

The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum barcoo Fingerlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Fish
2.3. Sampling
2.4. Biochemical Measurement of the Serum
2.5. Lipid Peroxidation Enzyme Activity Analysis
2.6. Hematological Analysis
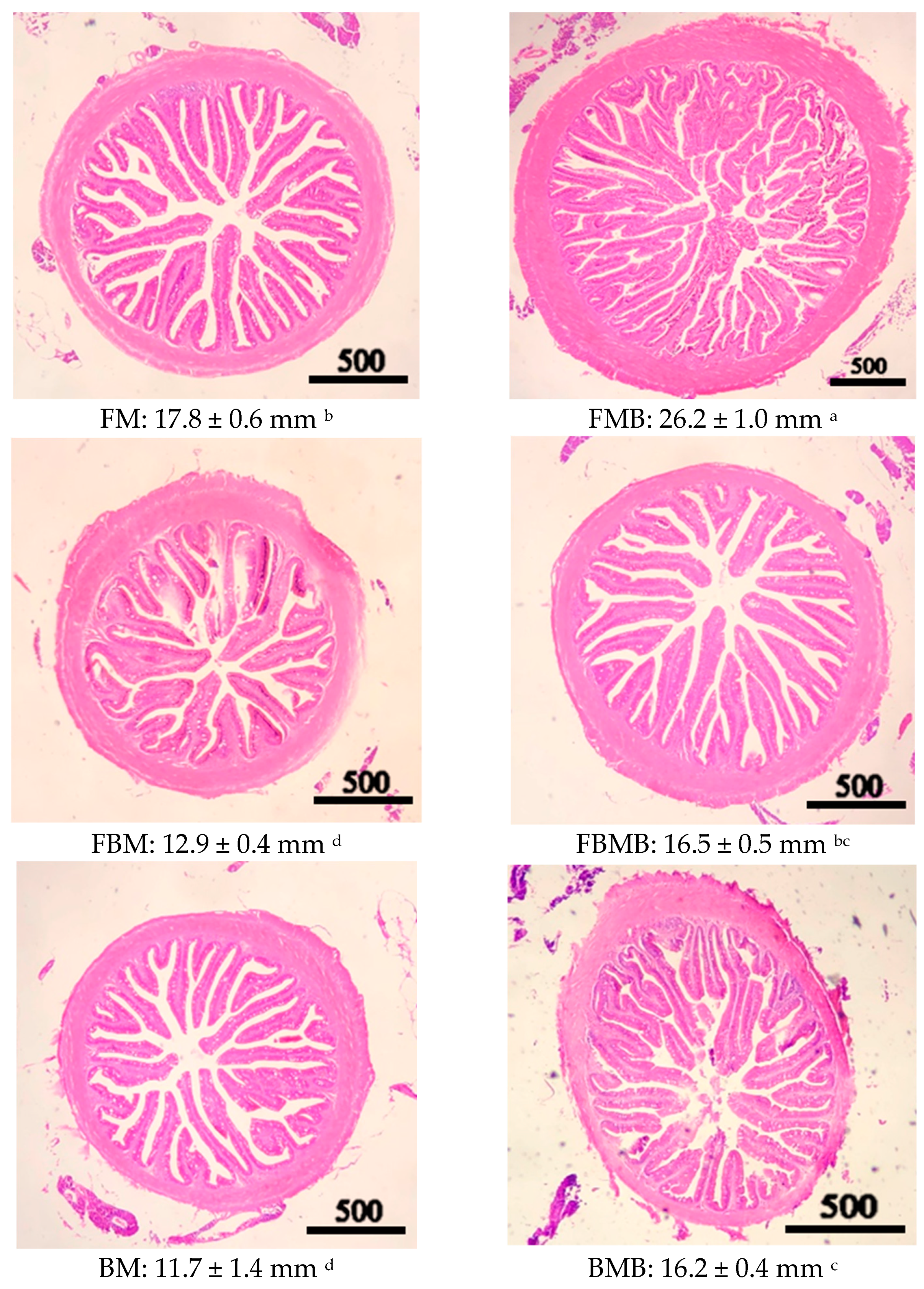
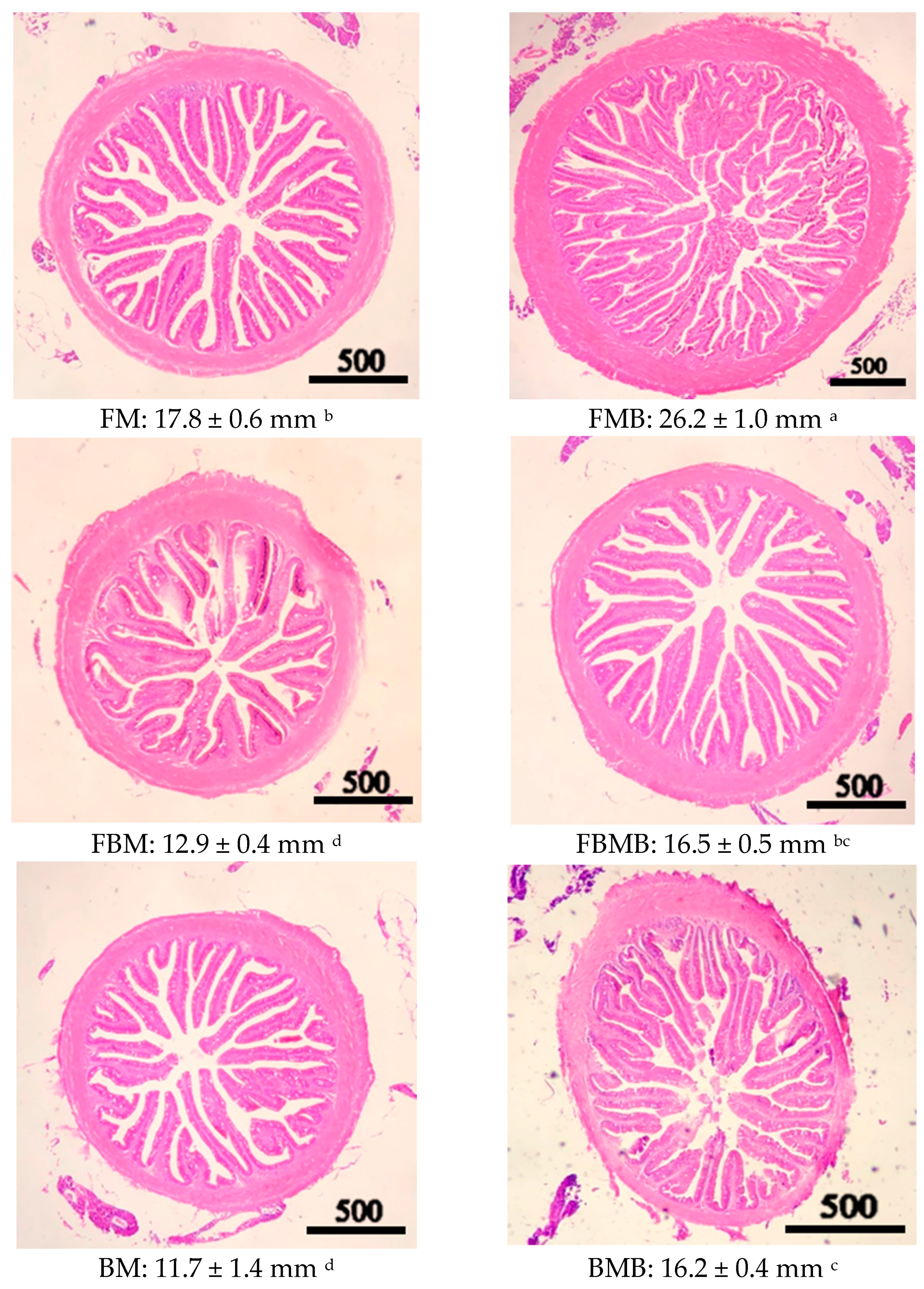
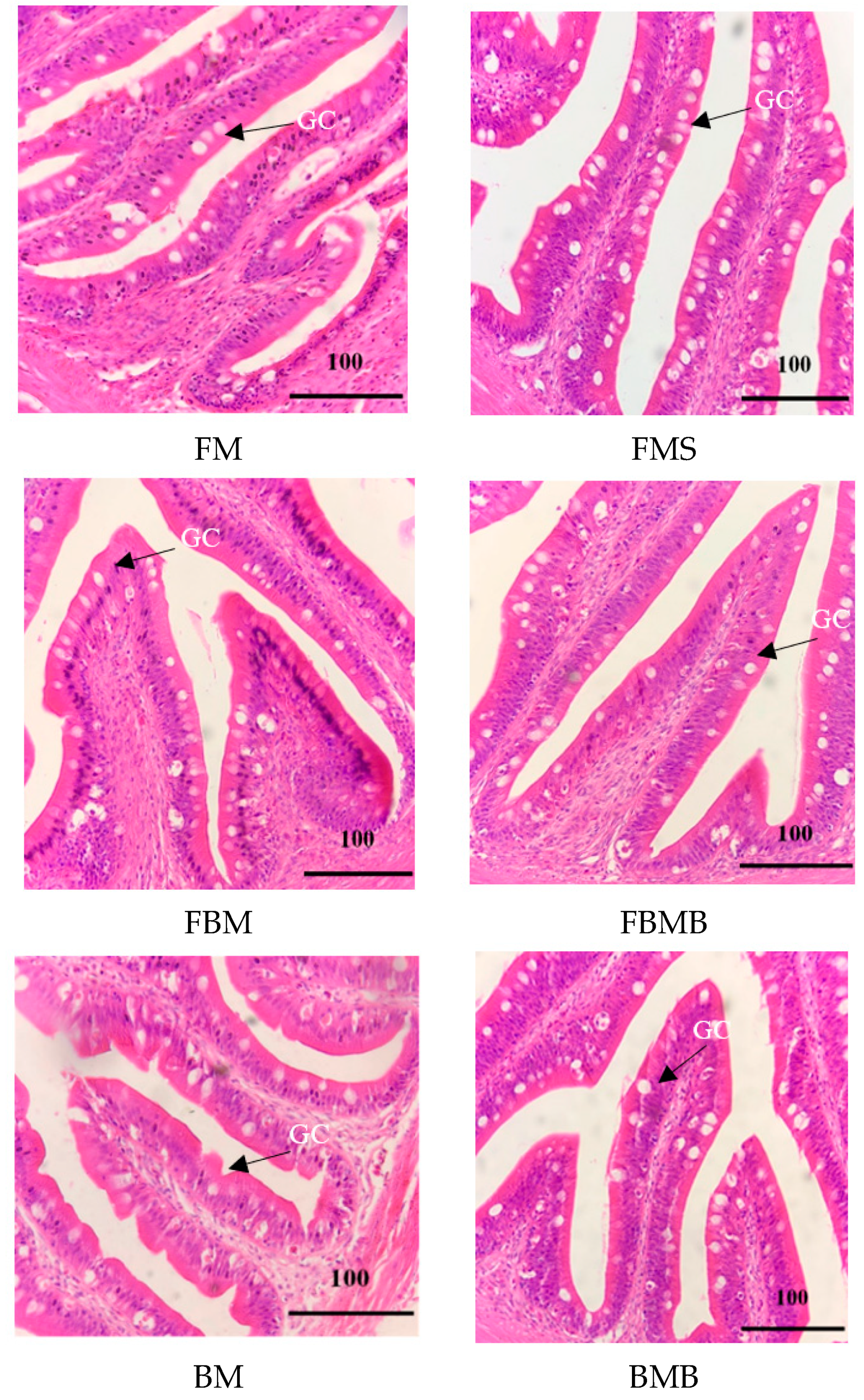
2.7. Intestinal Tract Section
2.8. Fatty Acid Composition (% Total Fatty Acids) Analysis
2.9. Amino Acid Analysis
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Blood Chemistry Test
3.3. Body Composition Analysis
3.4. Analysis of Antioxidant Enzyme Activity
3.5. Intestinal Tract Section
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tacon, A.G.J.; Silva, S.S.D. Feed preparation and feed management strategies within semi-intensive fish farming systems in the tropics. Aquaculture 1997, 151, 379–404. [Google Scholar] [CrossRef]
- Péron, G.; Mittaine, J.F.; Gallic, B.L. Where do fishmeal and fish oil products come from? An analysis of the conversion ratios in the global fishmeal industry. Mar. Policy 2010, 34, 815–820. [Google Scholar] [CrossRef]
- Zhou, F.; Song, W.; Shao, Q.; Peng, X.; Xiao, J.; Hua, Y.; Ng, W.K. Partial replacement of fish meal by fermented soybean meal. in diets for black sea bream, Acanthopagrus schlegelii, juveniles. J. World Aquacult. Soc. 2011, 42, 184–197. [Google Scholar] [CrossRef]
- Kim, S.S.; Galaz, G.B.; Pham, M.A.; Jang, J.W.; Oh, D.H.; Yeo, I.K.; Lee, K.J. Effects of dietary supplementation of a Meju, Fermented soybean meal, and aspergillus oryzae for juvenile Parrot fish (Oplegnathus fasciatus). Asian-Aust. J. Anim. Sci. 2009, 22, 849–856. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Parolini, M.; Ganzaroli, A.; Bacenetti, J. Earthworm as an alternative protein source in poultry and fish farming: Current applications and future perspectives. Sci. Total Environ. 2020, 734, 139460. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2023. [Google Scholar]
- Van Huis, A.; Tomberlin, J.K. Insects as Food and Feed: From Production to Consumption, 1st ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; pp. 73–96. [Google Scholar]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 53, 53. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Dosdat, A. Environmental impact of aquaculture in the Mediterranean: Nutritional and feeding aspects. Zaragoza: CIHEAM. Cah. Options Méditerranéennes 2001, 55, 23–36. [Google Scholar]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M.R. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hemato-biochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar] [CrossRef]
- Xiao, X.; Jin, P.; Zheng, L.; Cai, M.; Yu, Z.; Yu, J.; Zhang, J. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fishmeal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aquac. Res. 2018, 49, 1569–1577. [Google Scholar] [CrossRef]
- Huyben, D.; Vidaković, A.; Hallgren, S.W.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout fed larval and prepupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Fisher, H.J.; Collins, S.A.; Hanson, C.; Mason, B.; Colombo, S.M.; Anderson, D.M. Black soldier fly larvae meal as a protein source in low fish meal diets for Atlantic salmon (Salmo salar). Aquaculture 2020, 521, 734978. [Google Scholar] [CrossRef]
- Devic, E.; Leschen, W.; Murray, F.; Little, D.C. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing black soldier fly (Hermetia illucens) larvae meal. Aquac. Nutr. 2018, 24, 416–423. [Google Scholar] [CrossRef]
- Foysal, M.J.; Fotedar, R.; Tay, C.Y.; Gupta, S.K. 2019. Dietary supplementation of black soldier fly (Hermetica illucens) meal modulates gut microbiota, innate immune response and health status of marron (Cherax cainii, Austin 2002) fed poultry-by-product and fishmeal based diets. PeerJ 2019, 7, e6891. [Google Scholar] [CrossRef]
- Gebremichael, A.; Kucska, B.; Ardó, L.; Biró, J.; Berki, M.; Lengyel-Kónya, É.; Tömösközi-Farkas, R.; Egessa, R.; Müller, T.; Gyalog, G.; et al. Physiological Response of Grower African Catfish to Dietary Black Soldier Fly and Mealworm Meal. Animals 2023, 13, 968. [Google Scholar] [CrossRef] [PubMed]
- Gorka, P.; Kowalski, Z.M.; Pietrzak, P.; Kotunia, A.; Jagusiak, W.; Holst, J.J. Effect of method of delivery of sodium butyrate on rumen development in newborn calves. J. Diary Sci. 2011, 94, 5578–5588. [Google Scholar] [CrossRef]
- Berni Canani, R.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal disease. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Astbury, S.M.; Corfe, B.M. Uptake and metabolism of the short-chain fatty acid butyrate, a critical review of the literature. Curr. Drug Metab. 2012, 13, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.W.; Jiang, S.; Qian, D.W.; Duan, J.A. Modulation of microbially derived short-chain fatty acids on intestinal homeostasis, metabolism, and neuropsychiatric disorder. Appl. Microbiol. Biotechnol. 2020, 104, 589–601. [Google Scholar] [CrossRef]
- Mog, M.; Ngasotter, S.; Tesia, S.; Waikhom, D.; Panda, S.P.; Sharma, S.; Varshney, S. Problems of antibiotic resistance associated with oxytetracycline use in aquaculture: A review. J. Entomol. Zool. Stud. 2020, 8, 1075–1082. [Google Scholar]
- Manrique Vergara, D.; González Sánchez, M.E. Short chain fatty acids (butyric acid) and intestinal diseases. Nutr. Hosp. 2017, 34, 58–61. [Google Scholar] [CrossRef]
- Zarei, S.; Badzohreh, G.; Davoodi, R.; Bahabadi, M.N.; Salehi, F. Effects of dietary butyric acid glycerides on growth performance, haemato-immunological and antioxidant status of yellowfin seabream (Acanthopagrus latus) fingerlings. Aquac. Res. 2021, 52, 5840–5848. [Google Scholar] [CrossRef]
- Aalamifar, H.; Soltanian, S.; Vazirzadeh, A.; Akhlaghi, M.; Morshedi, V.; Gholamhosseini, A.; Mozanzadeh, M.T. Dietary butyric acid improved growth, digestive enzyme activities and humoral immune parameters in Barramundi (Lates calcarifer). Aquac. Nutr. 2020, 26, 156–164. [Google Scholar] [CrossRef]
- Sangari, M.; Sotoudeh, E.; Bagheri, D.; Morammazi, S.; Mozanzadeh, M.T. Growth, body composition, and hematology of yellowfin seabream (Acanthopagrus latus) given feeds supplemented with organic acid salts (sodium acetate and sodium propionate). Aquac. Int. 2020, 29, 261–273. [Google Scholar] [CrossRef]
- Sotoudeh, E.; Sangari, M.; Bagheri, D.; Morammazi, S.; Torfi Mozanzadeh, M. Dietary organic acid salts mitigate plant protein induced inflammatory response and improve humoral immunity, antioxidative status and digestive enzyme activities in yellowfin seabream, Acanthopagrus latus. Aquac. Nutr. 2020, 2, 1669–1680. [Google Scholar] [CrossRef]
- Chu, J.H.; Liu, K.J.; Wu, T.M. Effects of pitaya (Hylocereus polyrhizus) fermentation waste dietary supplement on growth performance, and anti-oxidation of Pinnate batfish, Platax pinnatus. Aquac. Res. 2021, 52, 6689–6698. [Google Scholar] [CrossRef]
- Chu, J.H.; Chen, S.M.; Huang, C.H. Growth, hematological parameters and tissue lipid peroxidation of soft-shelled turtles, Pelodiscus sinensis, fed diets supplemented with different levels of ferrous sulphate. Aquac. Nutr. 2009, 15, 54–59. [Google Scholar] [CrossRef]
- Han, X.; Kun, Y.E.; Wang, Z.; Chen, F.; Chai, Z.; Zhan, W.; Wang, Q. Effect of substitution of fish meal with defatted black soldier fly larvae meal on growth, body composition, serum biochemical parameters, and antioxidant capacity of juvenile large yellow croaker (Larimichthys crocea). J. Fish. Sci. China 2020, 27, 524–535. [Google Scholar]
- Hu, J.R.; Wang, G.X.; Huang, Y.H. Effects of substitution of fish meal with black soldier fly (Hermetia illucens) larvae meal, in yellow catfish (Pelteobagrus fulvidraco) diets. Isr. J. Aquac. Bamidgeh 2017, 69, 1382–1391. [Google Scholar]
- Wang, G.X.; Peng, H.U., Jr. Evaluation of defatted Hermetia illucens larvae meal for Litopenaeus vannamei: Effects on growth performance, nutrition retention, antioxidant and immune response, digestive enzyme activity and hepatic morphology. Aquac. Nutr. 2021, 27, 986–997. [Google Scholar] [CrossRef]
- Thrastardottir, R.; Olafsdottir, H.T.; Thorarinsdottir, R.I. Yellow Mealworm and Black Soldier Fly Larvae for Feed and Food Production in Europe, with Emphasis on Iceland. Foods 2021, 10, 2744. [Google Scholar] [CrossRef]
- Dong, B.; Yi, Y.H.; Liang, L.F. High throughput identification of antimicrobial peptides from fish gastrointestinal microbiota. Toxins 2017, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Robles, R.; Lozano, A.B.; Sevilla, A.; Máarquez, L.; Nuez-Ortin, W.; Moyano, F.J. Effect of partially protected butyrate used as feed additive on growth and intestinal metabolism in sea bream (Sparus aurata). Fish Physiol. Biochem. 2013, 39, 1567–1580. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Finke, M.D. Estimate of Chitin in Raw Whole Insect. Zoo Biol. 2007, 26, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Tawwab, M.; Shukry, M.; Farrag, F.A.; El-Shafai, N.M.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Dietary sodium butyrate nanoparticles enhanced growth, digestive enzyme activities, intestinal histomorphometry, and transcription of growth-related genes in Nile tilapia juveniles. Aquaculture 2021, 536, 736467. [Google Scholar] [CrossRef]
- Abdel-Mohsen, H.H.; Wassef, E.A.; El-Bermawy, N.M.; Abdel-Meguid, N.E.; Saleh, N.E.; Barakat, K.M.; Shaltout, O.E. Advantageous effects of dietary butyrate on growth, immunity response, intestinal microbiota and histomorphology of European Seabass (Dicentrarchus labrax) fry. Egypt. J. Aquat. Biol. Fish. 2018, 22, 93–110. [Google Scholar] [CrossRef]
- Liu, M.; Guo, W.; Wu, F.; Qu, Q.C.; Tan, Q.S.; Gong, W.B. Dietary supplementation of sodium butyrate may benefit growth performance and intestinal function in juvenile grass carp (Ctenopharyngodon idellus). Aquac. Res. 2016, 47, 4102–4111. [Google Scholar] [CrossRef]
- Zhou, J.S.; Guo, P.; Yu, H.B.; Ji, H.; Lai, Z.W.; Chen, Y.A. Growth performance, lipid metabolism, and health status of grass carp (Ctenopharyngodon idella) fed three different forms of sodium butyrate. Fish Physiol. Biochem. 2019, 45, 287–298. [Google Scholar] [CrossRef]
- Abu Elala, N.M.; Ragaa, N.M. Eubiotic effect of a dietary acidifier (potassium diformate) on the health status of cultured Oreochromis niloticus. J. Adv. Res. 2015, 6, 621–629. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, C.; Cao, K.; Li, Z.; Zhao, Z.; Li, X.; Leng, X. Dietary Sodium Butyrate Changed Intestinal Histology and Microbiota of Rainbow Trout (Oncorhynchus mykiss), but Did Not Promote Growth and Nutrient Utilization. Aquac. Nutr. 2023, 2023, 3706109. [Google Scholar]
- Vielma, J.; Lall, S. Dietary formic acid enhances apparent digestibility of minerals in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Nutr. 1997, 3, 265–268. [Google Scholar] [CrossRef]
- Bröer, S. Amino Acid Transport Across Mammalian Intestinaland Renal Epithelia. Physiol. Rev. 2008, 88, 249–286. [Google Scholar]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 262–1267. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Abdel-Tawwab, M.; Khafagac, A.F.; Dawood, M.A.O. Dietary oregano essential oil improved the growth performance via enhancing the intestinal morphometry and hepato-renal functions of common carp (Cyprinus carpio L.) fingerlings. Aquaculture 2020, 526, 735432. [Google Scholar] [CrossRef]
- Caballero, M.J.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Montero, D.; Socorro, J.; Fernández, A.J.; Rosenlund, G. Morphological aspects of intestinal cells from gilthead seabream (Sparus aurata) fed diets containing different lipid sources. Aquaculture 2003, 225, 325–340. [Google Scholar]
- Ostaszewska, T.; Dabrowski, K.; Kamaszewski, M.; Grochowski, P.; Verri, T.; Rzepkowska, M.; Wolnicki, J. The effect of plant protein-based diet supplemented with dipeptide or free amino acids on digestive tract morphology and PepT1 and PepT2 expressions in common carp (Cyprinus carpio L.). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 157, 158–169. [Google Scholar]
- Buscaino, G.; Filiciotto, F.; Buffa, G.; Bellante, A.; Di Stefano, V.; Assenza, A.; Fazio, F.; Caola, G.; Mazzola, S. Impact of an acoustic stimulus on the motility and blood parameters of European sea bass (Dicentrarchus labrax L.) and gilthead sea bream (Sparus aurata L.). Mar. Environ. Res. 2010, 69, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Fazio, F.; Marafioti, S.; Arfuso, F.; Piccione, G.; Faggio, C. Influence of different salinity on haematological and biochemical parameters of the widely cultured mullet, Mugil cephalus. Mar. Freshw. Behav. Physiol. 2013, 46, 211–218. [Google Scholar] [CrossRef]
- Caldwell, S.H.; Oelsner, D.H.; Iezzoni, J.C.; Hespenheide, E.E.; Battle, E.H.; Driscoll, C.J. Cryptogenic cirrhosis: Clinical characterization and risk factors for underlying disease. Hepatology 1999, 29, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.H.; Ye, J.Y.; Zhang, Y.X. The influence of maggot meal and L-carnitine on growth, immunity, antioxidant indices and disease resistance of black carp (Mylopharyngodon piceus). J. Chin. Cereals Oil Assoc. 2013, 28, 80–86. [Google Scholar]
- Wen, Y.H.; Huang, Y.H.; Wang, G.X. Effect of replacement of fish meal with maggot meal on antioxidant indexes, digestive enzyme activities, foregut and hepatopancreas histological structure of Pelteobagrus falvidraco. Feed. Ind. 2015, 36, 29–35. [Google Scholar]
- Abd El-Naby, A.S.; Khattaby, A.E.-R.A.; Samir, F.; Awad, S.M.M.; Abdel-Tawwab, M. Stimulatory effect of dietary butyrate on growth, immune response, and resistance of Nile tilapia, Oreochromis niloticus against Aeromonas hydrophila infection. Anim. Feed Sci. Technol. 2019, 254, 114212. [Google Scholar] [CrossRef]
- Mirghaed, A.T.; Yarahmadi, P.; Soltani, M.; Paknejad, H.; Hoseini, S.M. Dietary sodium butyrate (Butirex® C4) supplementation modulates intestinal transcriptomic responses and augments disease resistance of rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2019, 92, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar]
- Slaninova, A.; Smutna, M.; Modar, H.; Svobodova, Z. A review: Oxidative stress in fish induced by pesticides. Neuroendocrinol. Lett. 2009, 30, 2–12. [Google Scholar]
- Birnie-Gauvin, K.; Costantini, D.; Cooke, S.J.; Willmore, W.G. A comparative and evolutionary approach to oxidative stress in fish: A review. Fish Fish. 2017, 18, 928–942. [Google Scholar] [CrossRef]
- Ibrahim, R.E.; El-Houseiny, W.; Behairy, A.; Abo-Elmaaty, A.; Al-Sagheer, A.A. The palliative role of Eruca sativa leaves dietary supplementation against oxidative stress, immunosuppression, and growth retardation in temperature-stressed Oreochromis niloticus. J. Therm. Biol. 2019, 84, 26–35. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Caipang, C.M. Short-chain fatty acids as feed supplements for sustainable aquaculture: An updated view. Aquac. Res. 2017, 48, 1380–1391. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Eweedah, N.M.; Elbialy, Z.I.; Abdelhamid, A.I. Dietary sodium butyrate ameliorated the blood stress biomarkers, heat shock proteins, and immune response of Nile tilapia (Oreochromis niloticus) exposed to heat stress. J. Therm. Biol. 2020, 88, 102500. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Wang, Q.; Guo, X.; Pan, X.; Li, X. Effects of dietary sodium butyrate on growth performance, antioxidant capacity, intestinal histomorphology and immune response in juvenile Pengze crucian carp (Carassius auratus Pengze). Aquac. Rep. 2021, 21, 100828. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Treatments | FM | FMB | FBM | FBMB | BM | BMB |
|---|---|---|---|---|---|---|
| Fish meal | 50 | 50 | 25 | 25 | 0 | 0 |
| BSF 5 meal | 0 | 0 | 26.4 | 26.4 | 52.8 | 52.8 |
| Oil 4 | 6.06 | 6.06 | 3.92 | 3.92 | 1.78 | 1.78 |
| Yeast | 1 | 1 | 1 | 1 | 1 | 1 |
| α-starch | 10 | 10 | 10 | 10 | 10 | 10 |
| Vitamin mix 1 | 2 | 2 | 2 | 2 | 2 | 2 |
| Mineral mix 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| α-Cellulose | 28.94 | 27.94 | 29.68 | 28.68 | 30.42 | 29.42 |
| Butyric acid | 0 | 1 | 0 | 1 | 0 | 1 |
| Analyzed composition (as fed) 3 | ||||||
| Moisture | 10.72 | 9.58 | 10.19 | 8.65 | 9.18 | 9.77 |
| Crude protein | 31.58 | 31.42 | 31.38 | 31.29 | 31.66 | 31.13 |
| Crude lipid | 8.51 | 8.80 | 8.80 | 8.02 | 8.07 | 8.29 |
| Ash | 15.77 | 16.53 | 11.37 | 11.76 | 6.91 | 7.45 |
| Crude fiber | 26.42 | 25.76 | 28.22 | 28.65 | 29.52 | 28.64 |
| Fatty Acids | FM/FMB | FBM/FBMB | BM/BMB |
|---|---|---|---|
| 12:0 | 3.96 | 6.67 | 9.38 |
| 14:0 | 5.56 | 2.87 | 2.18 |
| 14:1 | 0.01 | 0.01 | 0.01 |
| 16:0 | 22.66 | 17.71 | 12.75 |
| 16:1 | 0.46 | 0.40 | 0.34 |
| 18:0 | 5.72 | 4.22 | 2.73 |
| 18:1 | 0.23 | 7.96 | 15.69 |
| 18:2 n-6 | 21.57 | 28.20 | 34.82 |
| 18:3 n-3 | 2.31 | 3.12 | 2.93 |
| 20:0 | 0.17 | 0.18 | 0.19 |
| 20:1 | 0.15 | 0.19 | 0.23 |
| 20:2 n-6 | 0.39 | 0.31 | 0.23 |
| 20:3 n-6 | 1.22 | 0.94 | 0.66 |
| 20:3 n-3 | 1.56 | 1.93 | 2.30 |
| 20:4 n-6 | 14.46 | 11.35 | 6.25 |
| 20:5 n-3 | 0.74 | 0.43 | 0.11 |
| 22:0 | 0.23 | 2.19 | 3.14 |
| 22:1 | 0.90 | 0.73 | 0.56 |
| 22:5 n-3 | 5.20 | 3.10 | 1.01 |
| 22:6 n-3 | 12.50 | 7.50 | 4.50 |
| SFAs | 38.29 | 33.84 | 30.37 |
| MUFAs | 1.76 | 9.29 | 16.83 |
| PUFAs | 27.05 | 34.5 | 40.94 |
| HUFAs | 32.90 | 22.38 | 11.87 |
| Amino acids | |||
| Arginine | 2.20 | 1.56 | 0.99 |
| Histidine | 0.77 | 0.66 | 0.54 |
| Isoleucine | 1.58 | 1.09 | 0.60 |
| Leucine | 2.44 | 1.70 | 0.95 |
| Lysine | 2.67 | 1.67 | 0.66 |
| Methionine | 0.86 | 0.95 | 1.04 |
| Phenylalanine | 1.34 | 0.98 | 0.61 |
| Threonine | 1.37 | 0.94 | 0.50 |
| Valine | 1.70 | 1.25 | 0.79 |
| Tryptophan | 0.34 | 0.39 | 0.43 |
| Sum of EAA | 15.27 | 11.19 | 7.11 |
| Treatments | |||||||
|---|---|---|---|---|---|---|---|
| Initial Weight | Final Weight | WG | FCR | PER | FI | Survival | |
| FM | 7.3 ± 0.1 | 52.3 ± 2.8 b | 616.4 ± 47.9 b | 1.7 ± 0.1 cd | 1.8 ± 0.1 b | 77.7 ± 5.6 ab | 100 |
| FMB | 7.3 ± 0.1 | 66.2 ± 5.2 a | 806.5 ± 60.9 a | 1.3 ± 0.2 d | 2.5 ± 0.4 a | 77.5 ± 17.7 ab | 100 |
| FBM | 7.3 ± 0.2 | 33.6 ± 3.0 c | 359.9 ± 42.3 c | 2.3 ± 0.3 ab | 1.3 ± 0.2 c | 62.4 ± 3.2 bc | 100 |
| FBMB | 7.3 ± 0.1 | 49.9 ± 5.1 b | 580.1 ± 64.2 b | 2.0 ± 0.3 bc | 1.7 ± 0.2 bc | 83.0 ± 5.2 a | 100 |
| BM | 7.3 ± 0.1 | 32.1 ± 2.7 c | 339.3 ± 32.1 c | 2.7 ± 0.7 a | 1.2 ± 0.3 c | 66.4 ± 9.7 abc | 100 |
| BMB | 7.3 ± 0.1 | 33.7 ± 2.7 c | 359.2 ± 37.0 c | 2.2 ± 0.2 abc | 1.5 ± 0.1 bc | 58.6 ± 7.5 c | 100 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| RBC | HGB | Hct | MCV | MCH | MCHC | |
| FM | 2.2 ± 0.2 b | 7.7 ± 0.2 c | 29.2 ± 0.9 b | 36.1 ± 1. 7 a | 136.9 ± 11.7 a | 26.4 ± 1.1 c |
| FMB | 3.1 ± 0.2 a | 9.4 ± 0.2 a | 32.5 ± 1.2 a | 30.3 ± 3.0 b | 105.3 ± 11.6 b | 28.8 ± 1.2 ab |
| FBM | 2.2 ± 0.1 b | 7.5 ± 0.4 c | 25.0 ± 1.5 c | 34.8 ± 3.4 a | 116.0 ± 7.5 b | 29.9 ± 1.7 a |
| FBMB | 3.1 ± 0.1 a | 8.6 ± 0.2 b | 31.2 ± 1.0 ab | 27.9 ± 09 b | 101.2 ± 5.2 b | 27.6 ± 0.9 abc |
| BM | 2.1 ± 0.1 b | 7.2 ± 0.2 c | 24.6 ± 1.2 c | 34.3 ± 0.8 a | 116.5 ± 4.0 b | 29.4 ± 0.7 a |
| BMB | 3.0 ± 0.1 a | 8.3 ± 0.3 b | 30.8 ± 0.6 ab | 28.1 ± 1.5 b | 104.3 ± 5.1 b | 27.0 ± 1.5 bc |
| Treatments | ||||
|---|---|---|---|---|
| Moisture | Crude Protein | Crude Lipids | Ash | |
| FM | 77.7 ± 0.1 a | 86.3 ± 0.2 b | 15.7 ± 0.1 a | 4.4 ± 0.2 |
| FMB | 77.7 ± 0.1 ab | 88.4 ± 0.2 a | 14.2 ± 0.1 b | 4.4 ± 0.4 |
| FBM | 778 ± 0.1 a | 85.0 ± 0.5 c | 14.1 ± 0.7 b | 4.4 ± 0.5 |
| FBMB | 77.7 ± 0.1 ab | 86.3 ± 0.2 b | 13.6 ± 0.1 bc | 4.4 ± 0.1 |
| BM | 77.7 ± 0.1 ab | 84.1 ± 0.4 d | 13.5 ± 0.2 c | 4.5 ± 0.1 |
| BMB | 77.6 ± 0.1 b | 85.5 ± 0.1 c | 13.2 ± 0.1 c | 4.4 ± 0.2 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| SOD | CAT | GPX | ALT | AST | MDA | |
| FM | 10.4 ± 0.5 c | 9.3 ± 0.6 c | 11.0 ± 0.5 c | 19.8 ± 0.3 b | 20.8 ± 0.6 b | 1.2 ± 0.1 d |
| FMB | 14.5 ± 0.7 a | 13.8 ± 0.6 a | 16.2 ± 0.3 a | 27.2 ± 0.8 a | 28.5 ± 0.1 a | 1.0 ± 0.1 e |
| FBM | 9.4 ± 1.0 cd | 8.4 ± 1.2 cd | 10.0 ± 0.6 d | 19.8 ± 0.3 b | 20.7 ± 1.1 b | 1.3 ± 0.1 c |
| FBMB | 11.9 ± 0.7 b | 11.1 ± 0.6 b | 14.2 ± 0.3 b | 27.3 ± 1.1 a | 28.5 ± 0.1 a | 1.3 ± 0.2 c |
| BM | 8.2 ± 0.7 d | 7.5 ± 0.5 d | 8.8 ± 0.2 e | 19.9 ± 0.3 b | 20.7 ± 1.8 b | 1.6 ± 0.1 a |
| BMB | 10.1 ± 0.3 c | 9.0 ± 0.9 c | 11.3 ± 0.8 c | 26.8 ± 2.4 a | 28.6 ± 1.2 a | 1.5 ± 0.1 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, J.-H.; Weng, T.-S.; Huang, T.-W. The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum barcoo Fingerlings. Fishes 2023, 8, 437. https://doi.org/10.3390/fishes8090437
Chu J-H, Weng T-S, Huang T-W. The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum barcoo Fingerlings. Fishes. 2023; 8(9):437. https://doi.org/10.3390/fishes8090437
Chicago/Turabian StyleChu, Jen-Hong, Tsung-Sheng Weng, and Tzu-Wei Huang. 2023. "The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum barcoo Fingerlings" Fishes 8, no. 9: 437. https://doi.org/10.3390/fishes8090437
APA StyleChu, J.-H., Weng, T.-S., & Huang, T.-W. (2023). The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum barcoo Fingerlings. Fishes, 8(9), 437. https://doi.org/10.3390/fishes8090437