
Effects of Different Protein Sources on Growth Performance, Muscle Flavor Substances and Quality Structure in Triploid Crucian Carp

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets and Procedures
2.2. Fish, Cage Culture, and Experimental Procedures
2.3. Sample Collection
2.4. Biochemical Composition
2.5. Determination of Amino Acids in Muscle
2.6. Assay of Muscle Textural Properties
2.7. Detection of Plasma and Intestinal Enzyme Activity
2.8. RNA Isolation and cDNA Synthesis
2.9. Real-Time Polymerase Chain Reaction (PCR) Analysis
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Biochemical Composition
3.3. Free Amino Acids
3.4. Muscle Texture Properties
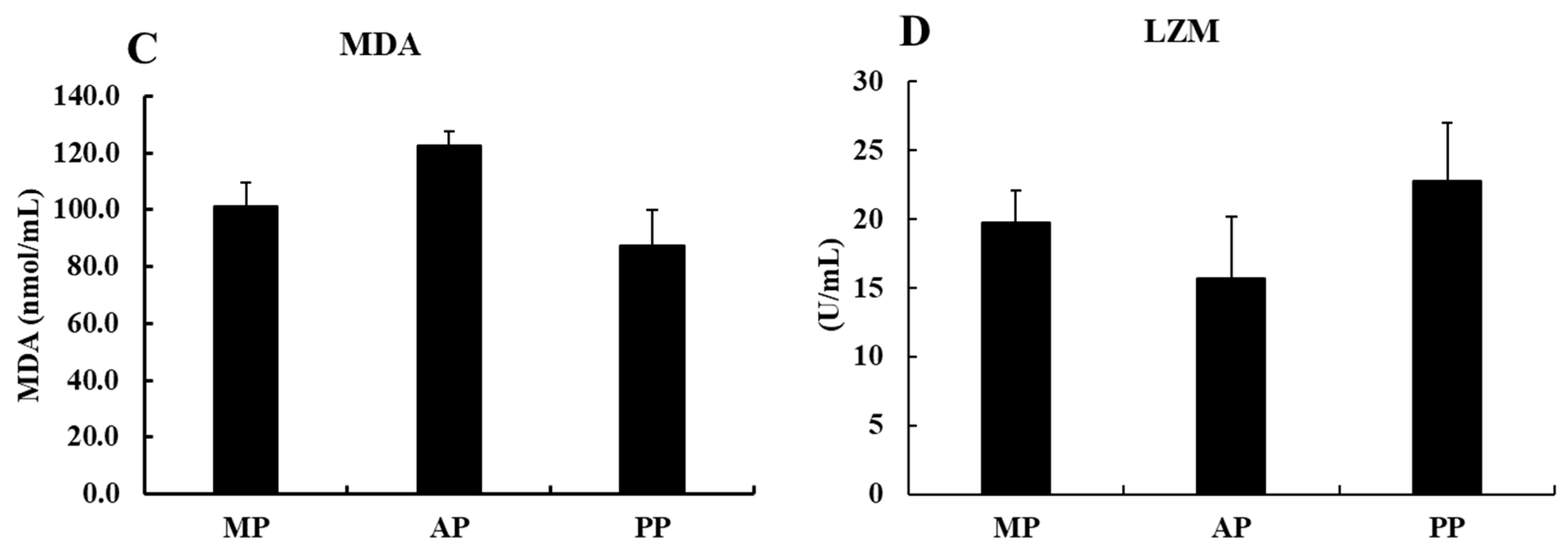
3.5. Antioxidant and Intestinal Digestive Enzymes
3.6. The Expression of Genes in the TOR Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, S.; Mo, P.; Xiao, Y.; Chen, Y.; Shi, Y.; Hu, Y.; Tang, J.; Qu, F.; Luo, M.; Ai, X.; et al. Dietary supplementation with fermented plant meal enhances growth, antioxidant capacity and expression of TOR signaling pathway genes in gibel carp (Carassius auratus gibelio var. CAS V). Aquac. Rep. 2021, 19, 100559. [Google Scholar] [CrossRef]
- Zhang, C.; Rahimnejad, S.; Wang, Y.; Lu, K.; Song, K.; Wang, L.; Mai, K. Substituting fish meal with soybean meal in diets for Japanese seabass (Lateolabrax japonicus): Effects on growth, digestive enzymes activity, gut histology, and expression of gut inflammatory and transporter genes. Aquaculture 2018, 483, 173–182. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, Y.; Ji, H.; Yu, E. The effect of replacing fish meal with fermented meal mixture of silkworm pupae, rapeseed and wheat on growth, body composition and health of mirror carp (Cyprinus carpio var. Specularis). Aquac. Nutr. 2017, 23, 741–754. [Google Scholar] [CrossRef]
- Mamauag, R.E.P.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Gao, J.; Nguyen, B.T.; Ragaza, J.A. Soy peptide inclusion levels influence the growth performance, proteolytic enzyme activities, blood biochemical parameters and body composition of Japanese flounder (Paralichthys olivaceus). Aquaculture 2011, 321, 252–258. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Laining, A.; Rachmansyah; Ahmad, T.; Williams, K. Apparent digestibility of selected feed ingredients for humpback grouper (Cromileptes altivelis). Aquaculture 2003, 218, 529–538. [Google Scholar] [CrossRef]
- Li, H.; Mai, K.; Ai, Q.; Zhang, L.; Zhang, C.; Zhang, W.; Liufu, Z. Apparent digestibility of selected protein ingredients for larger yellow croaker (Pseudosciaena crocea). Acta Hydrobiol. Sin. 2007, 31, 370–376. [Google Scholar]
- Pervin, M.A.; Jahan, H.; Akter, R.; Omri, A.; Hossain, Z. Appraisal of different levels of soybean meal in diets on growth, digestive enzyme activity, antioxidation, and gut histology of tilapia (Oreochromis niloticus). Fish Physiol. Biochem. 2020, 46, 1397–1407. [Google Scholar] [CrossRef]
- Mohammadinafchi, F.; Mohammadiazarm, H.; Yavari, V. Evaluation effect of soybean meal and baker’syeast on resistance to anoxia stress and blood biochemical parameters of fingerlings (Mesopotamichthys sharpeyi Günther, 1874). Int. J. Biosci. 2014, 5, 215–222. [Google Scholar]
- Yu, D.; Gong, S.; Yuan, Y.; Lin, Y. Effects of replacing fish meal with soybean meal on growth, body composition and digestive enzyme activities of juvenile Chinese sucker (Myxocyprinus asiaticus). Aquac. Nutr. 2013, 19, 84–90. [Google Scholar] [CrossRef]
- Luo, Y.; Ai, Q.; Mai, K.; Zhang, W.; Xu, W.; Zhang, Y. Effects of dietary rapeseed meal on growth performance, digestion and protein metabolism in relation to gene expression of juvenile cobia (Rachycentron canadum). Aquaculture 2012, 368–369, 109–116. [Google Scholar] [CrossRef]
- Cheng, Z.; Ai, Q.; Mai, K.; Xu, W.; Ma, H.; Li, Y.; Zhang, J. Effects of dietary canola meal on growth performance, digestion and metabolism of Japanese seabass (Lateolabrax japonicus). Aquaculture 2010, 305, 102–108. [Google Scholar] [CrossRef]
- Lee, S.M.; Azarm, H.M.; Chang, K.H. Effects of dietary inclusion of fermented soybean meal on growth, body composition, antioxidant enzyme activity and disease resistance of rockfish (Sebastes schlegeli). Aquaculture 2016, 459, 110–116. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, Y.; Wu, T.; Chu, W. Effects of high dietary levels of cottonseed meal and rapeseed meal on growth performance, muscle texture, and expression of muscle-related genes in grass carp. N. Am. J. Aquac. 2019, 81, 235–241. [Google Scholar] [CrossRef]
- Xu, J.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Zhou, X. Different dietary protein levels affect flesh quality, fatty acids and alter gene expression of Nrf2-mediated antioxidant enzymes in the muscle of grass carp (Ctenopharyngodon idella). Aquaculture 2018, 493, 272–282. [Google Scholar] [CrossRef]
- Ma, X.; Feng, L.; Wu, P.; Liu, Y.; Kuang, S.; Tang, L.; Zhou, X.; Jiang, W. Enhancement of flavor and healthcare substances, mouthfeel parameters and collagen synthesis in the muscle of on-growing grass carp (Ctenopharyngodon idella) fed with graded levels of glutamine. Aquaculture 2020, 528, 735486. [Google Scholar] [CrossRef]
- Peng, K.; Fu, B.; Li, J.; Zhao, H.; Cao, J.; Huang, W.; Chen, B.; Li, X.; Peng, Z.; Wei, M. Effects of replacing soybean meal and rapeseed meal with faba bean meal on growth performance and muscle quality of tilapia (Oreochromis niloticus). Aquac. Rep. 2022, 26, 101328. [Google Scholar] [CrossRef]
- Grabež, V.; Egelandsdal, B.; Kjos, N.P.; Håkenåsen, I.M.; Mydland, L.T.; Vik, J.O.; Hallenstvedt, E.; Devle, H.; Øverland, M. Replacing soybean meal with rapeseed meal and faba beans in a growing-finishing pig diet: Effect on growth performance, meat quality and metabolite changes. Meat Sci. 2020, 166, 108134. [Google Scholar] [CrossRef]
- Mao, Z.; Luo, S.; Zhao, D.; Zhou, X.; Zhang, Z.; Xiao, Y.; Cao, S.; Zhou, Y.; Liu, S.; Tang, J.; et al. Molecular characterization and nutrition regulation of the neutral amino acid transporter ASCT2 in Triploid Crucian Carp. Fishes 2021, 6, 77. [Google Scholar] [CrossRef]
- Cai, L.; Ao, Z.; Tang, T.; Tong, F.; Wei, Z.; Yang, F.; Shu, Y.; Liu, S.; Mai, K. Corrigendum to “Characterization of difference in muscle volatile compounds between triploid and diploid crucian carp” [Aquacult. Rep. 20 (July) (2021) 100641]. Aquac. Rep. 2021, 20, 100689. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemist, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2005. [Google Scholar]
- Cao, S.; Xiong, D.; Luo, W.; Tang, J.; Qu, F.; Zhou, Y.; He, Z.; Xie, S.; Liu, Z. Effects of dietary soy isoflavones on growth, antioxidant status, immune response and resistance of juvenile grass carp (Ctenopharyngodon idella) to aeromonas hydrophila challenge. Aquac. Res. 2020, 51, 2472–2482. [Google Scholar] [CrossRef]
- Qu, F.; Tang, J.; Hu, R.; Hao, G.; Zhou, Y.; Lu, S.; Zhao, Q.; You, C.; Li, Y.; Zhou, Z.; et al. Molecular characterization and nutritional regulation of specificity protein 1 (Sp1) in grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2018, 24, 940–951. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.T.; Wade, M.N.; Pirozzi, I.; Glencross, B.D. Effects of canola meal on growth, feed utilisation, plasma biochemistry, histology of digestive organs and hepatic gene expression of barramundi (Asian seabass; Lates calcarifer). Aquaculture 2016, 464, 95–105. [Google Scholar] [CrossRef]
- Zhou, Q.L.; Habte-Tsion, H.M.; Ge, X.; Xie, J.; Ren, M.; Liu, B.; Miao, L.; Pan, L. Graded replacing fishmeal with canola meal in diets affects growth and target of rapamycin pathway gene expression of juvenile blunt snout bream (Megalobrama amblycephala). Aquac. Nutr. 2018, 24, 300–309. [Google Scholar] [CrossRef]
- Lovell, R.T. Nutrition and Feeding of Fish; Van Nostrand Reinhold: New York, NY, USA, 1989; 268p. [Google Scholar]
- Luo, Z.; Liu, Y.; Mai, K.; Tian, L.; Tan, X.; Yang, H.; Liang, G.; Liu, D. Quantitative L-lysine requirement of juvenile grouper Epinephelus coioides. Aquac. Nutr. 2006, 12, 165–172. [Google Scholar] [CrossRef]
- Feng, L.; Feng, L.; Jiang, W.; Liu, Y.; Zhang, L.; Kuang, S.; Ren, H.; Jin, X.; Li, S.; Mi, H.; et al. The beneficial effects of exogenous protease K originated from Parengyodontium album on growth performance of grass carp (Ctenopharyngodon idella) in relation to the enhanced intestinal digestion and absorption capacities. Aquaculture 2023, 563, 738929. [Google Scholar] [CrossRef]
- Mir, I.N.; Srivastava, P.P.; Bhat, I.A.; Muralidhar, A.P.; Varghese, T.; Gireesh-Babu, P.; Jain, K.K. Expression and activity of trypsin and pepsin during larval development of Indian walking catfish (Clarias magur). Aquaculture 2018, 491, 266–272. [Google Scholar] [CrossRef]
- Maulu, S.; Hualiang, L.; Ke, J.; Ren, M.; Ge, X.; Huang, D.; Yu, H. Dietary clostridium autoethanogenum protein modulates intestinal absorption, antioxidant status, and immune response in GIFT (Oreochromis niloticus) juveniles. Aquac. Res. 2021, 52, 5787–5799. [Google Scholar] [CrossRef]
- Liu, X.; Han, B.; Xu, J.; Zhu, J.; Hu, J.; Wan, W.; Miao, S. Replacement of fishmeal with soybean meal affects the growth performance, digestive enzymes, intestinal microbiota and immunity of Carassius auratus gibelio♀ × Cyprinus carpio♂. Aquac. Rep. 2020, 18, 100472. [Google Scholar] [CrossRef]
- Ye, H.; Xu, M.; Liu, Q.; Sun, Z.; Zou, C.; Chen, L.; Su, N.; Ye, C. Effects of replacing fish meal with soybean meal on growth performance, feed utilization and physiological status of juvenile obscure puffer (Takifugu obscurus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 216, 75–81. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M.; Hamad, S.M.; Gomaa, M. Protective effects of spirulina on the liver function and hyperlipidemia of rats and human. Braz. Arch. Biol. Technol. 2014, 57, 77–86. [Google Scholar] [CrossRef]
- Saito, M.; Saito, K.; Kunisaki, N.; Kimura, S. Green tea polyphenols inhibit metalloproteinase activities in the skin, muscle, and blood of rainbow trout. J. Agric. Food Chem. 2002, 50, 7169–7174. [Google Scholar] [CrossRef] [PubMed]
- Seiliez, I.; Panserat, S.; Skiba-Cassy, S.; Fricot, A.; Vachot, C.; Kaushik, S.; Tesseraud, S. Feeding status regulates the polyubiquitination step of the ubiquitin-proteasome-dependent proteolysis in rainbow trout (Oncorhynchus mykiss) muscle. J. Nutr. 2008, 138, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Jiang, G.; Cheng, H.; Cao, X.; Wang, C.; Dai, Y.; Liu, W. Replacing fish meal with cottonseed meal protein hydrolysate affects growth, intestinal function, and growth hormone/insulin-like growth factor I axis of juvenile blunt snout bream (Megalobrama amblycephala). J. World Aquac. Soc. 2020, 51, 1235–1249. [Google Scholar] [CrossRef]
- Savolainen, L.; Gatlin, D.M., III. Evaluation of sulfur amino acid and zinc supplements to soybean-meal-based diets for hybrid striped bass. Aquaculture 2010, 307, 260–265. [Google Scholar] [CrossRef]
- Mai, K.; Wan, J.; Ai, Q.; Xu, W.; Liufu, Z.; Zhang, L.; Zhang, C.; Li, H. Dietary methionine requirement of large yellow croaker (Pseudosciaena crocea R). Aquaculture 2006, 253, 564–572. [Google Scholar] [CrossRef]
- Wu, F.; Tian, J.; Yu, L.; Wen, H.; Jiang, M.; Lu, X. Effects of dietary rapeseed meal levels on growth performance, biochemical indices and flesh quality of juvenile genetically improved farmed tilapia. Aquac. Rep. 2021, 20, 100679. [Google Scholar] [CrossRef]
- Liu, T.; Han, T.; Wang, J.; Liu, T.; Bian, P.; Wang, Y.; Cai, X. Effects of replacing fish meal with soybean meal on growth performance, feed utilization and physiological status of juvenile redlip mullet Liza haematocheila. Aquac. Rep. 2021, 20, 100756. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, W.; Wu, P.; Liu, Y.; Zeng, Y.; Jiang, J.; Kuang, S.; Tang, L.; Tang, W.; Wang, S.; et al. Soybean isoflavones improve the health benefits, flavour quality indicators and physical properties of grass carp (Ctenopharygodon idella). PLoS ONE 2019, 14, e0209570. [Google Scholar] [CrossRef]
- Lin, H.; Fang, J.; Bi, X.; Yu, X.; He, Q.; Liu, P.; Ding, W.; Che, Z. Optimization of the extraction technique of free amino acids from Pixian board-bean paste by response surface methodology and analysis of their taste characteristics. Sci. Technol. Food Ind. 2019, 40, 56–63. [Google Scholar]
- Ginés, R.; Valdimarsdottir, T.; Sveinsdottir, K.; Thorarensen, H. Effects of rearing temperature and strain on sensory characteristics, texture, colour and fat of Arctic charr (Salvelinus alpinus). Food Qual. Prefer. 2004, 15, 177–185. [Google Scholar] [CrossRef]
- Bugeon, J.; Lefevre, F.; Fauconneau, B. Fillet texture and muscle structure in brown trout (Salmo trutta) subjected to long-term exercise. Aquac. Res. 2003, 34, 1287–1295. [Google Scholar] [CrossRef]
- Johnston, I.; Li, X.; Vieira, V.; Nickell, D.; Dingwall, A.; Alderson, R.; Campbell, P.; Bickerdike, R. Muscle and flesh quality traits in wild and farmed Atlantic salmon. Aquaculture 2006, 256, 323–336. [Google Scholar] [CrossRef]
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Mokrani, A.; Ren, M.; Liang, H.; Yang, Q.; Ji, K.; Kasiya, H.; Ge, X. Effect of the total replacement of fishmeal with plant proteins and supplemental essential amino acids in the extruded diet on antioxidants genes, enzyme activities, and immune response in juvenile blunt snout bream. Aquac. Int. 2019, 28, 555–568. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Peña-Llopis, S.; Gómez-Requeni, P.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J. Effect of fish meal replacement by plant protein sources on non-specific defence mechanisms and oxidative stress in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 387–400. [Google Scholar] [CrossRef]
- Rajan, V.K.; Muraleedharan, K. A computational investigation on the structure, global parameters and antioxidant capacity of a polyphenol (Gallic acid). Food Chem. 2017, 220, 93–99. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Mixed Protein Group (MP) | Animal Protein Group (AP) | Plant Derived Protein Group (PP) |
|---|---|---|---|
| Fishmeal 1 | 12.00 | 44.40 | 0.00 |
| Soybean meal 1 | 20.00 | 0.00 | 37.10 |
| Rapeseed meal 1 | 15.00 | 0.00 | 15.00 |
| Casein 1 | 6.50 | 0.00 | 6.50 |
| Fish oil l | 3.00 | 1.63 | 3.50 |
| Soybean oil l | 3.00 | 1.63 | 3.50 |
| Corn starch | 16.80 | 31.00 | 10.00 |
| Wheat flour | 10.00 | 10.00 | 10.00 |
| Choline chloride | 0.50 | 0.50 | 0.50 |
| Premix 2 | 3.00 | 3.00 | 3.00 |
| CMC 3 | 3.00 | 3.00 | 3.00 |
| Cellulose | 7.20 | 4.84 | 7.90 |
| Total | 100.00 | 100.00 | 100.00 |
| Proximate composition | |||
| Crude protein | 32.68 | 32.68 | 32.28 |
| Crude lipid | 8.00 | 7.80 | 8.66 |
| Moisture | 10.05 | 9.73 | 9.86 |
| Ash | 7.54 | 10.09 | 6.27 |
| Gross energy (MJ kg−1) | 18.06 | 18.07 | 18.00 |
| Acronym | Primer Sequence | Accession No. | Annealing Temp. (°C) |
|---|---|---|---|
| TOR | F: TCAGGGTTGTCAGCGTATTG R: AGGGTTTTATGGGCTAGTGC | KF772613 | 60 |
| IGF1 | F: ATTGCCCGCATCTCATCCTC R: TGACCGCTAGACATCCCCTT | KF813006 | 60 |
| S6K1 | F: CGAGCTGGAGTTAATAGGGTT R: AGGTGACATGCACCATCTATG | KF880601 | 57 |
| 4E-BP2 | F: CACTTTATTCTCCACCACCC R: GATGTTGTTAGCCTCATTCCT | KF900277 | 60 |
| β-actin | F: TTGAGCAGGAGATGGGAACCG R: AGAGCCTCAGGGCAACGGAAA | AB039726.2 | 60 |
| Items | MP | AP | PP |
|---|---|---|---|
| IBW 1 | 11.83 ± 0.07 | 11.69 ± 0.08 | 11.72 ± 0.11 |
| FBW 2 | 24.26 ± 0.33 b | 27.75 ± 0.28 c | 22.80 ± 0.33 a |
| WGR 3 | 105.11 ± 3.84 b | 137.32 ± 3.01 c | 94.57 ± 1.07 a |
| SGR 4 | 1.20 ± 0.03 b | 1.44 ± 0.02 c | 1.11 ± 0.01 a |
| FE 5 | 41.35 ± 2.64 a | 53.61 ± 0.30 b | 39.75 ± 1.51 a |
| FR 6 | 2.58 ± 0.07 ab | 2.49 ± 0.01 a | 2.69 ± 0.04 b |
| CF 7 | 1.98 ± 0.08 ab | 2.08 ± 0.03 b | 1.88 ± 0.04 a |
| VSI 8 | 10.44 ± 0.18 | 10.49 ± 0.63 | 9.21 ± 0.38 |
| Item | MP | AP | PP |
|---|---|---|---|
| Whole body of fish | |||
| Crude protein | 14.34 ± 0.36 | 15.06 ± 0.07 | 15.00 ± 0.16 |
| Crude lipid | 10.88 ± 0.43 b | 11.37 ± 0.27 b | 9.61 ± 0.31 a |
| Moisture | 70.65 ± 0.63 ab | 70.09 ± 0.27 a | 71.89 ± 0.21 b |
| Ash | 2.39 ± 0.03 | 2.33 ± 0.05 | 2.49 ± 0.06 |
| Dorsal muscle | |||
| Crude protein | 19.47 ± 0.27 ab | 18.80 ± 0.20 a | 20.33 ± 0.34 b |
| Crude lipid | 1.36 ± 0.12 b | 1.10 ± 0.14 ab | 0.72 ± 0.07 a |
| Moisture | 78.59 ± 0.48 ab | 77.69 ± 0.81 a | 81.43 ± 1.46 b |
| Ash | 1.11 ± 0.07 | 1.20 ± 0.03 | 1.09 ± 0.19 |
| Item | MP | AP | PP |
|---|---|---|---|
| Flavor amino acid | |||
| Glycine | 256.63 ± 18.03 a | 363.19 ± 13.89 b | 271.4 ± 17.10 a |
| Alanine | 250.35 ± 20.19 | 273.42 ± 3.69 | 254.28 ± 23.30 |
| Glutamic acid | 208.89 ± 33.67 | 189.75 ± 21.47 | 177.14 ± 11.70 |
| Tyrosine | 16.99 ± 4.15 | 15.19 ± 7.48 | 11.68 ± 2.05 |
| Aspartic acid | 305.27 ± 16.42 | 201.46 ± 94.55 | 202.82 ± 85.78 |
| Serine | 74.84 ± 3.16 | 72.38 ± 29.59 | 66.21 ± 18.44 |
| Other free amino acids | |||
| Histidine | 3260.22 ± 75.45 ab | 3033.35 ± 119.37 a | 3433.52 ± 55.73 b |
| Arginine | 2675.69 ± 230.78 | 2923.03 ± 18.16 | 3063.52 ± 43.56 |
| Threonine | 115.58 ± 24.07 | 117.27 ± 16.43 | 79.75 ± 14.29 |
| Valine | 30.82 ± 11.43 | 25.01 ± 14.21 | 18.97 ± 4.22 |
| Methionine | 35.76 ± 3.05 b | 45.84 ± 5.21 b | 20.64 ± 2.13 a |
| Isoleucine | 13.91 ± 5.45 | 10.21 ± 6.78 | 5.24 ± 1.90 |
| Leucine | 27.89 ± 9.70 | 24.47 ± 13.81 | 12.83 ± 2.85 |
| Lysine | 635.78 ± 272.75 a | 1350.54 ± 178.08 b | 516.26 ± 61.36 a |
| Proline | 276.93 ± 12.07 b | 193.97 ± 13.86 a | 220.36 ± 19.89 a |
| Item | MP | AP | PP |
|---|---|---|---|
| Springiness (mm) | 0.67 ± 0.04 | 0.70 ± 0.03 | 0.69 ± 0.03 |
| Hardness (N) | 18.65 ± 1.18 a | 17.51 ± 0.91 a | 21.49 ± 0.76 b |
| Gumminess (g × mm) | 3.10 ± 0.21 a | 2.90 ± 0.20 a | 3.98 ± 0.23 b |
| Chewiness (mJ) | 2.14 ± 0.25 ab | 2.06 ± 0.20 a | 2.75 ± 0.21 b |
| Adhesiveness (N × mm) | 0.029 ± 0.006 | 0.039 ± 0.007 | 0.027 ± 0.005 |
| Cohesiveness (%) | 0.170 ± 0.012 | 0.167 ± 0.009 | 0.185 ± 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Yi, C.; Mo, Y.; He, Z.; Xu, Z.; He, Y.; Ouyang, Y.; Mao, Z.; Qu, F.; Tang, J.; et al. Effects of Different Protein Sources on Growth Performance, Muscle Flavor Substances and Quality Structure in Triploid Crucian Carp. Fishes 2024, 9, 23. https://doi.org/10.3390/fishes9010023
Yang L, Yi C, Mo Y, He Z, Xu Z, He Y, Ouyang Y, Mao Z, Qu F, Tang J, et al. Effects of Different Protein Sources on Growth Performance, Muscle Flavor Substances and Quality Structure in Triploid Crucian Carp. Fishes. 2024; 9(1):23. https://doi.org/10.3390/fishes9010023
Chicago/Turabian StyleYang, Liquan, Chenglin Yi, Yujian Mo, Zhimin He, Zhehua Xu, Yimiao He, Yongkang Ouyang, Zhuangwen Mao, Fufa Qu, Jianzhou Tang, and et al. 2024. "Effects of Different Protein Sources on Growth Performance, Muscle Flavor Substances and Quality Structure in Triploid Crucian Carp" Fishes 9, no. 1: 23. https://doi.org/10.3390/fishes9010023
APA StyleYang, L., Yi, C., Mo, Y., He, Z., Xu, Z., He, Y., Ouyang, Y., Mao, Z., Qu, F., Tang, J., Liu, Z., Fang, Z., & Cao, S. (2024). Effects of Different Protein Sources on Growth Performance, Muscle Flavor Substances and Quality Structure in Triploid Crucian Carp. Fishes, 9(1), 23. https://doi.org/10.3390/fishes9010023