
Estimating Cognitive Ability in the Wild: Validation of a Detour Test Paradigm Using a Cichlid Fish (Neolamprologus pulcher)


Abstract
1. Introduction
2. Methods
2.1. Study Species
2.2. Field Work
2.3. Video Analyses (Structural Complexity)
2.4. Experimental Setup
2.5. Video Analyses (Behavioural)
2.6. Statistical Analysis
3. Results
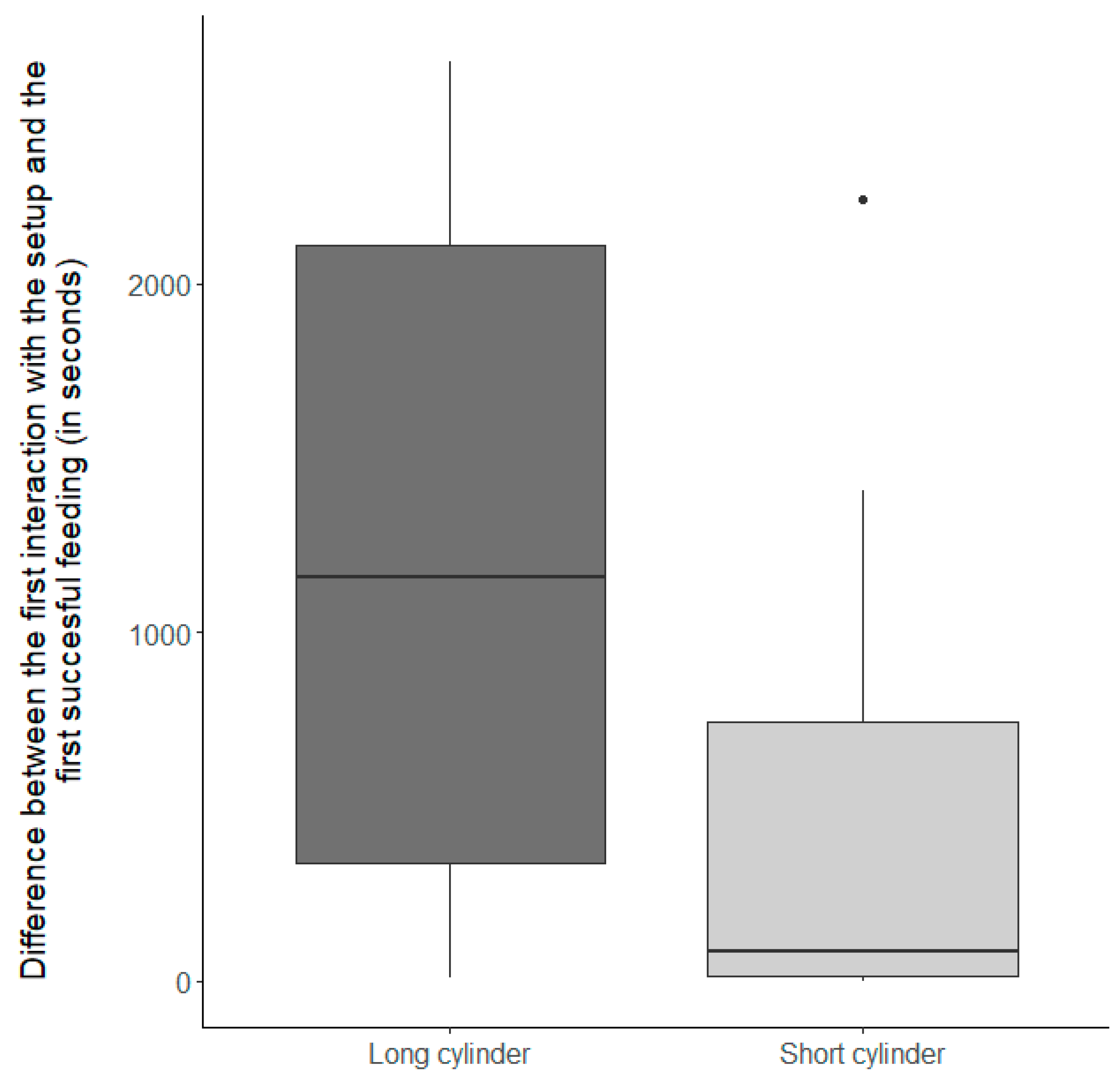
3.1. Does the Setup Pose a Cognitive Challenge to the Fish?
3.1.1. Control Condition
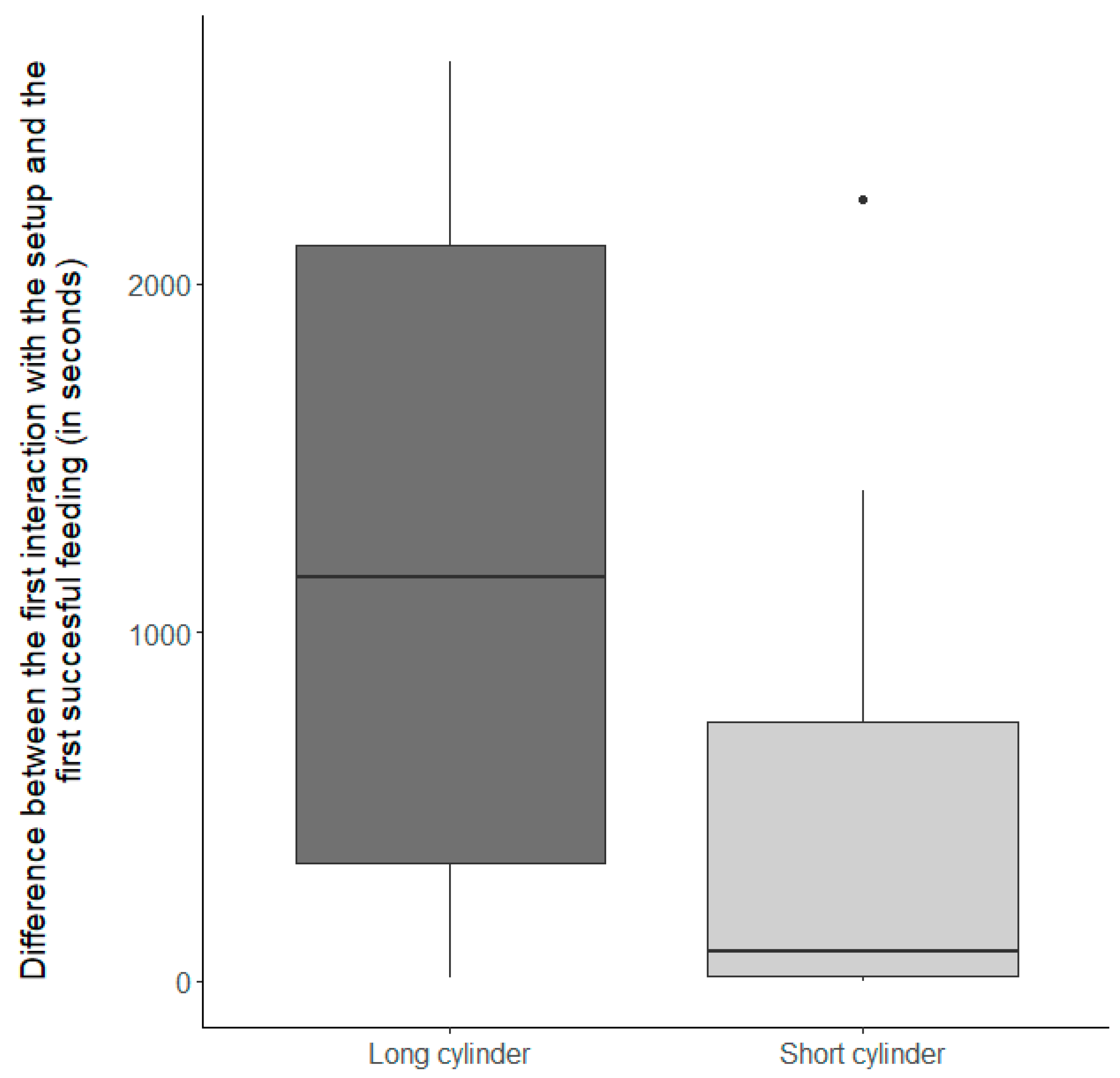
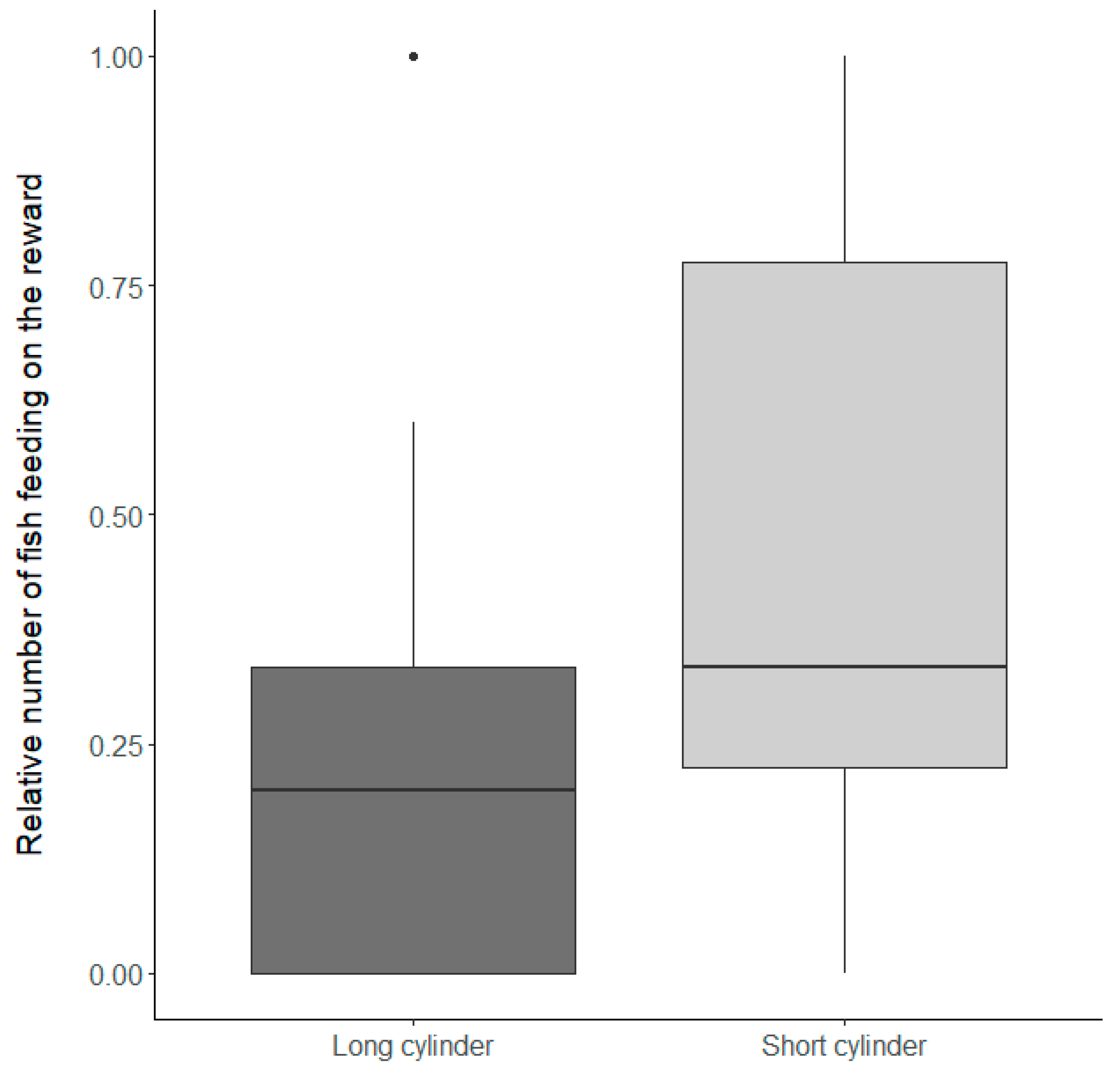
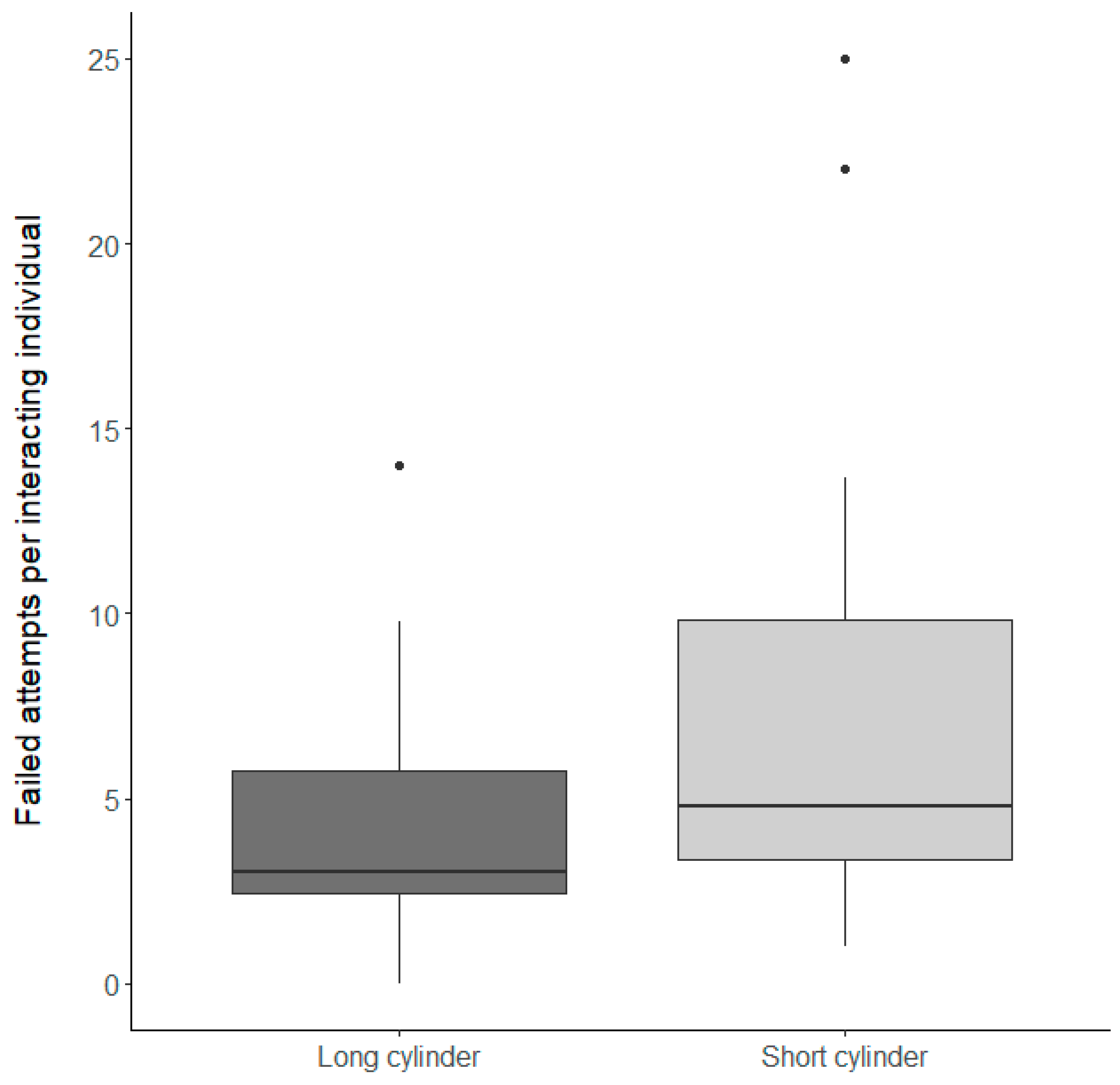
3.1.2. Inhibitory Control Conditions
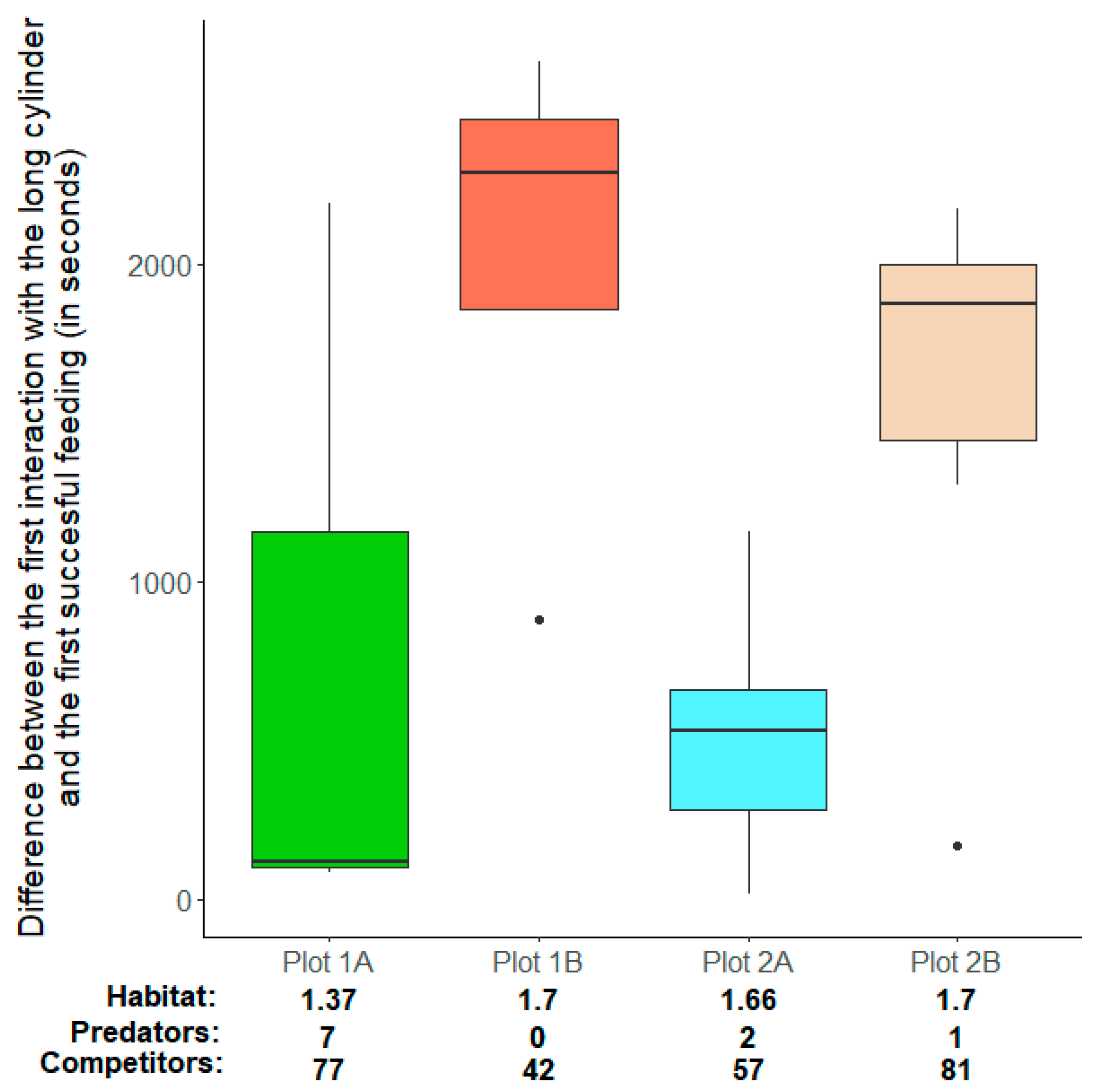
3.2. Do Habitat Complexity and/or Group Size Influence the Motivation to Find Food and the Ability to Solve a Cognitive Challenge?
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaufman, A.B.; Call, J.; Kaufman, J.C. (Eds.) The Cambridge Handbook of Animal Cognition, 1st ed.; Cambridge University Press: Cambridge, UK, 2021; ISBN 978-1-108-42674-9. [Google Scholar]
- Maclean, E.L. Unraveling the Evolution of Uniquely Human Cognition. Proc. Natl. Acad. Sci. USA 2016, 113, 6348–6354. [Google Scholar] [CrossRef]
- Pravosudov, V.V.; Roth, T.C., II. Cognitive Ecology of Food Hoarding: The Evolution of Spatial Memory and the Hippocampus. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 173–193. [Google Scholar] [CrossRef]
- Berghänel, A.; Lazzaroni, M.; Cimarelli, G.; Marshall-Pescini, S.; Range, F. Cooperation and Cognition in Wild Canids. Curr. Opin. Behav. Sci. 2022, 46, 101173. [Google Scholar] [CrossRef]
- Healy, S.D. The Face of Animal Cognition. Integr. Zool. 2019, 14, 132–144. [Google Scholar] [CrossRef] [PubMed]
- van Horik, J.; Emery, N.J. Evolution of Cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2011, 2, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Mettke-Hofmann, C. Cognitive Ecology: Ecological Factors, Life-Styles, and Cognition. WIREs Cogn. Sci. 2014, 5, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Henke-von der Malsburg, J.; Kappeler, P.M.; Fichtel, C. Linking Ecology and Cognition: Does Ecological Specialisation Predict Cognitive Test Performance? Behav. Ecol. Sociobiol. 2020, 74, 154. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S. Evolution in the Social Brain. Science 2007, 317, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Sewall, K.B. Social Complexity as a Driver of Communication and Cognition. Integr. Comp. Biol. 2015, 55, 384–395. [Google Scholar] [CrossRef]
- Shumway, C.A. Habitat Complexity, Brain, and Behavior. Brain. Behav. Evol. 2008, 72, 123–134. [Google Scholar] [CrossRef]
- DeCasien, A.R.; Williams, S.A.; Higham, J.P. Primate Brain Size Is Predicted by Diet but Not Sociality. Nat. Ecol. Evol. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Sandi, C. Stress and Cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2013, 4, 245–261. [Google Scholar] [CrossRef]
- Griffin, A.S.; Guillette, L.M.; Healy, S.D. Cognition and Personality: An Analysis of an Emerging Field. Trends Ecol. Evol. 2015, 30, 207–214. [Google Scholar] [CrossRef]
- Fischer, S.; Balshine, S.; Hadolt, M.C.; Schaedelin, F.C. Siblings Matter: Family Heterogeneity Improves Associative Learning Later in Life. Ethology 2021, 127, 897–907. [Google Scholar] [CrossRef]
- Alves, J.; de Sá Couto-Pereira, N.; Merscher Sobreira de Lima, R.; Quillfeldt, J.A.; Dalmaz, C. Effects of Early Life Adversities upon Memory Processes and Cognition in Rodent Models. Neuroscience 2022, 497, 282–307. [Google Scholar] [CrossRef] [PubMed]
- Heldstab, S.A.; Isler, K.; Graber, S.M.; Schuppli, C.; van Schaik, C.P. The Economics of Brain Size Evolution in Vertebrates. Curr. Biol. 2022, 32, R697–R708. [Google Scholar] [CrossRef] [PubMed]
- Reichert, M.S.; Quinn, J.L. Cognition in Contests: Mechanisms, Ecology, and Evolution. Trends Ecol. Evol. 2017, 32, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Dukas, R. Evolutionary Biology of Animal Cognition. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 347–374. [Google Scholar] [CrossRef]
- Beran, M.J.; Parrish, A.E.; Perdue, B.M.; Washburn, D.A. Comparative Cognition: Past, Present, and Future. Int. J. Comp. Psychol. 2014, 27, 1. [Google Scholar] [CrossRef]
- Bräuer, J.; Hanus, D.; Pika, S.; Gray, R.; Uomini, N. Old and New Approaches to Animal Cognition: There Is Not “One Cognition”. J. Intell. 2020, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- McGrew, W.C. Is Primate Tool Use Special? Chimpanzee and New Caledonian Crow Compared. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120422. [Google Scholar] [CrossRef]
- Krasheninnikova, A.; Chow, P.K.Y.; von Bayern, A.M.P. Comparative Cognition: Practical Shortcomings and Some Potential Ways Forward. Can. J. Exp. Psychol. Can. Psychol. Expérimentale 2020, 74, 160–169. [Google Scholar] [CrossRef]
- Mikhalevich, I.; Powell, R.; Logan, C. Is Behavioural Flexibility Evidence of Cognitive Complexity? How Evolution Can Inform Comparative Cognition. Interface Focus 2017, 7, 20160121. [Google Scholar] [CrossRef]
- Kabadayi, C.; Bobrowicz, K.; Osvath, M. The Detour Paradigm in Animal Cognition. Anim. Cogn. 2018, 21, 21–35. [Google Scholar] [CrossRef]
- MacLean, E.L.; Hare, B.; Nun, C.L.; Addess, E.; Amic, F.; Anderson, R.C.; Aureli, F.; Baker, J.M.; Bania, A.E.; Barnard, A.M.; et al. The Evolution of Self-Control. Proc. Natl. Acad. Sci. USA 2014, 111, E2140–E2148. [Google Scholar] [CrossRef]
- Santacà, M.; Busatta, M.; Lucon-Xiccato, T.; Bisazza, A. Sensory Differences Mediate Species Variation in Detour Task Performance. Anim. Behav. 2019, 155, 153–162. [Google Scholar] [CrossRef]
- Shaw, R.C.; Schmelz, M. Cognitive Test Batteries in Animal Cognition Research: Evaluating the Past, Present and Future of Comparative Psychometrics. Anim. Cogn. 2017, 20, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Salena, M.G.; Turko, A.J.; Singh, A.; Pathak, A.; Hughes, E.; Brown, C.; Balshine, S. Understanding Fish Cognition: A Review and Appraisal of Current Practices. Anim. Cogn. 2021, 24, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Bshary, R.; Triki, Z. Fish Ecology and Cognition: Insights from Studies on Wild and Wild-Caught Teleost Fishes. Curr. Opin. Behav. Sci. 2022, 46, 101174. [Google Scholar] [CrossRef]
- Vila Pouca, C.; Brown, C. Contemporary Topics in Fish Cognition and Behaviour. Curr. Opin. Behav. Sci. 2017, 16, 46–52. [Google Scholar] [CrossRef]
- Healy, S.D.; Patton, B.W. It Began in Ponds and Rivers: Charting the Beginnings of the Ecology of Fish Cognition. Front. Vet. Sci. 2022, 9, 823143. [Google Scholar] [CrossRef] [PubMed]
- Schnell, A.K.; Amodio, P.; Boeckle, M.; Clayton, N.S. How Intelligent Is a Cephalopod? Lessons from Comparative Cognition. Biol. Rev. 2021, 96, 162–178. [Google Scholar] [CrossRef]
- Braga Goncalves, I.; Ashton, B.J.; Fischer, S. Causes and Consequences of Cognitive Variation in Fishes. Fishes 2023, 8, 277. [Google Scholar] [CrossRef]
- Herrmann, E.; Call, J.; Hernández-Lloreda, M.V.; Hare, B.; Tomasello, M. Humans Have Evolved Specialized Skills of Social Cognition: The Cultural Intelligence Hypothesis. Science 2007, 317, 1360–1366. [Google Scholar] [CrossRef]
- Gingins, S.; Bshary, R. The Cleaner Wrasse Outperforms Other Labrids in Ecologically Relevant Contexts, but Not in Spatial Discrimination. Anim. Behav. 2016, 115, 145–155. [Google Scholar] [CrossRef]
- Salwiczek, L.H.; Prétôt, L.; Demarta, L.; Proctor, D.; Essler, J.; Pinto, A.I.; Wismer, S.; Stoinski, T.; Brosnan, S.F.; Bshary, R. Adult Cleaner Wrasse Outperform Capuchin Monkeys, Chimpanzees and Orang-Utans in a Complex Foraging Task Derived from Cleaner—Client Reef Fish Cooperation. PLoS ONE 2012, 7, e49068. [Google Scholar] [CrossRef]
- Triki, Z.; Emery, Y.; Teles, M.C.; Oliveira, R.F.; Bshary, R. Brain Morphology Predicts Social Intelligence in Wild Cleaner Fish. Nat. Commun. 2020, 11, 6423. [Google Scholar] [CrossRef]
- Ronco, F.; Matschiner, M.; Böhne, A.; Boila, A.; Büscher, H.H.; Indermaur, A.; El Taher, A.; Malinsky, M.; Ricci, V.; Kahmen, A.; et al. Drivers and Dynamics of a Massive Adaptive Radiation in African Cichlid Fish. Nature 2021, 589, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Taborsky, M.; Wong, M.Y.L. Sociality in Fishes. In Comparative Social Evolution; Rubenstein, D.R., Abbot, P., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 354–389. ISBN 9781107338319. [Google Scholar]
- Fischer, S.; Bohn, L.; Oberhummer, E.; Nyman, C.; Taborsky, B. Divergence of Developmental Trajectories Is Triggered Interactively by Early Social and Ecological Experience in a Cooperative Breeder. Proc. Natl. Acad. Sci. USA 2017, 114, E9300–E9307. [Google Scholar] [CrossRef]
- Fischer, S.; Bessert-Nettelbeck, M.; Kotrschal, A.; Taborsky, B. Rearing-Group Size Determines Social Competence and Brain Structure in a Cooperatively Breeding Cichlid. Am. Nat. 2015, 186, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Reddon, A.R.; O’Connor, C.M.; Ligocki, I.Y.; Hellmann, J.K.; Marsh-rollo, S.E.; Hamilton, I.M.; Balshine, S. No Evidence for Larger Brains in Cooperatively Breeding Cichlid Fishes. Can. J. Zool. 2016, 94, 373–378. [Google Scholar] [CrossRef]
- Salena, M.G.; Singh, A.; Weller, O.; Fang, X.X.; Balshine, S. Rapid Spatial Learning in Cooperative and Non-Cooperative Cichlids. Behav. Process. 2022, 194, 104550. [Google Scholar] [CrossRef]
- Wallace, K.J.; Choudhary, K.D.; Kutty, L.A.; Le, D.H.; Lee, M.T.; Wu, K.; Hofmann, H.A. Social Ascent Changes Cognition, Behaviour and Physiology in a Highly Social Cichlid Fish. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20200448. [Google Scholar] [CrossRef] [PubMed]
- Lindenfors, P.; Wartel, A.; Lind, J. “Dunbar’s Number” Deconstructed. Biol. Lett. 2021, 17, 20210158. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.E.; Isler, K.; Barton, R.A. Re-Evaluating the Link between Brain Size and Behavioural Ecology in Primates. Proc. R. Soc. B Biol. Sci. 2017, 284, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.; Dunbar, R.I.M. Both Social and Ecological Factors Predict Ungulate Brain Size. Proc. R. Soc. B Biol. Sci. 2006, 273, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Bergman, T.J.; Beehner, J.C. Measuring Social Complexity. Anim. Behav. 2015, 103, 203–209. [Google Scholar] [CrossRef]
- Kappeler, P.M. A Framework for Studying Social Complexity. Behav. Ecol. Sociobiol. 2019, 73, 13. [Google Scholar] [CrossRef]
- Byrne, R.W.; Bates, L.A. Sociality, Evolution and Cognition. Curr. Biol. 2007, 17, 714–723. [Google Scholar] [CrossRef]
- Chapman, B.B.; Ward, A.J.W.; Krause, J. Schooling and Learning: Early Social Environment Predicts Social Learning Ability in the Guppy, Poecilia reticulata. Anim. Behav. 2008, 76, 923–929. [Google Scholar] [CrossRef]
- Jordan, L.A.; Taborsky, B.; Taborsky, M. Cichlids as a Model System for Studying Social Behaviour and Evolution. In The Behaviour, Ecology, and Evolution of Cichlid Fishes: A Contemporary Modern Synthesis; Abate, M.E., Noakes, D.L.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 587–635. [Google Scholar]
- Shumway, C.A. The Evolution of Complex Brains and Behaviors in African Cichlid Fishes. Curr. Zool. 2010, 56, 144–156. [Google Scholar] [CrossRef]
- Kotrschal, A.; Taborsky, B. Environmental Change Enhances Cognitive Abilities in Fish. PLoS Biol. 2010, 8, e1000351. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Laland, K.N. Social Learning in Fishes: A Review. Fish Fish. 2003, 4, 280–288. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental Enrichment for Fish in Captive Environments: Effects of Physical Structures and Substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Taborsky, M. Cichlid Fishes: A Model for the Integrative Study of Social Behavior. In Cooperative Breeding in Vertebrates: Studies of Ecology, Evolution and Behavior; Koenig, W.D., Dickinson, J.L., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 272–293. [Google Scholar]
- Heg, D.; Bachar, Z.; Brouwer, L.; Taborsky, M. Predation Risk Is an Ecological Constraint for Helper Dispersal in a Cooperatively Breeding Cichlid. Proc. R. Soc. B Biol. Sci. 2004, 271, 2367–2374. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Zöttl, M.; Bonfils, D.; Josi, D.; Frommen, J.G.; Taborsky, M. Philopatry Yields Higher Fitness than Dispersal in a Cooperative Breeder with Sex-Specific Life History Trajectories. Sci. Adv. 2023, 9, eadd2146. [Google Scholar] [CrossRef] [PubMed]
- Dey, C.J.; Reddon, A.R.; O’Connor, C.M.; Balshine, S. Network Structure Is Related to Social Conflict in a Cooperatively Breeding Fish. Anim. Behav. 2013, 85, 395–402. [Google Scholar] [CrossRef]
- Hellmann, J.K.; Ligocki, I.Y.; O’Connor, C.M.; Reddon, A.R.; Garvy, K.A.; Marsh-Rollo, S.E.; Gibbs, H.L.; Balshine, S.; Hamilton, I.M. Reproductive Sharing in Relation to Group and Colony-Level Attributes in a Cooperative Breeding Fish. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150954. [Google Scholar] [CrossRef]
- Jungwirth, A.; Taborsky, M. First- and Second-Order Sociality Determine Survival and Reproduction in Cooperative Cichlids. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151971. [Google Scholar] [CrossRef]
- Zöttl, M.; Frommen, J.G.; Taborsky, M. Group Size Adjustment to Ecological Demand in a Cooperative Breeder. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122772. [Google Scholar] [CrossRef]
- Fischer, S.; Zöttl, M.; Groenewoud, F.; Taborsky, B. Group-Size-Dependent Punishment of Idle Subordinates in a Cooperative Breeder Where Helpers Pay to Stay. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140184. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Walker, J.; Taborsky, M. Prospecting Precedes Dispersal and Increases Survival Chances in Cooperatively Breeding Cichlids. Anim. Behav. 2015, 106, 107–114. [Google Scholar] [CrossRef]
- Groenewoud, F.; Frommen, J.G.; Josi, D.; Tanaka, H.; Jungwirth, A.; Taborsky, M. Predation Risk Drives Social Complexity in Cooperative Breeders. Proc. Natl. Acad. Sci. USA 2016, 113, 4104–4109. [Google Scholar] [CrossRef] [PubMed]
- Lein, E.; Jordan, A. Studying the Evolution of Social Behaviour in One of Darwin’s Dreamponds: A Case for the Lamprologine Shell-Dwelling Cichlids. Hydrobiologia 2021, 848, 3699–3726. [Google Scholar] [CrossRef]
- Duftner, N.; Sefc, K.M.; Koblmüller, S.; Salzburger, W.; Taborsky, M.; Sturmbauer, C. Parallel Evolution of Facial Stripe Patterns in the Neolamprologus brichardi/pulcher Species Complex Endemic to Lake Tanganyika. Mol. Phylogenet. Evol. 2007, 45, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Nührenberg, P.; Jordan, A. On the Importance of Defendable Resources for Social Evolution: Applying New Techniques to a Long-Standing Question. Ethology 2021, 127, 872–885. [Google Scholar] [CrossRef]
- Hartley, R.; Zisserman, A. Multiple View Geometry in Computer Vision, 2nd ed.; Cambridge University Press: Cambridge, UK, 2004; ISBN 978-0-521-54051-3. [Google Scholar]
- Griwodz, C.; Gasparini, S.; Calvet, L.; Gurdjos, P.; Castan, F.; Maujean, B.; De Lillo, G.; Lanthony, Y. AliceVision Meshroom: An Open-Source 3D Reconstruction Pipeline. In Proceedings of the MMSys ’21: 12th ACM Multimedia Systems Conference, Istanbul, Turkey, 28 September–1 October 2021; pp. 241–247. [Google Scholar]
- Blender Online Community. Blender—A 3D Modelling and Rendering Package; Blender Foundation, Stichting Blender Foundation: Amsterdam, The Netherlands, 2018. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.M.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Singmann, H.; Bolker, B.; Westfall, J.; Aust, F.; Ben-Sachar, M.S.; Højsgaard, S.; Fox, J.; Lawrence, M.A.; Mertens, U.; Love, J.; et al. Afex: Analysis of Factorial Experiments. 2023. Available online: https://CRAN.R-project.org/package=afex (accessed on 20 November 2023).
- Dunbar, R.I.M. The Social Brain Hypothesis and Human Evolution. Oxford Res. Encycl. Psychol. 2016. [Google Scholar] [CrossRef]
- Jungwirth, A.; Balzarini, V.; Zöttl, M.; Salzmann, A.; Taborsky, M.; Frommen, J.G. Long-Term Individual Marking of Small Freshwater Fish: The Utility of Visual Implant Elastomer Tags. Behav. Ecol. Sociobiol. 2019, 73, 49. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Bisazza, A. Sex Differences in Spatial Abilities and Cognitive Flexibility in the Guppy. Anim. Behav. 2017, 123, 53–60. [Google Scholar] [CrossRef]
- Fischer, S.; Jungwirth, A. The Costs and Benefits of Larger Brains in Fishes. J. Evol. Biol. 2022, 35, 973–985. [Google Scholar] [CrossRef]
- Brown, V.J.; Tait, D.S. Behavioral Flexibility: Attentional Shifting, Rule Switching, and Response Reversal. In Encyclopedia of Psychopharmacology; Stoleman, I.P., Price, L.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Tebbich, S.; Stereln, K.; Teschke, I. The Tale of the Finch: Adaptive Radiation and Behavioural Flexibility. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A. Executive Functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed]
- Amici, F.; Aureli, F.; Call, J. Fission-Fusion Dynamics, Behavioral Flexibility, and Inhibitory Control in Primates. Curr. Biol. 2008, 18, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.R.; Ericson, B.N.; Hauser, M.D. Constraints on Problem Solving and Inhibition: Object Retrieval in Cotton-Top Tamarins (Saguinus oedipus oedipus). J. Comp. Psychol. 1999, 113, 186–193. [Google Scholar] [CrossRef]
- Brucks, D.; Marshall-Pescini, S.; Range, F. Dogs and Wolves Do Not Differ in Their Inhibitory Control Abilities in a Non-Social Test Battery. Anim. Cogn. 2019, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Ulrich, L.; Holekamp, K.E. Group Size and Social Rank Predict Inhibitory Control in Spotted Hyaenas. Anim. Behav. 2020, 160, 157–168. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Montalbano, G.; Reddon, A.R.; Bertolucci, C. Social Environment Affects Inhibitory Control via Developmental Plasticity in a Fish. Anim. Behav. 2022, 183, 69–76. [Google Scholar] [CrossRef]
- Logan, C.J.; McCune, K.B.; MacPherson, M.; Johnson-Ulrich, Z.; Rowney, C.; Seitz, B.; Blaisdell, A.P.; Deffner, D.; Wascher, C.A.F. Are the More Flexible Individuals Also Better at Inhibition? Anim. Behav. Cogn. 2022, 9, 14–36. [Google Scholar] [CrossRef]
- Izquierdo, A.; Brigman, J.L.; Radke, A.K.; Rudebeck, P.H.; Holmes, A. The Neural Basis of Reversal Learning: An Updated Perspective. Neuroscience 2017, 345, 12–26. [Google Scholar] [CrossRef]
- Schluessel, V.; Leo, V.; Bawolt, M.; Kreuter, N. When the Penny Drops: Sharks Outsmart Cichlids in Serial Reversal Learning. Behaviour 2023, 160, 1259–1281. [Google Scholar] [CrossRef]
- Boogert, N.J.; Fawcett, T.W.; Lefebvre, L. Mate Choice for Cognitive Traits: A Review of the Evidence in Nonhuman Vertebrates. Behav. Ecol. 2011, 22, 447–459. [Google Scholar] [CrossRef]
- Ashton, B.; Ridley, A.R.; Edwards, E.K.; Thornton, A. Cognitive Performance Is Linked to Group Size and Affects Fitness in Australian Magpies. Nature 2018, 554, 364–367. [Google Scholar] [CrossRef]
- Bebus, S.E.; Small, T.W.; Jones, B.C.; Elderbrock, E.K.; Schoech, S.J. Associative Learning Is Inversely Related to Reversal Learning and Varies with Nestling Corticosterone Exposure. Anim. Behav. 2016, 111, 251–260. [Google Scholar] [CrossRef]
- Boogert, N.J.; Monceau, K.; Lefebvre, L. A Field Test of Behavioural Flexibility in Zenaida Doves (Zenaida aurita). Behav. Process. 2010, 85, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Cauchoix, M.; Hermer, E.; Chaine, A.S.; Morand-Ferron, J. Cognition in the Field: Comparison of Reversal Learning Performance in Captive and Wild Passerines. Sci. Rep. 2017, 7, 12945. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, M.; Stark, M.; Dufour, V.; Jordan, A. Cognitive Flexibility in a Tanganyikan Bower-Building Cichlid, Aulonocranus dewindti. Anim. Cogn. 2023, 26, 1959–1971. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Estimate ± SE | Num. D.F. | Den. D.F. | F-Value | p-Value |
|---|---|---|---|---|---|
| (a) Latency to interact | |||||
| Intercept | 5.28 ± 0.24 | - | - | - | - |
| Inhibitory control condition | 0.01 ± 0.21 | 1 | 18 | 1 × 10−3 | 0.97 |
| (b) Latency to feed on the reward after the first interaction with the setup | |||||
| Intercept | 6.54 ± 0.40 | - | - | - | |
| Inhibitory control condition | −1.84 ± 0.48 | 1 | 18 | 14.91 | <0.01 |
| (c) Failed attempts | |||||
| Intercept | 1.45 ± 0.16 | ||||
| Inhibitory control condition | 0.44 ± 0.2 | 1 | 18 | 4.9 | 0.04 |
| Factors | Estimate ± SE | Χ2-Value | p-Value |
|---|---|---|---|
| Intercept | 1.67 ± 0.23 | - | - |
| Inhibitory control condition | −0.92 ± 0.31 | 8.81 | <0.01 |
| Factors | Estimate ± SE | Num. D.F. | Den. D.F. | F-Value | p-Value |
|---|---|---|---|---|---|
| (a) Latency to feed on the reward in the control condition | |||||
| Intercept | 7.56 ± 0.42 | - | - | - | - |
| Location | - | 3 | 19 | 0.01 | 0.99 |
| Plot 1B | 0.05 ± 0.46 | - | - | - | - |
| Plot 2A | −5 × 10−3 ± 0.41 | - | - | - | - |
| Plot 2B | 0.04 ± 0.38 | - | - | - | - |
| Group size | −0.05 ± 0.02 | 1 | 19 | 3.33 | 0.08 |
| (b) Latency to feed on the reward after the first interaction with the long cylinder | |||||
| Intercept | −219.59 ± 662.68 | - | - | - | |
| Location | - | 3 | 14 | 5.88 | <0.01 |
| Plot 1B | 1977.75 ± 655.71 | - | - | - | - |
| Plot 2A | 170.60 ± 539.15 | - | - | - | - |
| Plot 2B | 982.17 ± 498.20 | - | - | - | - |
| Group size | 62.20 | 1 | 14 | 3.67 | 0.07 |
| (c) Latency to feed on the reward after the first interaction with the short cylinder | |||||
| Intercept | 170.52 ± 711.77 | - | - | - | - |
| Location | - | 3 | 13 | 0.28 | 0.84 |
| Plot 1B | 446.08 ± 701.58 | - | - | - | - |
| Plot 2A | −8.21 ± 581.73 | - | - | - | - |
| Plot 2B | 198.78 ± 530.78 | - | - | - | - |
| Group size | 14.68 ± 35.06 | 1 | 13 | 0.18 | 0.68 |
| Contrast | Estimate ± SE | t-Value | p-Value |
|---|---|---|---|
| Intercept | 563.03 ± 384.21 | 1.47 | 0.16 |
| (a) Comparison among plots in locality 1 versus plots in locality 2 | |||
| (Plot 1A, Plot 1B) vs. (Plot 2A, Plot 2B) | −206.25 ± 165.11 | −1.25 | 0.23 |
| (b) Comparison among plots within locality 1 | |||
| Plot 1A vs. Plot 1B | 988.87 ± 327.86 | 3.02 | 0.01 |
| (c) Comparison among plots within locality 2 | |||
| Plot 2A vs. Plot 2B | 405.78 ± 208.06 | 1.95 | 0.07 |
| Factors | Estimate ± SE | Χ2-Value | p-Value |
|---|---|---|---|
| (a) Number of fish feeding relative to the number of fish interacting (long cylinder) | |||
| Intercept | 2.27 ± 0.82 | - | - |
| Location | - | 5.35 | 0.15 |
| Plot 1B | 0.74 ± 1.22 | ||
| Plot 2A | −0.94 ± 0.59 | ||
| Plot 2B | −0.58 ± 0.68 | ||
| Group size | −0.01 ± 0.05 | 0.09 | 0.76 |
| (b) Number of fish feeding relative to the number of fish interacting (short cylinder) | |||
| Intercept | 1.49 ± 0.91 | - | - |
| Location | - | 1.33 | 0.72 |
| Plot 1B | −0.07 ± 0.93 | - | - |
| Plot 2A | −0.52 ± 0.66 | - | - |
| Plot 2B | −0.57 ± 0.65 | - | - |
| Group size | −0.04 ± 0.05 | 0.58 | 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jungwirth, A.; Horsfield, A.; Nührenberg, P.; Fischer, S. Estimating Cognitive Ability in the Wild: Validation of a Detour Test Paradigm Using a Cichlid Fish (Neolamprologus pulcher). Fishes 2024, 9, 50. https://doi.org/10.3390/fishes9020050
Jungwirth A, Horsfield A, Nührenberg P, Fischer S. Estimating Cognitive Ability in the Wild: Validation of a Detour Test Paradigm Using a Cichlid Fish (Neolamprologus pulcher). Fishes. 2024; 9(2):50. https://doi.org/10.3390/fishes9020050
Chicago/Turabian StyleJungwirth, Arne, Anna Horsfield, Paul Nührenberg, and Stefan Fischer. 2024. "Estimating Cognitive Ability in the Wild: Validation of a Detour Test Paradigm Using a Cichlid Fish (Neolamprologus pulcher)" Fishes 9, no. 2: 50. https://doi.org/10.3390/fishes9020050
APA StyleJungwirth, A., Horsfield, A., Nührenberg, P., & Fischer, S. (2024). Estimating Cognitive Ability in the Wild: Validation of a Detour Test Paradigm Using a Cichlid Fish (Neolamprologus pulcher). Fishes, 9(2), 50. https://doi.org/10.3390/fishes9020050