
Effect of a Commercial Polyphenol Compound on the Performance and Antioxidant Status of Penaeus vannamei


 , , , ,
, , , , 
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Diets
2.2. Experimental Design
2.3. Growth Parameters
2.4. Shrimp Hepatopancreas Collection and Processing
2.5. Biochemical Analysis
2.6. Statistical Analyses
3. Results
3.1. Growth Performance
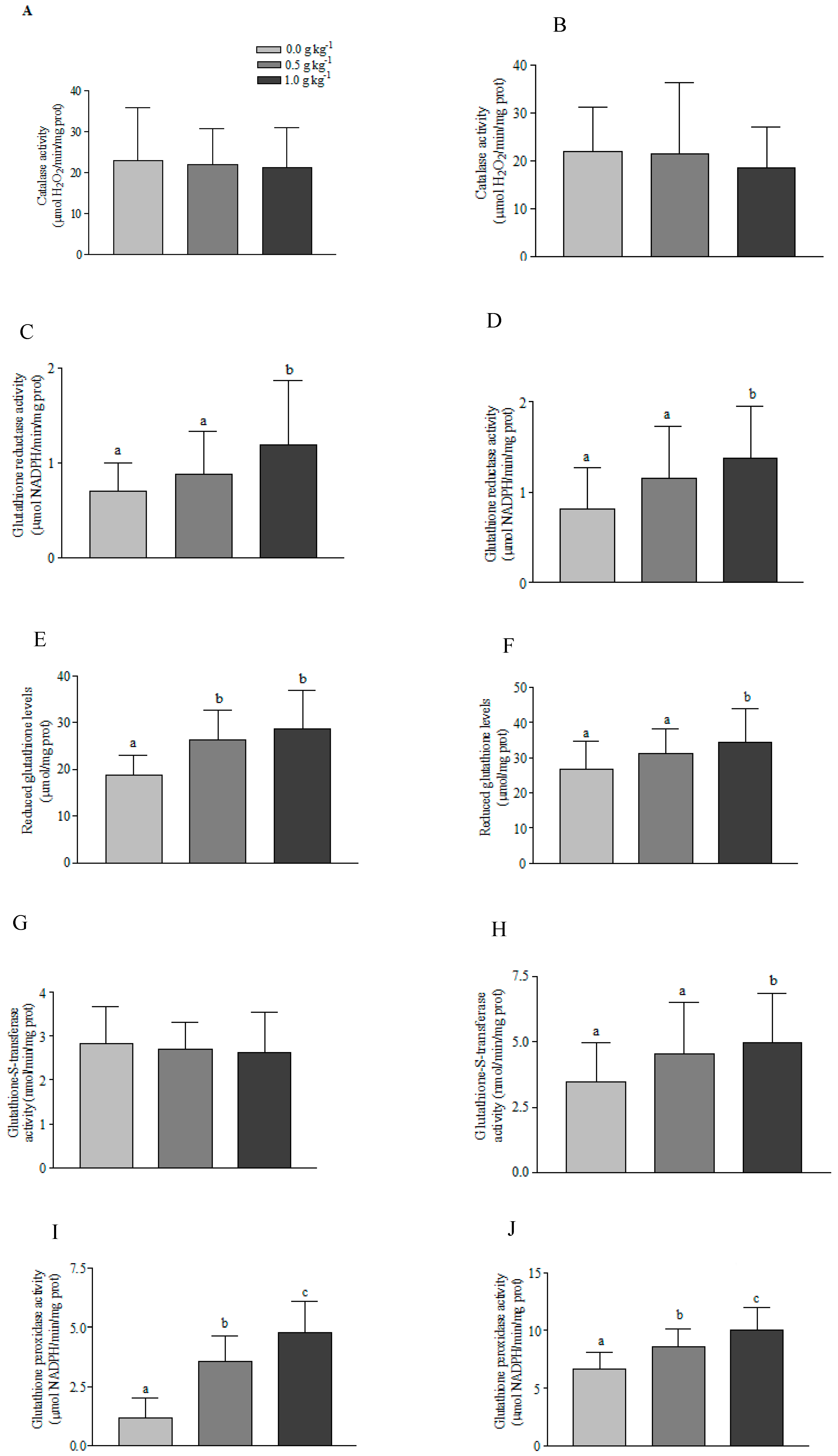
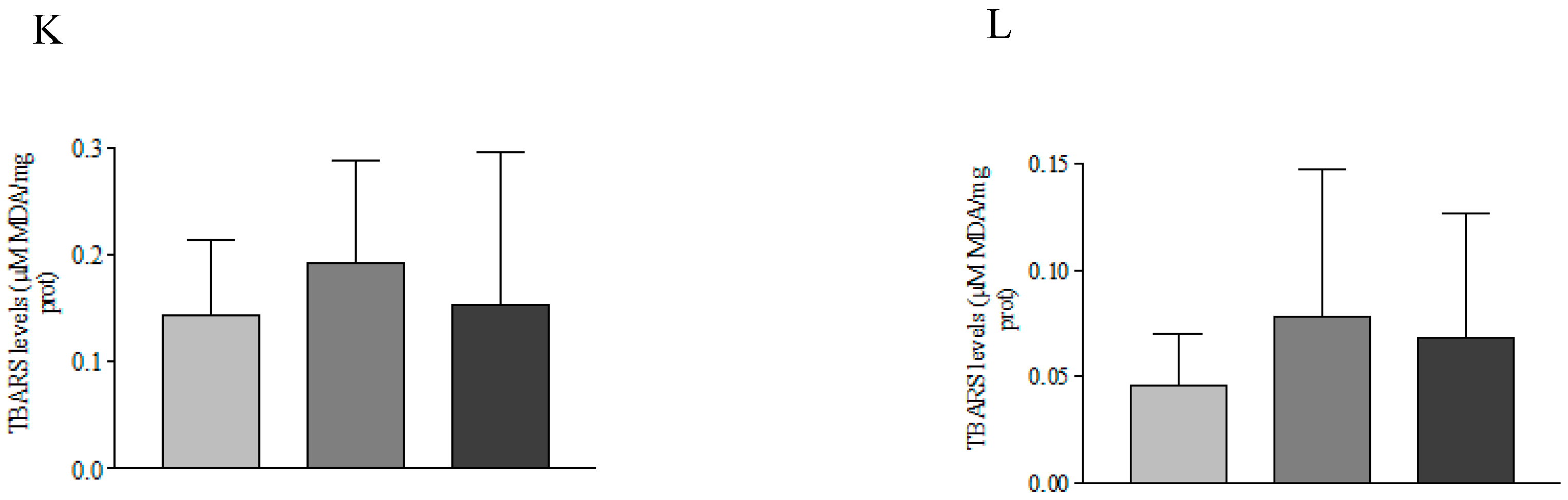
3.2. Antioxidant Defense System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mălina Petrescu-Mag, R.; Păsărin, B.; Gigi Şonea, C.; Petrscu-Mag, I.V. Customer preferences and trends for aquarium fish in Transylvania (Romania). North-West. J. Zool. 2013, 9, 166. [Google Scholar]
- FAO. The state of world fisheries and aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Ravichndran, P.; Panigrahi, A.; Kumaran, M. Biology and culture of Litopenaeus vannamei vis-Ã-vis Penaeus monodon. In Handbook on Seed Production and Farming of Litopenaeus vannamei; Central Institute of Brackishwater Aquaculture (CIBA), Indian Council of Agricultural Research (ICAR): New Delhi, India, 2009; Volume 46, p. 70. [Google Scholar]
- Cavalheiro, T.B.; Carvajal, J.C.L.; de Lucena, R.F.P.; do Nascimento, C.V.C.; Ribeiro, T.T.B.C. Water parameters correlated with the zootechnical performance of shrimp Litopenaeus vannamei grown in oligohaline waters. Acta Scientiarum. Anim. Sci. 2022, 45, e57700. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Liang, Z.; Liu, R.; Zhao, D.; Wang, L.; Sun, M.; Wang, M.; Song, L. Ammonia exposure induces oxidative stress, endoplasmic reticulum stress and apoptosis in hepatopancreas of pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2016, 54, 523–528. [Google Scholar] [CrossRef]
- Banh, S.; Wiens, L.; Sotiri, E.; Treberg, J.R. Mitochondrial reactive oxygen species production by fish muscle mitochondria: Potential role in acute heat-induced oxidative stress. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 191, 99–107. [Google Scholar] [CrossRef]
- Comporti, M. Free radicals, oxidative stress and antioxidants. J. Siena Acad. Sci. 2010, 2, 13–26. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef]
- Kütter, M.; Romano, L.; Ventura-Lima, J.; Tesser, M.; Monserrat, J. Antioxidant and toxicological effects elicited by alpha-lipoic acid in aquatic organisms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 162, 70–76. [Google Scholar] [CrossRef]
- Wang, X.; Xu, W.; Zhou, H.; Zhang, Y.; Gao, W.; Zhang, W.; Mai, K. Reduced glutathione supplementation in practical diet improves the growth, anti-oxidative capacity, disease resistance and gut morphology of shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 73, 152–157. [Google Scholar] [CrossRef]
- Wu, S.M.; Liu, J.-H.; Shu, L.-H.; Chen, C.H. Anti-oxidative responses of zebrafish (Danio rerio) gill, liver and brain tissues upon acute cold shock. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 187, 202–213. [Google Scholar] [CrossRef]
- Faria, S.C.; Klein, R.D.; Costa, P.G.; Crivellaro, M.S.; Santos, S.; de Siqueira Bueno, S.L.; Bianchini, A. Phylogenetic and environmental components of inter-specific variability in the antioxidant defense system in freshwater anomurans Aegla (Crustacea, Decapoda). Sci. Rep. 2018, 8, 2850. [Google Scholar] [CrossRef]
- Shakya, S.R. Medicinal uses of ginger (Zingiber officinale Roscoe) improves growth and enhances immunity in aquaculture. Int. J. Chem. Stud. 2015, 3, 83–87. [Google Scholar]
- Awad, E.; Awaad, A. Role of medicinal plants on growth performance and immune status in fish. Fish Shellfish Immunol. 2017, 67, 40–54. [Google Scholar] [CrossRef]
- de Araújo, F.F.; de Paulo Farias, D.; Neri-Numa, I.A.; Pastore, G.M. Polyphenols and their applications: An approach in food chemistry and innovation potential. Food Chem. 2021, 338, 127535. [Google Scholar] [CrossRef]
- Nakamura, K.; Ishiyama, K.; Sheng, H.; Ikai, H.; Kanno, T.; Niwano, Y. Bactericidal activity and mechanism of photoirradiated polyphenols against gram-positive and-negative bacteria. J. Agric. Food Chem. 2015, 63, 7707–7713. [Google Scholar] [CrossRef]
- Wickramasingha, W.; Wijendra, W.; Karunaratne, D.; Liyanapathirana, V.; Ekanayake, E.M.; Jayasinghe, S.; Karunaratne, V. Antibacterial, antifungal, antioxidant, brine shrimp lethality and polyphenolic content of Holarrhena mitis (Vahl) R. Br. ex Roem. & Schult. Ceylon J. Sci. 2018, 47, 269–274. [Google Scholar]
- Wink, M.; Schimmer, O. Modes of action of defensive secondary metabolites. In Annual Plant Reviews Online; Wiley: Hoboken, NJ, USA, 2018; pp. 18–137. [Google Scholar]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Landete, J. Updated knowledge about polyphenols: Functions, bioavailability, metabolism, and health. Crit. Rev. Food Sci. Nutr. 2012, 52, 936–948. [Google Scholar] [CrossRef]
- D Archivio, M.; Filesi, C.; Di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, dietary sources and bioavailability. Ann.-Ist. Super. Sanita 2007, 43, 348. [Google Scholar]
- da Silva Martins, Á.C.; Artigas Flores, J.; Porto, C.; Romano, L.A.; Wasielesky Junior, W.; Caldas, S.S.; Primel, E.G.; Külkamp-Guerreiro, I.; Monserrat, J.M. Antioxidant effects of nanoencapsulated lipoic acid in tissues and on the immune condition in haemolymph of Pacific white shrimp Litopenaeus vannamei (Boone, 1931). Aquac. Nutr. 2018, 24, 1255–1262. [Google Scholar] [CrossRef]
- Sheikh Asadi, M.; Gharaei, A.; Mirdar Harijani, J.; Arshadi, A. A Comparison between dietary effects of Cuminum cyminum essential oil and Cuminum cyminum essential oil, loaded with iron nanoparticles, on growth performance, immunity and antioxidant indicators of white leg shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1466–1473. [Google Scholar] [CrossRef]
- Kesselring, J.; Gruber, C.; Standen, B.; Wein, S. Effect of a phytogenic feed additive on the growth performance and immunity of Pacific white leg shrimp, fed a low fishmeal diet. J. World Aquac. Soc. 2021, 52, 303–315. [Google Scholar] [CrossRef]
- Sang, H.M.; Kien, N.T.; Thanh Thuy, N.T. Effects of dietary mannan oligosaccharide on growth, survival, physiological, immunological and gut morphological conditions of black tiger prawn (Penaeus monodon Fabricius 1798). Aquac. Nutr. 2014, 20, 341–348. [Google Scholar] [CrossRef]
- Boyd, C.; McNevin, A.A. Overview of aquaculture feeds: Global impacts of ingredient production, manufacturing, and use. In Feed and Feeding Practices in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2022; pp. 3–28. [Google Scholar]
- Lara, G.; Hostins, B.; Bezerra, A.; Poersch, L.; Wasielesky, W. The effects of different feeding rates and re-feeding of Litopenaeus vannamei in a biofloc culture system. Aquac. Eng. 2017, 77, 20–26. [Google Scholar] [CrossRef]
- FAO. Aquaculture Feed and Fertilizer Resources Information System. In Species Profile; Food and Agriculture Organization of the United Nations: Rome, Italy, 2024. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005; Volume 6. [Google Scholar]
- Servin Arce, K.; de Souza Valente, C.; do Vale Pereira, G.; Shapira, B.; Davies, S.J. Modulation of the gut microbiota of Pacific white shrimp (Penaeus vannamei Boone, 1931) by dietary inclusion of a functional yeast cell wall-based additive. Aquac. Nutr. 2021, 27, 1114–1127. [Google Scholar] [CrossRef]
- Zhou, Q.-C.; Li, C.-C.; Liu, C.-W.; Chi, S.-Y.; Yang, Q.-H. Effects of dietary lipid sources on growth and fatty acid composition of juvenile shrimp, Litopenaeus vannamei. Aquac. Nutr. 2007, 13, 222–229. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. [59] Glutathione reductase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1985; pp. 484–490. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Fleischner, G.; Gatmaitan, Z.; Arias, I.M.; Jakoby, W.B. The Identity of Glutathione -S-Transferase B with Ligandin, a Major Binding Protein of Liver. Proc. Natl. Acad. Sci. USA 1974, 71, 3879–3882. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Wendel, A. [44] Glutathione peroxidase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1981; pp. 325–333. [Google Scholar]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Federici, G.; Shaw, B.J.; Handy, R.D. Toxicity of titanium dioxide nanoparticles to rainbow trout (Oncorhynchus mykiss): Gill injury, oxidative stress, and other physiological effects. Aquat. Toxicol. 2007, 84, 415–430. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Yousefi, M.; Karimi, M.; Fadaei Raieni, R.; Dadar, M.; Yilmaz, S.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Benefits of Dietary Polyphenols and Polyphenol-Rich Additives to Aquatic Animal Health: An Overview. Rev. Fish. Sci. Aquac. 2021, 29, 478–511. [Google Scholar] [CrossRef]
- Ahmadi, A.; Bagheri, D.; Hoseinifar, S.H.; Morshedi, V.; Paolucci, M. Beneficial role of polyphenols as feed additives on growth performances, immune response and antioxidant status of Lates Calcarifer (Bloch, 1790) juveniles. Aquaculture 2022, 552, 737955. [Google Scholar] [CrossRef]
- Niyamosatha, H.; Chuchird, N.; Rairat, T. Effect of Dietary Polyphenol-Rich Feed Additive from Grape Pomace on Growth, Survival and Tolerance to Vibrio Infection in Pacific White Shrimp (Litopenaeus vannamei). J. Fish. Environ. 2015, 39, 1–9. [Google Scholar]
- Niti Chuchird, H.N. Tirawat Rairat and Arunothai Keetanon. Effect of Dietary Phytobiotics Products on Growth, Immune Responses and Vibriosis Resistance in Litopenaeus vannamei. J. Fish. Aquat. Sci. 2017, 12, 184–190. [Google Scholar] [CrossRef]
- Ruiz-Velazco, J.M.J.; González-Romero, M.A.; Estrada-Perez, N.; Hernandez-Llamas, A. Evaluating partial harvesting strategies for whiteleg shrimp Litopenaeus (Penaeus) vannamei semi-intensive commercial production: Profitability, uncertainty, and economic risk. Aquac. Int. 2021, 29, 1317–1329. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Parameters | Values |
|---|---|
| Humidity and volatiles (%) | 9.16 ± 1.15 |
| Crude protein (%) | 39.90 ± 1.95 |
| Ethereal extract by acid hydrolysis (%) | 10.25 ± 1.26 |
| Crude fiber (%) | 2.39 ± 0.60 |
| Mineral matter (%) | 13.14 ± 0.92 |
| Calcium (%) | 3.45 ± 0.54 |
| Nitrogen-free extract (%) | 38.54 ± 3.81 |
| Phosphorus (%) | 1.81 ± 0.35 |
| Sodium (%) | 0.48 ± 0.12 |
| Potassium (%) | 1.33 ± 0.30 |
| Acid index (mg NaOH g−1) | 0.91 ± 0.28 |
| Peroxide index (meq kg−1) | 0.00 |
| Growth Parameters | Experimental Groups (g kg−1 of ELIFE®) | p Value | ||
|---|---|---|---|---|
| 0.0 | 0.5 | 1.0 | ||
| Initial average weight (g) | 1.76 ± 0.16 | 1.76 ± 0.16 | 1.76 ± 0.16 | N/A |
| Final average weight (g) | 5.35 ± 0.65 b | 5.80 ± 0.53 a | 5.79 ± 0.63 a | <0.001 |
| Weight gain (g) | 3.59 ± 0.49 b | 4.04 ± 0.37 a | 4.03 ± 0.47 a | <0.001 |
| Final average length (cm) | 8.08 ± 0.54 b | 8.34 ± 0.20 a | 8.45 ± 0.61 a | <0.001 |
| Survival (%) | 82.50 ± 16.69 | 85.00 ± 14.14 | 92.50 ± 14.88 | 0.7543 |
| Final biomass (g) | 17.74 ± 2.88 b | 20.81 ± 2.44 ab | 23.06 ± 2.82 a | <0.001 |
| SGR (% d−1) | 3.35 ± 0.05 b | 3.62 ± 0.06 a | 3.57 ± 0.04 a | <0.001 |
| FCR (g g−1) | 1.86 ± 0.15 a | 1.49 ± 0.29 ab | 1.24 ± 0.39 b | <0.001 |
| Variable | Experimental Groups (g kg−1 of ELIFE®) | p Value | ||
|---|---|---|---|---|
| 0.0 | 0.5 | 1.0 | ||
| Initial average weight (g) | 5.58 ± 0.41 | 5.58 ± 0.41 | 5.58 ± 0.41 | N/A |
| Final average weight (g) | 9.40 ± 2.5 | 9.11 ± 1.33 | 10.09 ± 0.41 | 0.2853 |
| Weight gain (g) | 3.82 ± 2.09 | 3.53 ± 0.92 | 4.51 ± 0.01 | 0.2853 |
| Final average length (cm) | 10.11 ± 1.23 | 10.04 ± 0.80 | 10.03 ± 1.0 | 0.9583 |
| Survival (%) | 70.24 ± 1.48 c | 77.35 ± 0.76 b | 85.88 ± 1.45 a | <0.001 |
| Final biomass (g) | 33.63 ± 2.93 b | 34.98 ± 1.99 b | 41.04 ± 1.04 a | 0.0019 |
| SGR (%) | 1.00 ± 0.25 b | 1.06 ± 0.16 b | 1.38 ± 0.14 a | 0.0215 |
| FCR (g g−1) | 1.68 ± 0.15 a | 1.23 ± 0.29 b | 1.12 ± 0.45 b | 0.0026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira Colares, H.D.; de Souza Valente, C.; Zadinelo, I.V.; do Nascimento Ferreira, C.H.; Retcheski, M.C.; Cazarolli, L.H.; Rosseel, L.; Ballester, E.L.C. Effect of a Commercial Polyphenol Compound on the Performance and Antioxidant Status of Penaeus vannamei. Fishes 2024, 9, 410. https://doi.org/10.3390/fishes9100410
Ferreira Colares HD, de Souza Valente C, Zadinelo IV, do Nascimento Ferreira CH, Retcheski MC, Cazarolli LH, Rosseel L, Ballester ELC. Effect of a Commercial Polyphenol Compound on the Performance and Antioxidant Status of Penaeus vannamei. Fishes. 2024; 9(10):410. https://doi.org/10.3390/fishes9100410
Chicago/Turabian StyleFerreira Colares, Hallypher Deyrrikson, Cecília de Souza Valente, Izabel Volkweis Zadinelo, Caio Henrique do Nascimento Ferreira, Milena Cia Retcheski, Luisa Helena Cazarolli, Lodewijk Rosseel, and Eduardo Luis Cupertino Ballester. 2024. "Effect of a Commercial Polyphenol Compound on the Performance and Antioxidant Status of Penaeus vannamei" Fishes 9, no. 10: 410. https://doi.org/10.3390/fishes9100410
APA StyleFerreira Colares, H. D., de Souza Valente, C., Zadinelo, I. V., do Nascimento Ferreira, C. H., Retcheski, M. C., Cazarolli, L. H., Rosseel, L., & Ballester, E. L. C. (2024). Effect of a Commercial Polyphenol Compound on the Performance and Antioxidant Status of Penaeus vannamei. Fishes, 9(10), 410. https://doi.org/10.3390/fishes9100410