which reported that increasing CPC in the diet did not significantly affect EAA or TAA contents in muscles. However, studies on largemouth bass [
10] and Amur sturgeon [
34] have shown that replacing FM with a high proportion of CPC significantly reduced muscular TAA and EAA contents. This may be explained by higher dietary CPC content causing an increase in gossypol level or an imbalance in amino acid compositions.
The antioxidant defense system plays a crucial role in protecting cells and cell membranes against oxidative damage, thus maintaining normal physiological functions of the body [
44]. Antioxidant enzymes are important indicators and serve as markers for animal health and their response to external stimuli [
12]. Excessive production of reactive oxygen species (ROS) disrupts cellular redox balance, causing metabolic disorders and impairments in genetic material. SOD and CAT are widely recognized as antioxidant enzymes in fish responsible for scavenging excessive ROS, indicating stronger antioxidant properties within the fish’s body [
45,
46]. MDA is the product resulting from the reaction between oxygen free radicals and lipid oxidation. Its content reflects the degree of lipid peroxidation and damage to cell membranes [
47]. It has been reported that gossypol readily binds to proteins in the electron transport chain of mitochondria, thereby disrupting mitochondrial function and inducing excessive production of ROS, consequently inhibiting various enzyme activities and initiating oxidative damage [
48]. Studies have demonstrated that high dietary CPC replacement levels result in decreased liver antioxidant capacity (SOD, CAT, GSH-PX) or reduced expression of antioxidant-related genes in largemouth bass [
11] and Ussuri catfish (
Pseudobagrus ussuriensis) [
44]. Another study revealed that high substitution ratios of FM with a mixture of Clostridium ethanolic protein and CPC (1:1) could diminish the antioxidant capacity in rainbow trout [
49]. The findings from this study suggest that an increase in CPC substitution content within FM tends to decrease T-SOD and CAT activities. Notably, the T45 group exhibits significantly lower T-SOD and CAT activities compared to the T0 group, but there was no significant difference in MDA content. These findings suggest that elevated dietary CPC diminish the antioxidant capacity of juvenile hybrid culter, but the increased dietary CPC did not cause more serious oxidative damage to the liver. Indeed, this conclusion was further confirmed by the results of liver morphology and the number of hepatocyte cell nuclei. However, contrasting findings have been reported in previous studies: SOD activity was found to be enhanced in golden pompano [
12], while no significant effect on MDA content was observed for the replacement of FM with CPC. These discrepancies may be attributed to variations in feed formulations or stress resistance of fish species.
The intestine functions as the primary organ responsible for digestion and absorption of nutrients in fish, playing a crucial role in normal growth and development. The dimensions of villus height and width within the intestine determine the surface area of contact between mucosal epithelial cells and chyme, while the thickness of the muscle layer facilitates intestinal peristalsis and advancement of chyle [
49]. Studies have shown that high CPC levels (CPC60) in diets can lead to intestinal morphological changes (including villus height and number of goblet cells, etc.) [
11]. The reduction in intestinal villus height may contribute to inhibition of growth performance. Furthermore, higher replacement level of CPC has been found to significantly decrease both the height and width of intestinal villi in rainbow trout [
35], large yellow croaker [
43], and largemouth bass [
1]. The inclusion of a mixture consisting of clostridium ethanol protein and CPC (1:1) as a substitute for FM resulted in significant reductions in villus height, villus width, and muscle layer thickness within the intestines of rainbow trout [
49]. Similarly, substituting FM with low gossypol cottonseed meal can induce morphological damage to the intestines of silver sillago (
Sillago sihama) [
9]. The findings from this study revealed that the T45 group, which had a high proportion of CPC replacement, exhibited significantly decreased intestinal villi height compared to the control group, consistent with previous research [
1,
35,
43,
49]. These results demonstrate a notable decline in intestinal villi height of hybrid culter when replacing FM with a high proportion of CPC.



 ,
, 

{kind=link}
{kind=link}
{kind=link}