
A Comparison of the Physiological Responses to Heat Stress of Two Sizes of Juvenile Spotted Seabass (Lateolabrax maculatus)

Abstract
1. Introduction
2. Materials and Methods
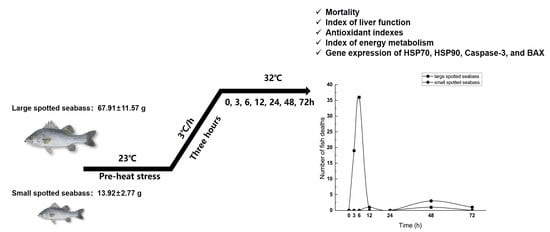
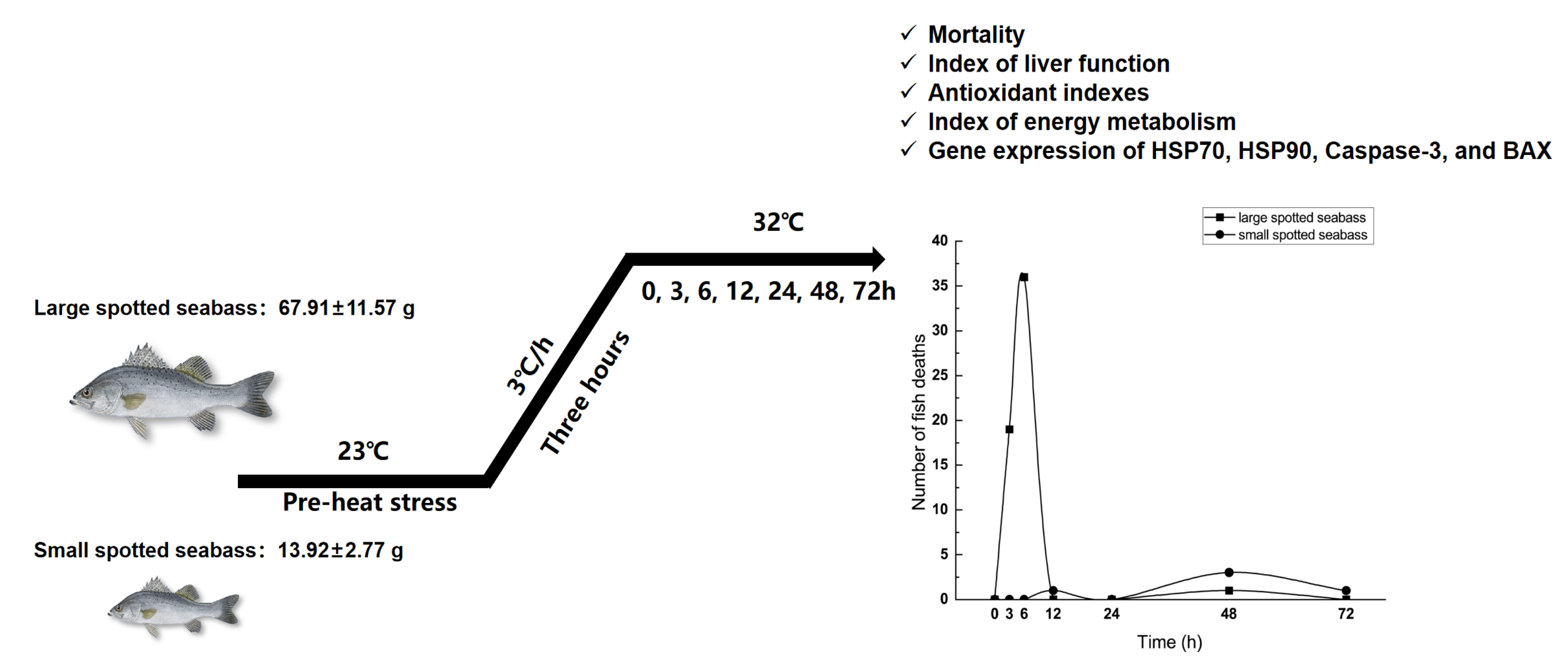
2.1. Experimental Fish and Rearing Condition
2.2. Heat Stress Test and Sample Collection
2.3. Methods for the Detection of Biochemical Indicator
2.4. RNA Extraction and Reverse Transcription
2.5. Real-Time Quantitative PCR (RT-qPCR) Reaction
2.6. Data Statistics and Analysis
3. Results
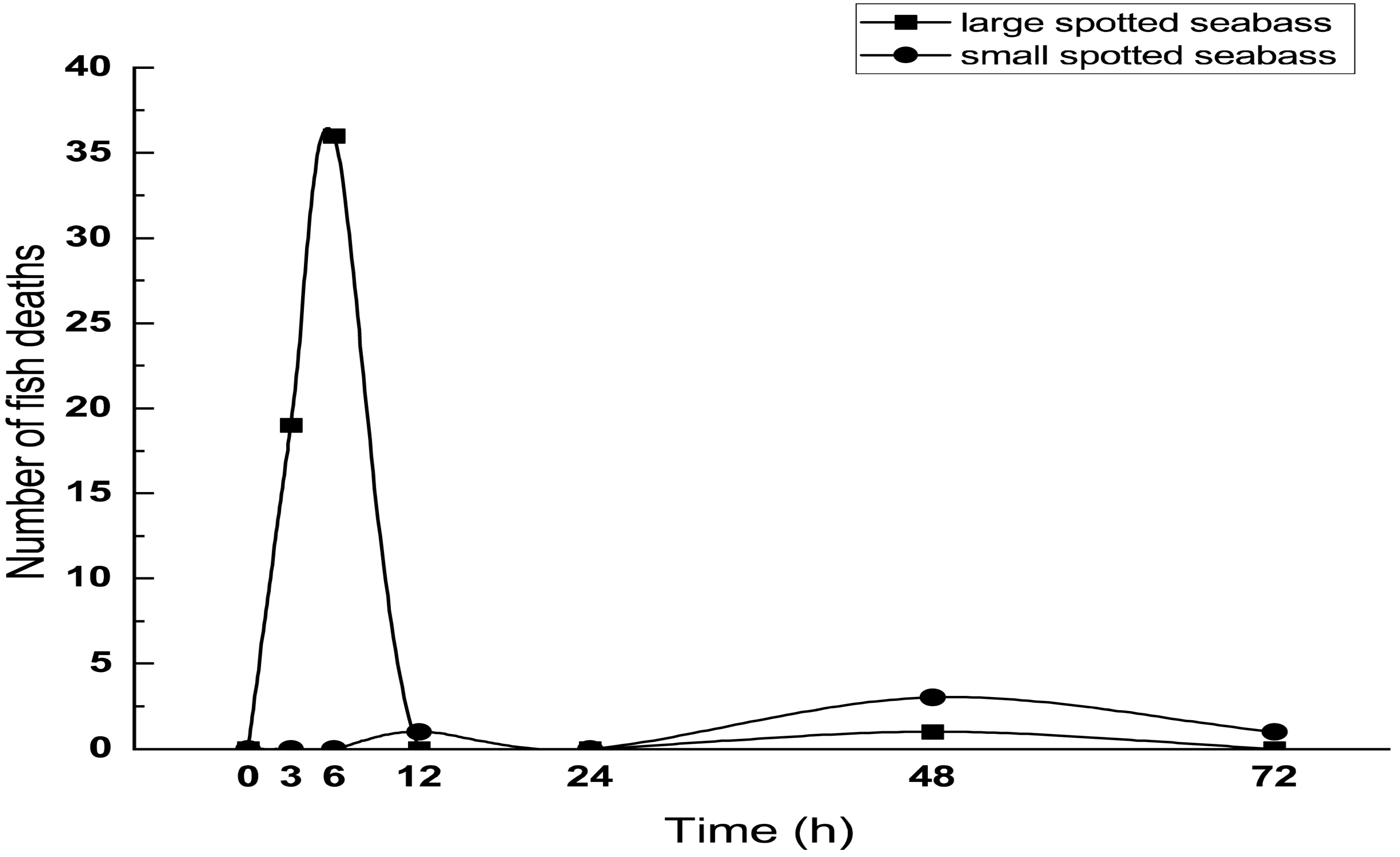
3.1. Mortality
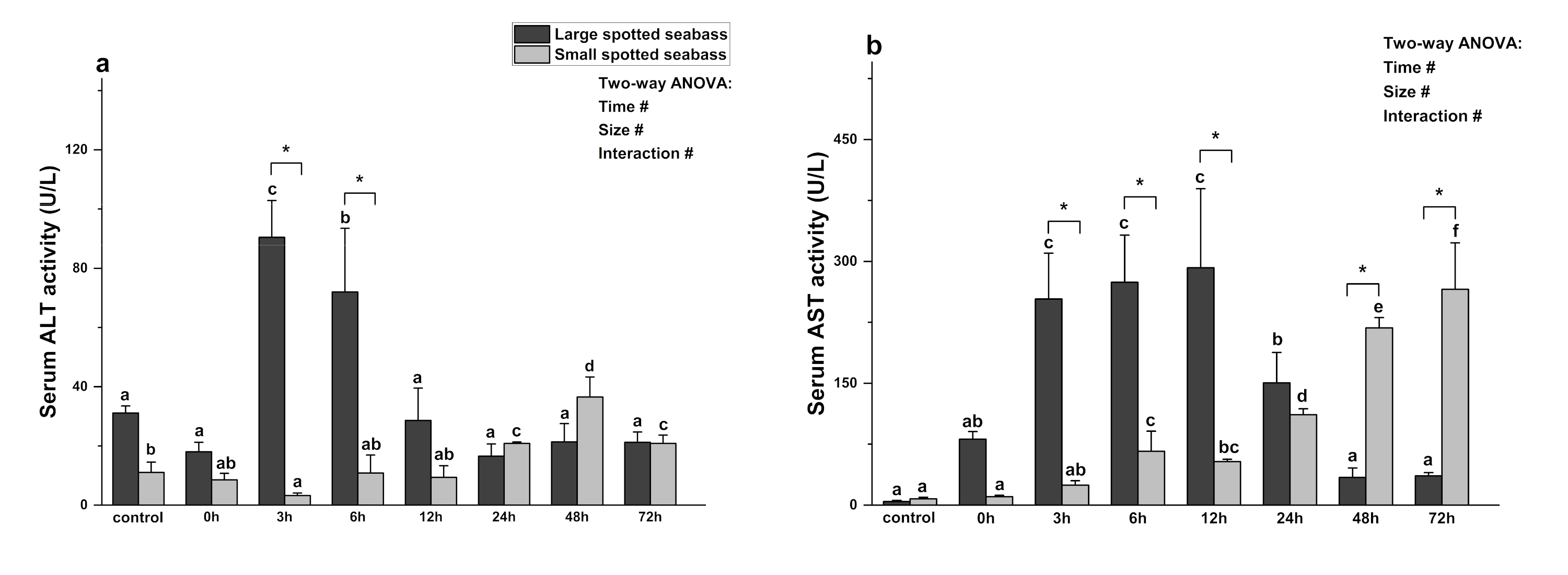
3.2. The Effect of Heat Stress on Index of Liver Function of Spotted Seabass
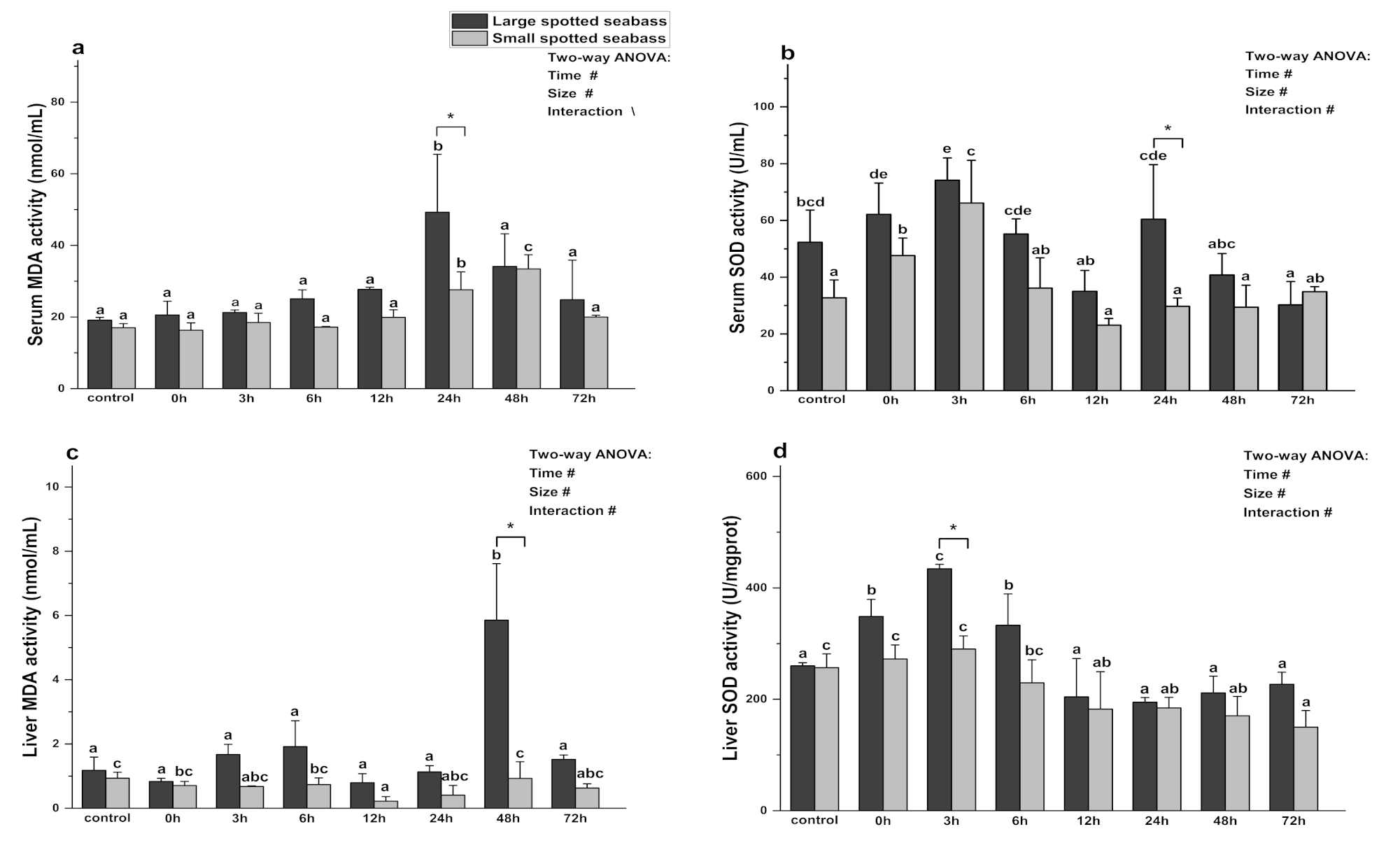
3.3. The Effect of Heat Stress on Antioxidant Indexes in Spotted Seabass Serum and Liver
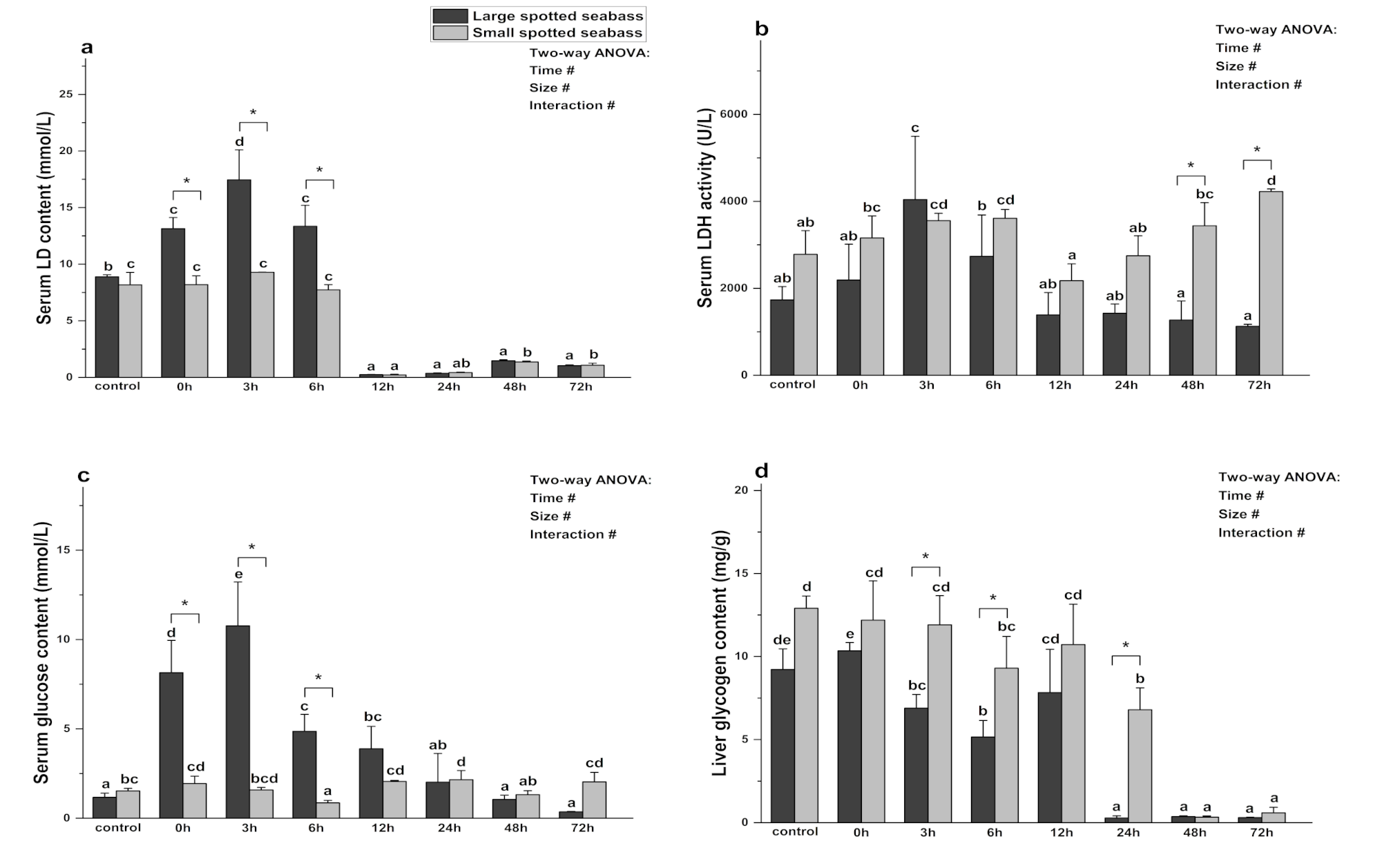
3.4. The Effect of Heat Stress on Spotted Seabass Energy Metabolism
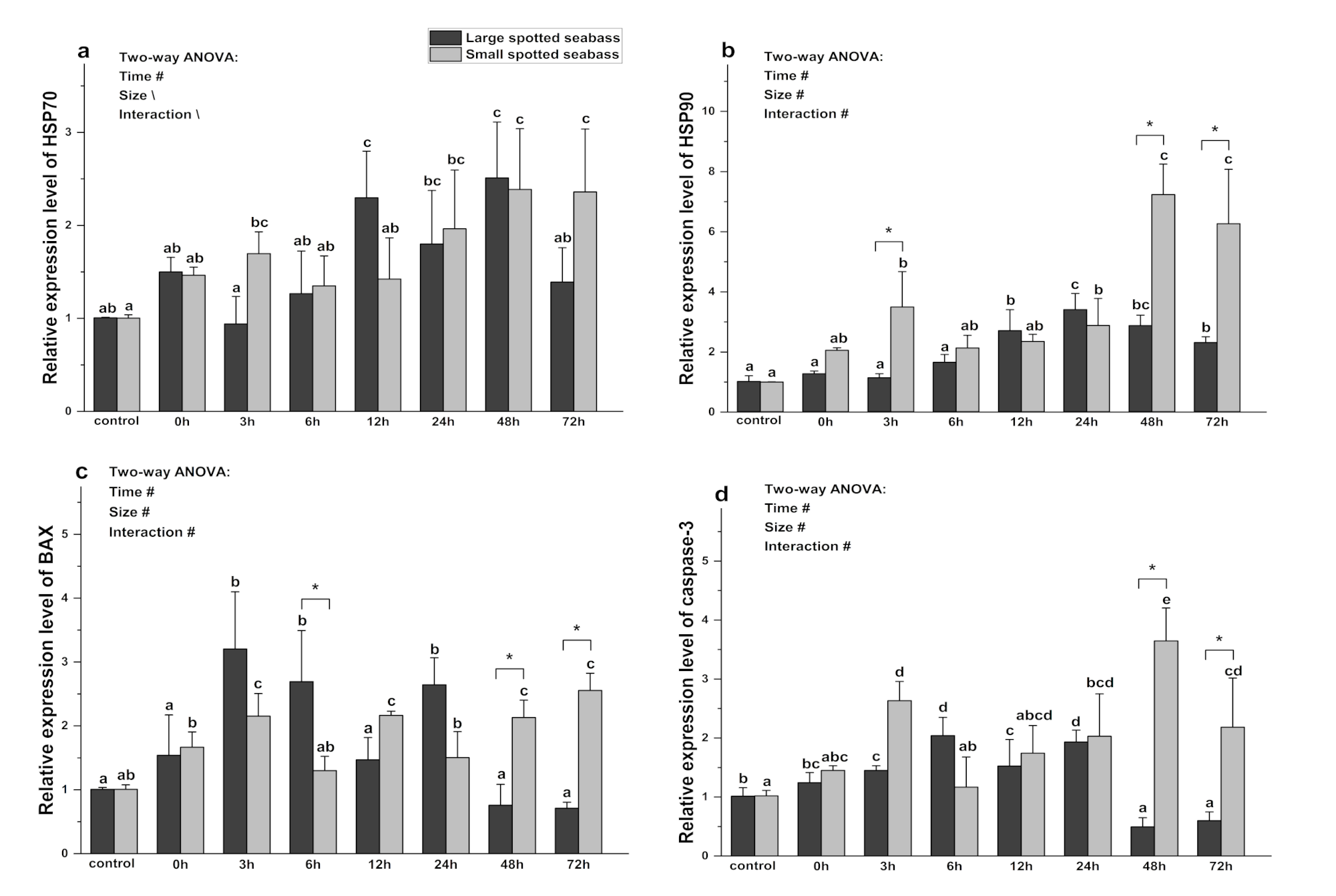
3.5. The Effect of Heat Stress on Gene Expression of HSP70, HSP90, Caspase-3, and BAX in Spotted Seabass
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- López-Olmeda, J.F.; Sánchez-Vázquez, F.J. Thermal biology of zebrafish (Danio rerio). J. Therm. Biol. 2011, 36, 91–104. [Google Scholar] [CrossRef]
- Allen, S.K.; Barros, V.; Burton, I.; Campbell-Lendrum, D.; Cardona, O.-D.; Cutter, S.L.; Dube, O.P.; Ebi, K.L.; Field, C.B.; Handmer, J.W.; et al. Summary for Policymakers. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Cambridge University Press: Cambridge, UK, 2012; pp. 3–22. [Google Scholar]
- Julia, S.; Kurt, P.; Ricardo, O.-S.; Christina, C.C.-H.; Navarro, J.M.; Luis, V.-C. Effects of warming rates on physiological and molecular components of response to CTMax heat stress in the Antarctic fish Harpagifer antarcticus. J. Therm. Biol. 2021, 99, 103021. [Google Scholar] [CrossRef]
- Imsland, A.K.; Sunde, L.M.; Folkvord, A.; Stefansson, S.O. The interaction of temperature and fish size on growth of juvenile turbot. J. Fish Biol. 1996, 49, 926–940. [Google Scholar] [CrossRef]
- Fraser, E.J.; Bosma, P.T.; Trudeau, V.L.; Docherty, K. The Effect of Water Temperature on the GABAergic and Reproductive Systems in Female and Male Goldfish (Carassius auratus). Gen. Comp. Endocrinol. 2002, 125, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Langston, A.L.; Hoare, R.; Stefansson, M.; Fitzgerald, R.; Wergeland, H.; Mulcahy, M. The effect of temperature on non-specific defence parameters of three strains of juvenile Atlantic halibut (Hippoglossus hippoglossus L.). Fish Shellfish. Immunol. 2002, 12, 61–76. [Google Scholar] [CrossRef]
- Miranda, L.A.; Chalde, T.; Elisio, M.; Strüssmann, C.A. Effects of global warming on fish reproductive endocrine axis, with special emphasis in pejerrey Odontesthes bonariensis. Gen. Comp. Endocrinol. 2013, 192, 45–54. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P. OXIDATIVE STRESS IN MARINE ENVIRONMENTS: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Wu, Y.; Huang, D.; Ren, X.; Wang, Y. Effect of blood glucose level on acute stress response of grass carp Ctenopharyngodon idella. Fish Physiol. Biochem. 2017, 43, 1433–1442. [Google Scholar] [CrossRef]
- Pottinger, T.G.; Carrick, T.R. A comparison of plasma glucose and plasma cortisol as selection markers for high and low stress-responsiveness in female rainbow trout. Aquaculture 1999, 175, 351–363. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Zheng, J.-L.; Peng, L.-B.; Zhu, Q.-L.; Zhang, X.-L.; Hu, W. Waterborne zinc induced lobe-dependent effect on oxidative stress and energy metabolism in hepatopancreas of Larimichthys crocea. Aquat. Toxicol. 2019, 215, 105270. [Google Scholar] [CrossRef]
- Vaglio, A.; Landriscina, C. Changes in Liver Enzyme Activity in the TeleostSparus aurata in Response to Cadmium Intoxication. Ecotoxicol. Environ. Saf. 1999, 43, 111–116. [Google Scholar] [CrossRef]
- Al-Deghayem, W.A.A.; Suliman, E.A.M. The effects of diet and temperature on enzymes, hormones, and fecundity of the African Catfish Clarias gariepinus (Burchell 1822). J. Appl. Biol. Biotechnol. 2019, 7, 71–77. [Google Scholar] [CrossRef]
- Yan, L.; Wang, P.; Zhao, C.; Fan, S.; Lin, H.; Guo, Y.; Ma, Z.; Qiu, L. Toxic responses of liver in Lateolabrax maculatus during hypoxia and re-oxygenation. Aquat. Toxicol. 2021, 236, 105841. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.P.; Mahanty, A.; Mitra, T.; Parija, S.C.; Mohanty, S. Heat shock proteins in stress in teleosts. Regul. Heat Shock. Protein Responses 2018, 13, 71–94. [Google Scholar]
- Takayama, S.; Reed, J.C.; Homma, S. Heat-shock proteins as regulators of apoptosis. Oncogene 2003, 22, 9041–9047. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Chen, X.; Wang, Z.; Meng, Z.; Huang, B.; Guan, C. Physiological response of juvenile turbot (Scophthalmus maximus L.) during hyperthermal stress. Aquaculture 2020, 529, 735645. [Google Scholar] [CrossRef]
- Madeira, D.; Vinagre, C.; Diniz, M.S. Are fish in hot water? Effects of warming on oxidative stress metabolism in the commercial species Sparus aurata. Ecol. Indic. 2016, 63, 324–331. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The Chaperone Function of hsp70 Is Required for Protection against Stress-Induced Apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Li, T.; Zeng, L.; Gao, W.; Cui, M.-Z.; Fu, X.; Xu, X. PSAP induces a unique Apaf-1 and Smac-dependent mitochondrial apoptotic pathway independent of Bcl-2 family proteins. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2013, 1832, 453–474. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Shang-Guan, J.; Li, Z.; Gao, Z.; Huang, Y.C.; Chen, Q. Effects of dietary Chinese herbal medicines mixture on feeding attraction activity, growth performance, nonspecific immunity and digestive enzyme activity of Japanese seabass (Lateolabrax japonicus). Aquac. Rep. 2020, 17, 100304. [Google Scholar] [CrossRef]
- Liu, X. Cultivation techniques for seabass in Bai Jiao. Contemp. Fish. 2010, 35, 72–73. (In Chinese) [Google Scholar]
- Zhao, W. Key points of the freshwater culture technology of Lateolabrax japonicus. J. Beijing Fish. 2002, 5, 22. (In Chinese) [Google Scholar]
- Bevelhimer, M.; Bennett, W. Assessing cumulative thermal stress in fish during chronic intermittent exposure to high temperatures. Environ. Sci. Policy 2000, 3, 211–216. [Google Scholar] [CrossRef]
- Huang, Z.; Li, Z.; Xu, A.; Zheng, D.; Ye, Y.; Wang, Z. Effects of exogenous multienzyme complex supplementation in diets on growth performance, digestive enzyme activity and non-specific immunity of the Japanese seabass, Lateolabrax japonicus. Aquac. Nutr. 2020, 26, 306–315. [Google Scholar] [CrossRef]
- Shen, Q.; Xu, S.; Wang, D.; Yan, X. Effects of water temperature on metabolizability of starved Lateolabrax japonicus. J. Fish. Sci. China 2008, 3, 500–505. (In Chinese) [Google Scholar]
- Shin, M.-K.; Park, H.-R.; Yeo, W.-J.; Han, K.-N. Effects of Thermal Stress on the mRNA Expression of SOD, HSP90, and HSP70 in the Spotted seabass (Lateolabrax maculatus). Ocean. Sci. J. 2018, 53, 43–52. [Google Scholar] [CrossRef]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.-O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65–70. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, X.; Liang, X.; Wu, X.; Gu, X.; Han, J.; Xue, M. N-carbamoylglutamate improves lipid metabolism, inflammation, and apoptosis responses in visceral adipocytes of Japanese seabass (Lateolabrax japonicus), in vivo and in vitro. Anim. Nutr. 2021, 7, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lu, K.; Wang, l.; Song, K.; Zhang, C. Effects of berberine supplementation in feed on the growth and fat metabolism of Japanese seabass (Lateolabrax japonicus). Chin. J. Anim. Nutr. 2021, 33, 5193–5202. (In Chinese) [Google Scholar]
- Phrompanya, P.; Panase, P.; Saenphet, S.; Saenphet, K. Histopathology and oxidative stress responses of Nile tilapia Oreochromis niloticus exposed to temperature shocks. Fish. Sci. 2021, 87, 491–502. [Google Scholar] [CrossRef]
- He, L.Q.; Wang, W.L.; Zeng, B.H.; Yang, R.B.; Liu, H.P.; Zeng, X.L.; Xu, Z.L.; Wang, J. The study on the temperature tolerance of different sizes of Schizothorax waltoni. Acta Hydrobiol. Sin. 2020, 44, 1231–1238. (In Chinese) [Google Scholar]
- Cheng, C.-H.; Guo, Z.-X.; Luo, S.-W.; Wang, A.-L. Effects of high temperature on biochemical parameters, oxidative stress, DNA damage and apoptosis of pufferfish (Takifugu obscurus). Ecotoxicol. Environ. Saf. 2018, 150, 190–198. [Google Scholar] [CrossRef]
- Ming, J.; Xie, J.; Xu, P.; Ge, X.; Liu, W.; Ye, J. Effects of emodin and vitamin C on growth performance, biochemical parameters and two HSP70s mRNA expression of Wuchang bream (Megalobrama amblycephala Yih) under high temperature stress. Fish Shellfish Immunol. 2012, 32, 651–661. [Google Scholar] [CrossRef]
- Klein, R.D.; Borges, V.D.; Rosa, C.E.; Colares, E.P.; Robaldo, R.B.; Martinez, P.E.; Bianchini, A. Effects of increasing temperature on antioxidant defense system and oxidative stress parameters in the Antarctic fish Notothenia coriiceps and Notothenia rossii. J. Therm. Biol. 2017, 68, 110–118. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Noreldin, A.E.; Sewilam, H. Blood biochemical variables, antioxidative status, and histological features of intestinal, gill, and liver tissues of African catfish (Clarias gariepinus) exposed to high salinity and high-temperature stress. Environ. Sci. Pollut. Res. 2022, 29, 56357–56369. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.; Li, Z.; Shi, H.; Kang, Y.; Wang, J.; Huang, J.; Jiang, L. Effects of heat stress on respiratory burst, oxidative damage and SERPINH1 (HSP47) mRNA expression in rainbow trout Oncorhynchus mykiss. Fish Physiol. Biochem. 2016, 42, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.J.; Kunzmann, A.; Slater, M.J. Extreme winter cold-induced osmoregulatory, metabolic, and physiological responses in European seabass (Dicentrarchus labrax) acclimatized at different salinities. Sci. Total Environ. 2021, 771, 145202. [Google Scholar] [CrossRef] [PubMed]
- Handy, J. Lactate—The bad boy of metabolism, or simply misunderstood? Curr. Anaesth. Crit. Care 2006, 17, 71–76. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, Z.; Song, Z.; Xiao, P.; Liu, Y.; Zhang, P.; You, F. Insight into the heat resistance of fish via blood: Effects of heat stress on metabolism, oxidative stress and antioxidant response of olive flounder Paralichthys olivaceus and turbot Scophthalmus maximus. Fish Shellfish Immunol. 2016, 58, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Mackey, T.E.; Hasler, C.T.; Durhack, T.; Jeffrey, J.D.; Macnaughton, C.J.; Ta, K.; Enders, E.C.; Jeffries, K.M. Molecular and physiological responses predict acclimation limits in juvenile brook trout (Salvelinus fontinalis). J. Exp. Biol. 2021, 224, 241885. [Google Scholar] [CrossRef]
- Zhang, H.; Jia, H.; Xiong, P.; Yao, G.; He, M. Transcriptome and enzyme activity analyses of tolerance mechanisms in pearl oyster (Pinctada fucata) under high-temperature stress. Aquaculture 2022, 550, 737888. [Google Scholar] [CrossRef]
- Lee, H.B.; Yoon, J.H.; Park, J.Y.; Lee, I.Y.; Lim, H.K. A comparison of the physiological responses to heat stress of juvenile and adult starry flounder (Platichthys stellatus). Isr. J. Aquac. Bamidgeh 2021, 73, 1–15. [Google Scholar] [CrossRef]
- Healy, T.M.; Tymchuk, W.E.; Osborne, E.J.; Schulte, P.M. Heat shock response of killifish (Fundulus heteroclitus): Candidate gene and heterologous microarray approaches. Physiol. Genom. 2010, 41, 171–184. [Google Scholar] [CrossRef]
- Stefanovic, D.I.; Manzon, L.A.; McDougall, C.S.; Boreham, D.R.; Somers, C.M.; Wilson, J.Y.; Manzon, R.G. Thermal stress and the heat shock response in embryonic and young of the year juvenile lake whitefish. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2016, 193, 1–10. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, C.; Xing, W.; Li, T.; Xu, G.; Ma, Z.; Jiang, N.; Luo, L. Comparative study on the non-specific immune response and hsp70 gene expression among three strains of koi (Cyprinus carpio) under acute heat stress. Aquac. Rep. 2020, 18, 100461. [Google Scholar] [CrossRef]
- Webb, P.W.; Gans, C. Is Bigger Better? BioScience 1986, 36, 340–342. [Google Scholar] [CrossRef]
- Suzanne, F.S.L.H.D.C. A comparison of the heat shock response in juvenile and adult rainbow trout (Oncorhynchus mykiss)—Implications for increased thermal sensitivity with age. Can. J. Fish. Aquat. Sci. 2009, 66, 91–100. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Yang, F.-F.; Liao, S.-A.; Miao, Y.-T.; Ye, C.-X.; Wang, A.-L.; Tan, J.-W.; Chen, X.-Y. High temperature induces apoptosis and oxidative stress in pufferfish (Takifugu obscurus) blood cells. J. Therm. Biol. 2015, 53, 172–179. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence 5′-3′ | References |
|---|---|---|
| Caspase-3 | F: TGACCAGACAGTCGAGCAGA | [35] |
| R: AACACACCCTCATCTCCGTG | ||
| BAX | F: TGGAAGTGGGAGCTGTCTTG | [36] |
| R: GGCGAGCTTCTTGTGGTTTG | ||
| HSP90 | F: ACACAACGATGATGAGCAGT | [33] |
| R: TCAGGTGCAGGATGATCTTT | ||
| HSP70 | F: CCCAAGGTCCAAGTTGAGT | [33] |
| R: AGGTAGGCTTCAGCAATCTC | ||
| β-actin | F: CGCCGCACTGGTTGTT | [33] |
| R: TTTGGGGTTCAGGGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, H.; Long, Z.; Huang, Z.; Ma, J.; Kong, L.; Lin, Y.; Lin, H.; Zhou, S.; Li, Z. A Comparison of the Physiological Responses to Heat Stress of Two Sizes of Juvenile Spotted Seabass (Lateolabrax maculatus). Fishes 2023, 8, 340. https://doi.org/10.3390/fishes8070340
Qin H, Long Z, Huang Z, Ma J, Kong L, Lin Y, Lin H, Zhou S, Li Z. A Comparison of the Physiological Responses to Heat Stress of Two Sizes of Juvenile Spotted Seabass (Lateolabrax maculatus). Fishes. 2023; 8(7):340. https://doi.org/10.3390/fishes8070340
Chicago/Turabian StyleQin, Huihui, Zhongying Long, Zhangfan Huang, Jianrong Ma, Lumin Kong, Yi Lin, Hao Lin, Sishun Zhou, and Zhongbao Li. 2023. "A Comparison of the Physiological Responses to Heat Stress of Two Sizes of Juvenile Spotted Seabass (Lateolabrax maculatus)" Fishes 8, no. 7: 340. https://doi.org/10.3390/fishes8070340
APA StyleQin, H., Long, Z., Huang, Z., Ma, J., Kong, L., Lin, Y., Lin, H., Zhou, S., & Li, Z. (2023). A Comparison of the Physiological Responses to Heat Stress of Two Sizes of Juvenile Spotted Seabass (Lateolabrax maculatus). Fishes, 8(7), 340. https://doi.org/10.3390/fishes8070340