
Exploring Biofouling Control by the California Sea Cucumber (Apostichopus californicus) in Integrated Multi-Trophic Aquaculture (IMTA) with Organic Chinook Salmon (Oncorhynchus tshawytscha)

Abstract
:1. Introduction
2. Materials and Methods
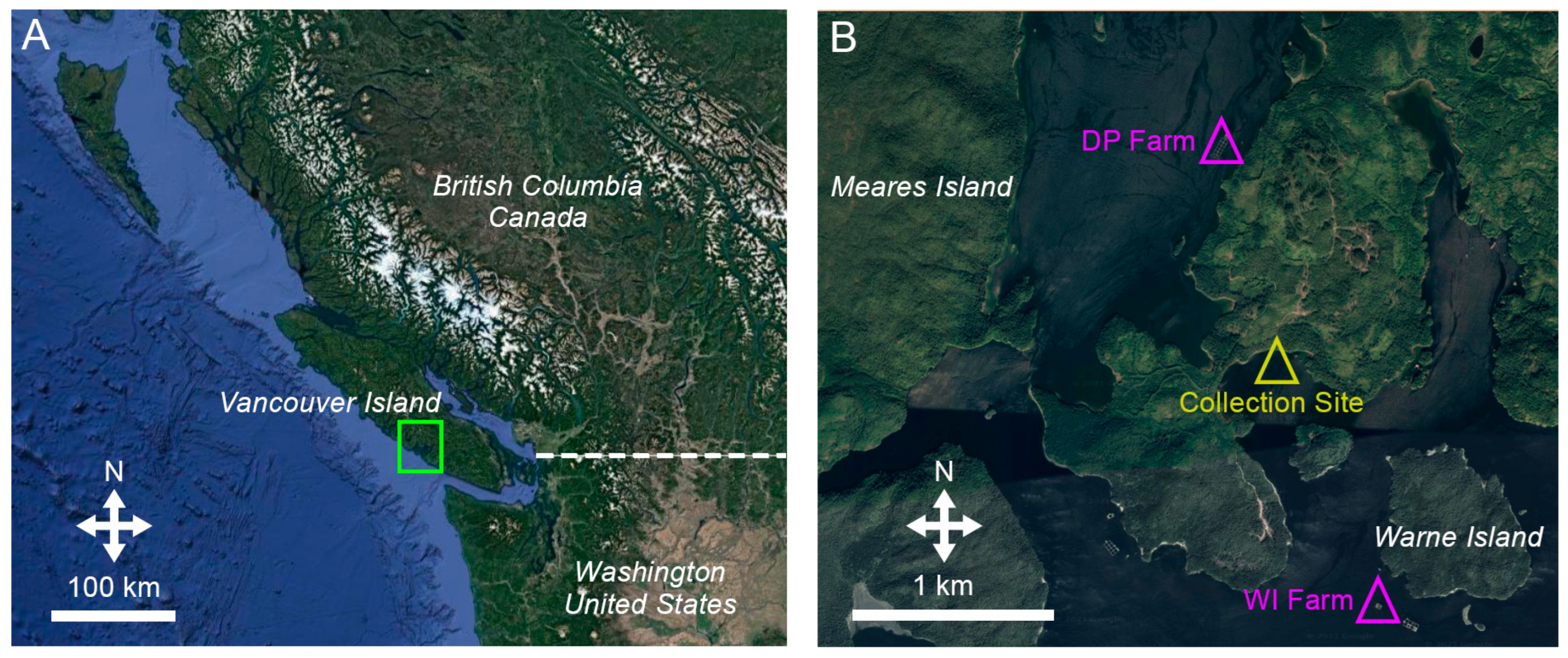
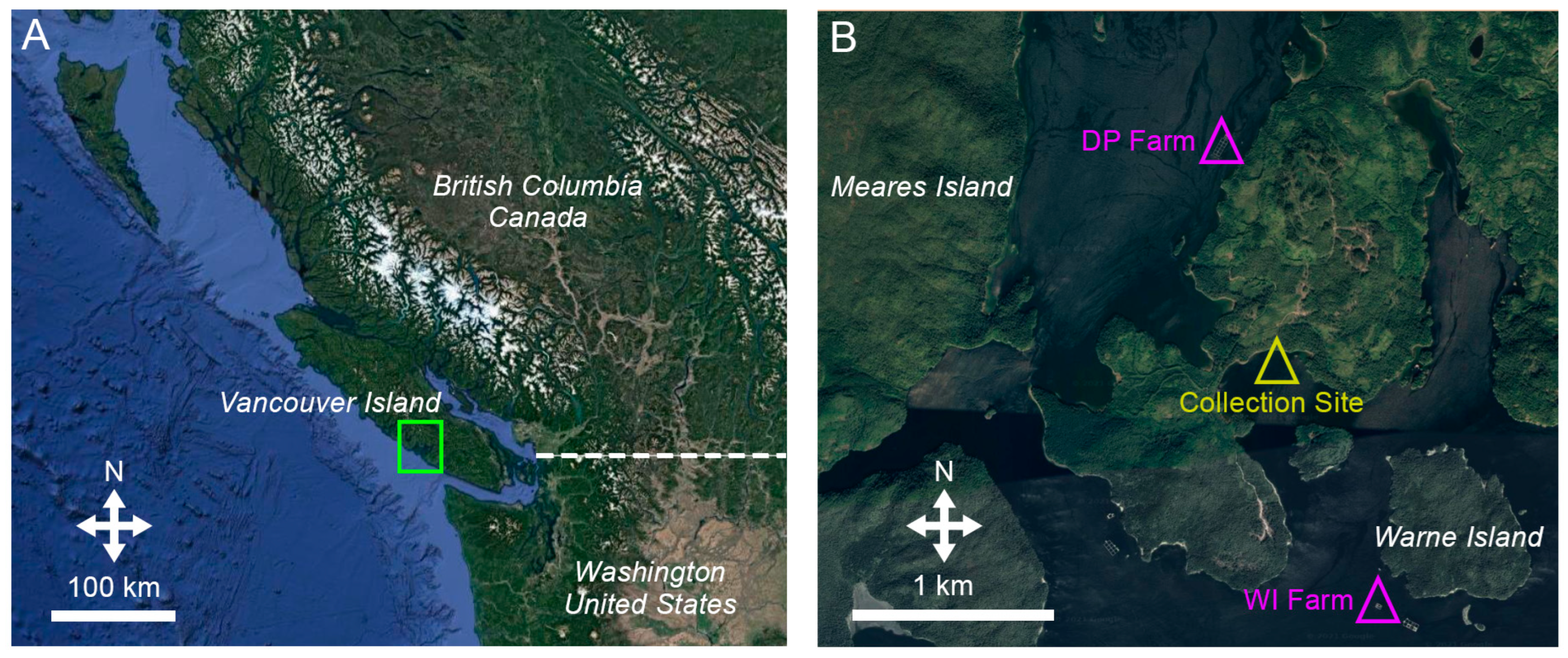
2.1. Study Area, Farm Parameters, and Source of Sea Cucumbers
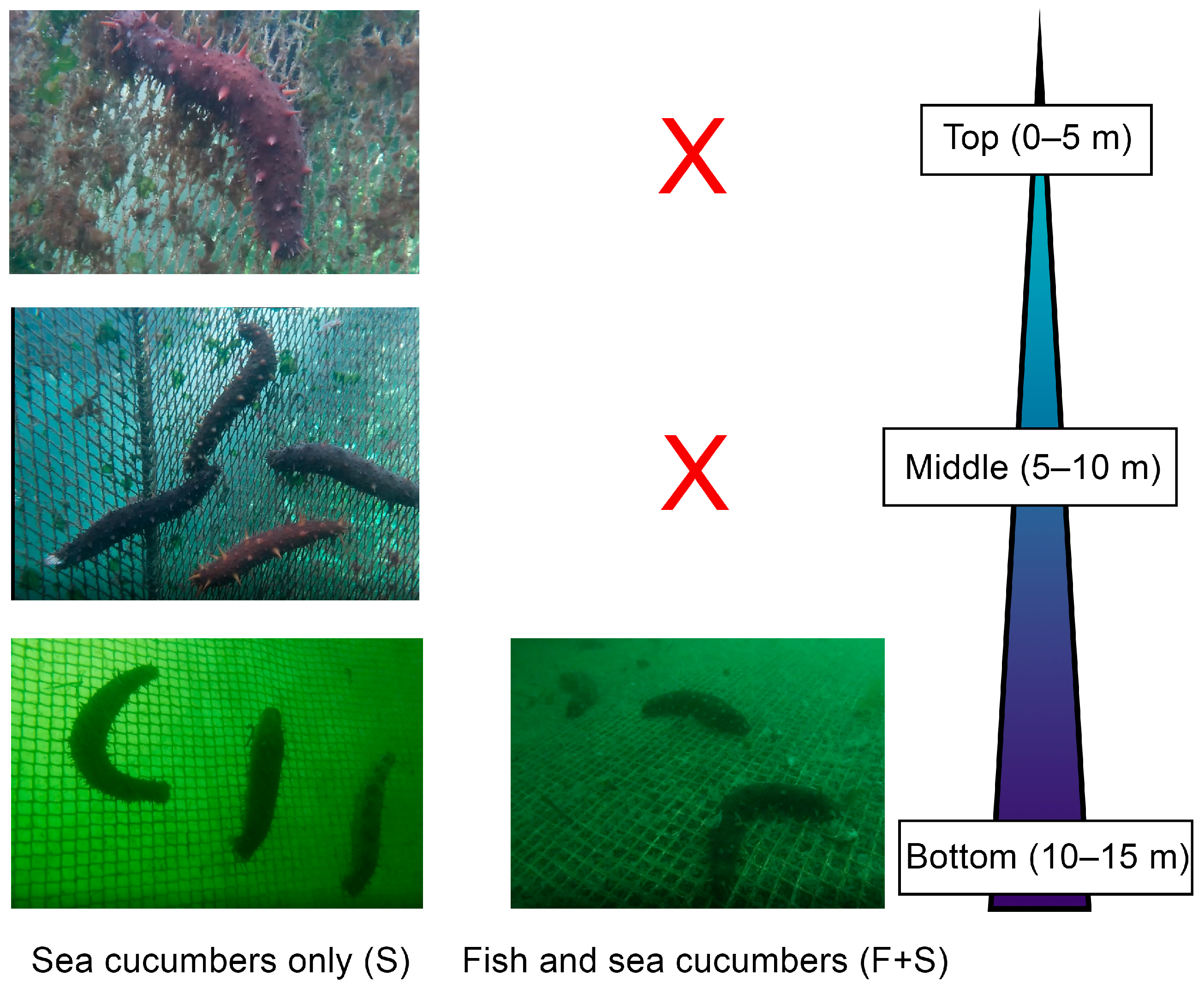
2.2. Experimental Design
2.3. Analysis of Net Percent Cleanliness
2.4. Stable Isotopes
2.5. Statistical Analysis
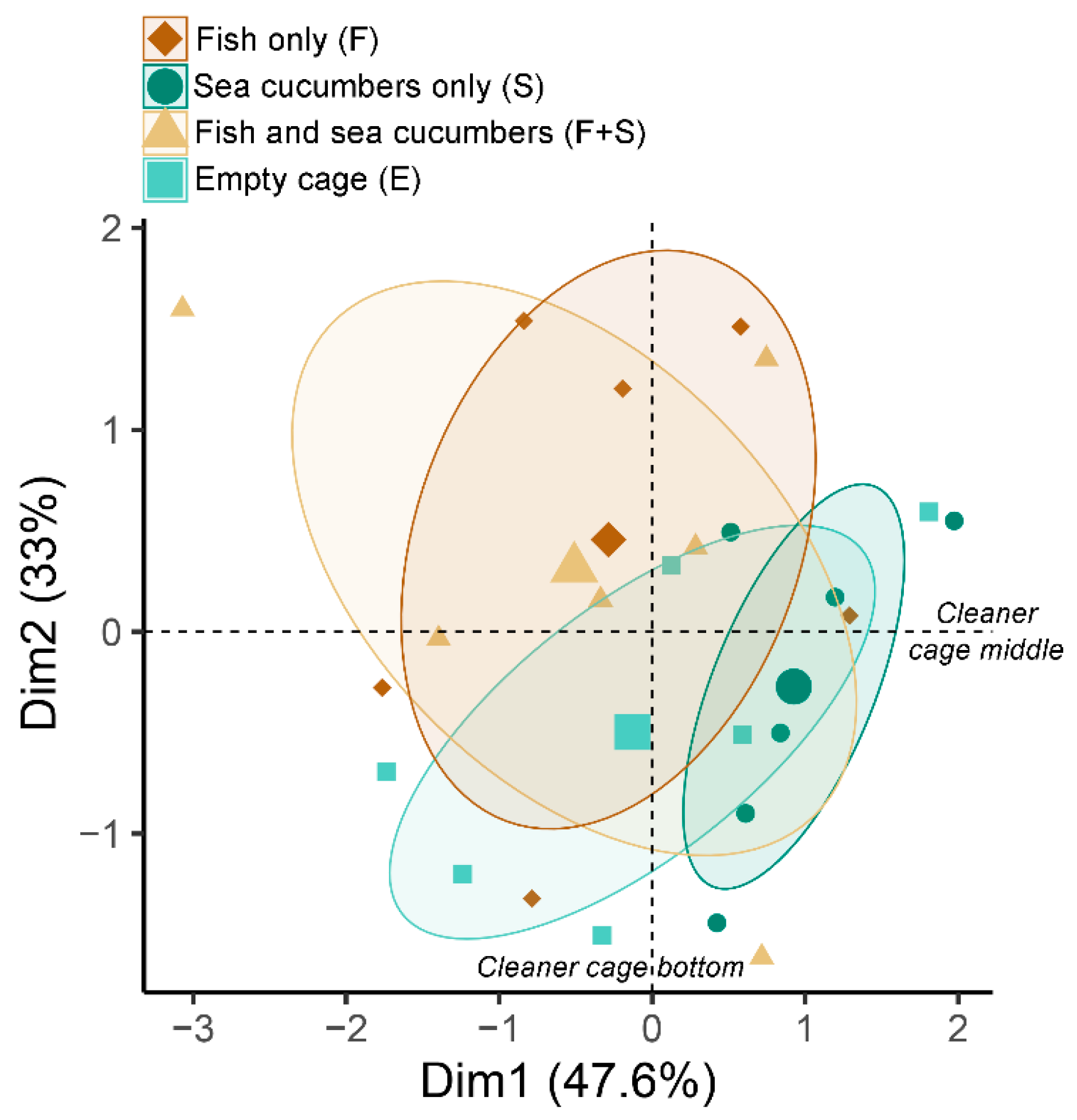
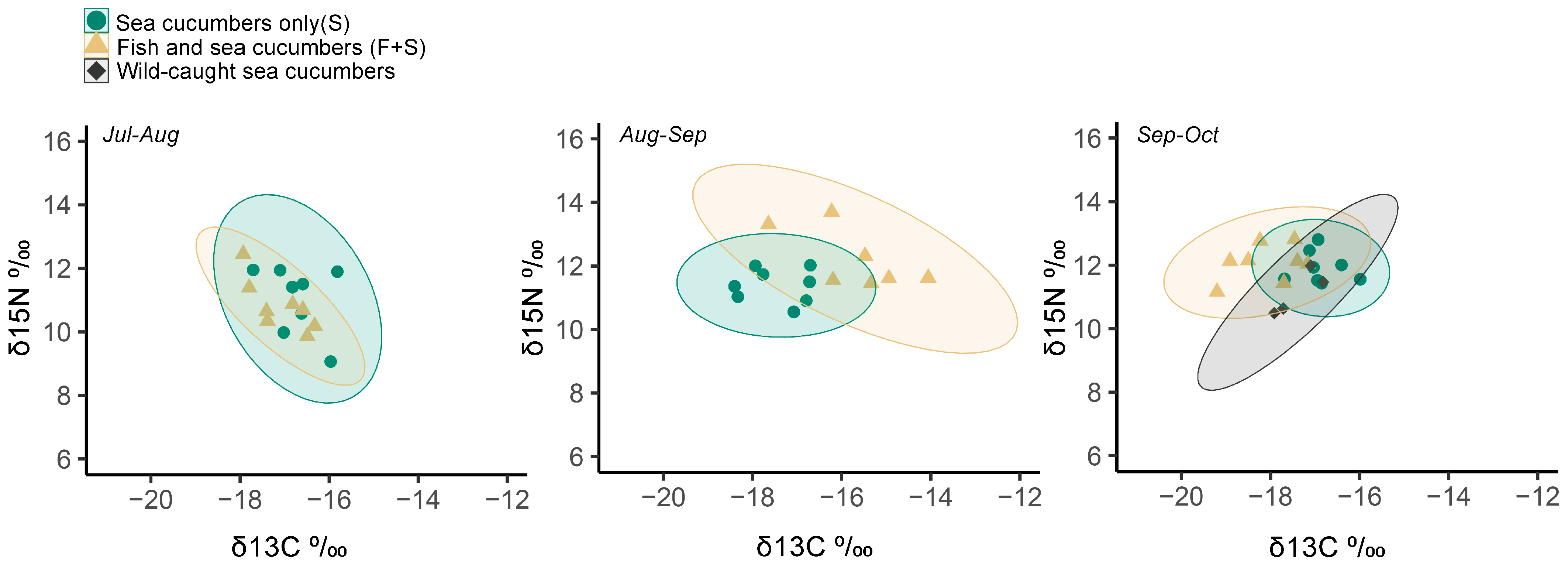
3. Results
3.1. Net Percent Cleanliness
3.2. Stable Isotopes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloecher, N.; Floerl, O. Towards Cost-Effective Biofouling Management in Salmon Aquaculture: A Strategic Outlook. Rev. Aquac. 2021, 13, 783–795. [Google Scholar] [CrossRef]
- Cornejo, P.; Guerrero, N.M.; Montes, R.M.; Quiñones, R.A.; Sepúlveda, H.H. Hydrodynamic Effect of Biofouling in Fish Cage Aquaculture Netting. Aquaculture 2020, 526, 735367. [Google Scholar] [CrossRef]
- Chen, Q.-P.; Bi, C.-W.; Zhang, Z.-X.; Zhao, Y.-P. Hydrodynamic Effect of Different Biofouling Types on Aquaculture Netting. Ocean. Eng. 2023, 279, 114430. [Google Scholar] [CrossRef]
- Yu, S.; Qin, H.; Li, P.; Gong, F. Drag Force Coefficient and Flow Field Variations of Net with Different Levels of Biological Fouling under Large-Eddy Simulation. J. Mar. Sci. Technol. 2023, 28, 506–523. [Google Scholar] [CrossRef]
- Floerl, O.; Sunde, L.M.; Bloecher, N. Potential Environmental Risks Associated with Biofouling Management in Salmon Aquaculture. Aquac. Environ. Interact. 2016, 8, 407–417. [Google Scholar] [CrossRef]
- Åtland, Å.; Dale, T.; Bloecher, N. Environmental Considerations in Aquaculture Health Management. In Aquaculture Health Management; Elsevier: Amsterdam, The Netherlands, 2020; pp. 235–280. [Google Scholar]
- Østevik, L.; Stormoen, M.; Nødtvedt, A.; Alarcón, M.; Lie, K.-I.; Skagøy, A.; Rodger, H. Assessment of Acute Effects of in situ Net Cleaning on Gill Health of Farmed Atlantic Salmon (Salmo salar). Aquaculture 2021, 545, 737203. [Google Scholar] [CrossRef]
- Fletcher, L.M.; Davidson, I.C.; Bucknall, B.G.; Atalah, J. Salmon Farm Biofouling and Potential Health Impacts to Fish from Stinging Cnidarians. Aquaculture 2023, 568, 739315. [Google Scholar] [CrossRef]
- Bloecher, N.; Floerl, O. Efficacy Testing of Novel Antifouling Coatings for Pen Nets in Aquaculture: How Good Are Alternatives to Traditional Copper Coatings? Aquaculture 2020, 519, 734936. [Google Scholar] [CrossRef]
- Ashraf, P.M.; Lekshmi, N.M.; Chinnadurai, S.; Anjitha, S.; Archana, M.; Kumar, C.M.V.; Sandhya, K.M.; Gop, A.P. Impact Assessment of Biofouling Resistant Nano Copper Oxide–Polyaniline Coating on Aquaculture Cage Nets. Aquac. Fish 2023, 8, 538–543. [Google Scholar] [CrossRef]
- Haberlin, D.; McAllen, R.; Doyle, T.K. Field and Flume Tank Experiments Investigating the Efficacy of a Bubble Curtain to Keep Harmful Jellyfish out of Finfish Pens. Aquaculture 2021, 531, 735915. [Google Scholar] [CrossRef]
- Ohrem, S.J.; Kelasidi, E.; Bloecher, N. Analysis of a Novel Autonomous Underwater Robot for Biofouling Prevention and Inspection in Fish Farms. In Proceedings of the 2020 28th Mediterranean Conference on Control and Automation (MED), Saint-Raphaël, France, 15–18 September 2020; IEEE: Piscataway, NJ, USA, 2020; pp. 1002–1008. [Google Scholar]
- Haugaløkken, B.O.A.; Kelasidi, E.; Mulelid, M.; Bloecher, N. Docking Stations for Net-Crawling Underwater Vehicles in Aquaculture Net Pens. In Proceedings of the OCEANS 2021: San Diego–Porto, San Diego, CA, USA, 20–23 September 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 1–10. [Google Scholar]
- Boissonnot, L.; Kharlova, I.; Iversen, N.S.; Staven, F.R.; Austad, M. Characteristics of Lumpfish (Cyclopterus lumpus) with High Cleaning Efficacy in Commercial Atlantic Salmon (Salmo salar) Production. Aquaculture 2022, 560, 738544. [Google Scholar] [CrossRef]
- Imsland, A.K.D.; Reynolds, P. In Lumpfish We Trust? The Efficacy of Lumpfish Cyclopterus lumpus to Control Lepeophtheirus salmonis Infestations on Farmed Atlantic Salmon: A Review. Fishes 2022, 7, 220. [Google Scholar] [CrossRef]
- Leclercq, E.; Zerafa, B.; Brooker, A.J.; Davie, A.; Migaud, H. Application of Passive-Acoustic Telemetry to Explore the Behaviour of Ballan Wrasse (Labrus bergylta) and Lumpfish (Cyclopterus lumpus) in Commercial Scottish Salmon Sea-Pens. Aquaculture 2018, 495, 1–12. [Google Scholar] [CrossRef]
- Zeinert, L.R.; Brooks, A.M.L.; Couturier, C.; McGaw, I.J. Potential Use of the Caribbean Spider Crab Maguimithrax spinosissimus for Biofouling Removal on Marine Aquaculture Cages. Aquaculture 2021, 545, 737202. [Google Scholar] [CrossRef]
- Ahlgren, M.O. Consumption and Assimilation of Salmon Net Pen Fouling Debris by the Red Sea Cucumber Parastichopus californicus: Implications for Polyculture. J. World Aquac. Soc. 1998, 29, 133–139. [Google Scholar] [CrossRef]
- Hannah, L.; Pearce, C.M.; Cross, S.F. Growth and Survival of California Sea Cucumbers (Parastichopus californicus) Cultivated with Sablefish (Anoplopoma fimbria) at an Integrated Multi-Trophic Aquaculture Site. Aquaculture 2013, 406–407, 34–42. [Google Scholar] [CrossRef]
- Tolon, M.T.; Emiroglu, D.; Gunay, D.; Ozgul, A. Sea Cucumber (Holothuria tubulosa Gmelin, 1790) Culture under Marine Fish Net Cages for Potential Use in Integrated Multi-Trophic Aquaculture (IMTA). Indian J. Geo-Mar. Sci. 2017, 46, 749–756. [Google Scholar]
- Zamora, L.N.; Yuan, X.; Carton, A.G.; Slater, M.J.; Marine, L. Role of Deposit-Feeding Sea Cucumbers in Integrated Multitrophic Aquaculture: Progress, Problems, Potential and Future Challenges. Rev. Aquac. 2018, 10, 57–74. [Google Scholar] [CrossRef]
- Cutajar, K.; Falconer, L.; Massa-Gallucci, A.; Cox, R.E.; Schenke, L.; Bardócz, T.; Sharman, A.; Deguara, S.; Telfer, T.C. Culturing the Sea Cucumber Holothuria poli in Open-Water Integrated Multi-Trophic Aquaculture at a Coastal Mediterranean Fish Farm. Aquaculture 2022, 550, 737881. [Google Scholar] [CrossRef]
- Purcell, S.W.; Mercier, A.; Conand, C.; Hamel, J.; Toral-Granda, M.V.; Lovatelli, A.; Uthicke, S. Sea Cucumber Fisheries: Global Analysis of Stocks, Management Measures and Drivers of Overfishing. Fish Fish. 2013, 14, 34–59. [Google Scholar] [CrossRef]
- Darya, M.; Sajjadi, M.; Yousefzadi, M.; Sourinejad, I.; Zarei, M. Bioactive Compounds of Sea Cucumber (Holothuria leucospilota) Added to Epoxy Resin as Environmental Friendly Antifouling Coats. Aquat. Physiol. Biotechnol. 2020, 8, 47–66. [Google Scholar]
- Darya, M.; Abdolrasouli, M.H.; Yousefzadi, M.; Sajjadi, M.M.; Sourinejad, I.; Zarei, M. Antifouling Coating Based on Biopolymers (PCL/PLA) and Bioactive Extract from the Sea Cucumber Stichopus herrmanni. AMB Express 2022, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Kamyab, E.; Goebeler, N.; Kellermann, M.Y.; Rohde, S.; Reverter, M.; Striebel, M.; Schupp, P.J. Anti-Fouling Effects of Saponin-Containing Crude Extracts from Tropical Indo-Pacific Sea Cucumbers. Mar. Drugs 2020, 18, 181. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar]
- Gansel, L.C.; Bloecher, N.; Floerl, O.; Guenther, J. Quantification of Biofouling on Nets: A Comparison of Wet Weight Measurements and Optical (Image Analysis) Methods. Aquac. Int. 2017, 25, 679–692. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2; Use R! Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only ANOVA Procedures. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Yokohama, Japan, 8–13 May 2011; pp. 143–146. [Google Scholar]
- Leys, C.; Schumann, S. A Nonparametric Method to Analyze Interactions: The Adjusted Rank Transform Test. J. Exp. Soc. Psychol. 2010, 46, 684–688. [Google Scholar] [CrossRef]
- Strain, P.M.; Hargrave, B.T. Salmon Aquaculture, Nutrient Fluxes and Ecosystem Processes in Southwestern New Brunswick. In Environmental Effects of Marine Finfish Aquaculture; Springer: Berlin/Heidelberg, Germany, 2005; pp. 29–57. [Google Scholar]
- Hodson, S.L.; Burke, C. Microfouling of Salmon-Cage Netting: A Preliminary Investigation. Biofouling 1994, 8, 93–105. [Google Scholar] [CrossRef]
- Bloecher, N.; Olsen, Y.; Guenther, J. Variability of Biofouling Communities on Fish Cage Nets: A 1-Year Field Study at a Norwegian Salmon Farm. Aquaculture 2013, 416–417, 302–309. [Google Scholar] [CrossRef]
- Bloecher, N.; Powell, M.; Hytterød, S.; Gjessing, M.; Wiik-Nielsen, J.; Mohammad, S.N.; Johansen, J.; Hansen, H.; Floerl, O.; Gjevre, A.G. Effects of Cnidarian Biofouling on Salmon Gill Health and Development of Amoebic Gill Disease. PLoS ONE 2018, 13, e0199842. [Google Scholar] [CrossRef]
- Park, H.J.; Han, E.; Lee, W.C.; Kwak, J.H.; Kim, H.C.; Park, M.S.; Kang, C.K. Trophic Structure in a Pilot System for the Integrated Multi-Trophic Aquaculture off the East Coast of Korean Peninsula as Determined by Stable Isotopes. Mar. Pollut. Bull. 2015, 95, 207–214. [Google Scholar] [CrossRef]
- Sadoul, B.; Caprioli, J.-P.; Barrier-Loiseau, C.; Cimiterra, N.; Laugier, T.; Lagarde, F.; Chary, K.; Callier, M.D.; Guillermard, M.-O.; D’Orbcastel, E.R. Is Holothuria tubulosa the Golden Goose of Ecological Aquaculture in the Mediterranean Sea? Aquaculture 2022, 554, 738149. [Google Scholar] [CrossRef]
- Cutajar, K.; Falconer, L.; Massa-Gallucci, A.; Cox, R.E.; Schenke, L.; Bardócz, T.; Andolina, C.; Signa, G.; Vizzini, S.; Sprague, M.; et al. Stable Isotope and Fatty Acid Analysis Reveal the Ability of Sea Cucumbers to Use Fish Farm Waste in Integrated Multi-Trophic Aquaculture. J. Environ. Manag. 2022, 318, 115511. [Google Scholar] [CrossRef] [PubMed]
- Orr, L.C. Co-Culture of Invertebrates with Sablefish (Anoplopoma fimbria) in IMTA in British Columbia: Use of Laboratory Feeding Trials to Assess the Organic Extractive Potential of Various Candidate Species. Master’s Thesis, University of Victoria, Victoria, BC, Canada, 2012. [Google Scholar]
- Hudson, B.; Hauser, L.; Vadopalas, B.; Hetrick, J.; Pride, A.; Hatchery, S.; Seward, H.; Carson, W. Development of Red Sea Cucumber (Parastichopus californicus) Poly-Aquaculture for Nutrient Uptake and Seafood Export; Final Report of the Saltonstall-Kennedy Program #NA15NMF4270322; 2019; Available online: https://www.pacshell.org/pdf/Final%20Report%20SK%20Cucumber%202019.pdf (accessed on 28 June 2023).
- Sun, J.; Hamel, J.-F.; Gianasi, B.L.; Graham, M.; Mercier, A. Growth, Health and Biochemical Composition of the Sea Cucumber Cucumaria frondosa after Multi-Year Holding in Effluent Waters of Land-Based Salmon Culture. Aquac. Environ. Interact. 2020, 12, 139–151. [Google Scholar] [CrossRef]
- Li, X.; Dong, Y.; Jiang, P.; Qi, L.; Lin, S. Identification of Changes in Volatile Compounds in Sea Cucumber Apostichopus japonicus during Seasonings Soaking Using HS-GC-IMS. LWT 2022, 154, 112695. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Mean ± SD (%) | ART Two-Way ANOVA | |
|---|---|---|---|
| Trial month | August | 59.2 ± 25.8 | NS |
| September | 70.8 ± 16.5 | ||
| October | 72.8 ± 24.3 | ||
| Site | Dawley Pass | 68.8 ± 23.0 | NS |
| Warne Island | 66.3 ± 24.9 | ||
| Trial month × site | NS | ||
| Cage type | Fish only (F) | 64.6 ± 25.5 | NS |
| Sea cucumbers only (S) | 79.4 ± 16.9 | ||
| Fish + sea cucumbers (F + S) | 62.1 ± 24.9 | ||
| Empty cage (E) | 64.2 ± 21.6 | ||
| Net depth * | Top (0–5 m) | 46.3 ± 33.5 A | F2,60 = 8.28, p < 0.01 |
| Middle (5–10 m) | 79.0 ± 18.1 B | ||
| Bottom (10–15 m) | 77.5 ± 20.0 B | ||
| Cage type × net depth | NS | ||
| Net Depth | Cage Type, Mean ± SD (%) | ART One-Way ANOVA | |||
|---|---|---|---|---|---|
| Empty Cage (E) | Fish (F) | Fish + Sea Cucumbers (F + S) | Sea Cucumbers (S) | ||
| Top (0–5 m) | 33.0 ± 37.1 | 49.1 ± 37.9 | 45.5 ± 26.4 | 57.5 ± 35.8 | NS |
| Middle (5–10 m) | 76.3 ± 18.5 | 79.4 ± 13.0 | 73.7 ± 28.6 | 86.7 ± 8.2 | NS |
| Bottom (10–15 m) * | 83.2 ± 9.2 AB | 65.3 ± 25.8 A | 67.2 ± 19.5 A | 94.2 ± 6.8 B | F3,20 = 4.32, p = 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montgomery, E.M.; Cannon, B.L.; Pearce, C.M. Exploring Biofouling Control by the California Sea Cucumber (Apostichopus californicus) in Integrated Multi-Trophic Aquaculture (IMTA) with Organic Chinook Salmon (Oncorhynchus tshawytscha). Fishes 2023, 8, 430. https://doi.org/10.3390/fishes8090430
Montgomery EM, Cannon BL, Pearce CM. Exploring Biofouling Control by the California Sea Cucumber (Apostichopus californicus) in Integrated Multi-Trophic Aquaculture (IMTA) with Organic Chinook Salmon (Oncorhynchus tshawytscha). Fishes. 2023; 8(9):430. https://doi.org/10.3390/fishes8090430
Chicago/Turabian StyleMontgomery, Emaline M., Barb L. Cannon, and Christopher M. Pearce. 2023. "Exploring Biofouling Control by the California Sea Cucumber (Apostichopus californicus) in Integrated Multi-Trophic Aquaculture (IMTA) with Organic Chinook Salmon (Oncorhynchus tshawytscha)" Fishes 8, no. 9: 430. https://doi.org/10.3390/fishes8090430
APA StyleMontgomery, E. M., Cannon, B. L., & Pearce, C. M. (2023). Exploring Biofouling Control by the California Sea Cucumber (Apostichopus californicus) in Integrated Multi-Trophic Aquaculture (IMTA) with Organic Chinook Salmon (Oncorhynchus tshawytscha). Fishes, 8(9), 430. https://doi.org/10.3390/fishes8090430