
A Synthesis of the Ecology and Conservation of Pseudoplatystoma Catfishes in the Neotropics
by
 , and
, and
Luciana A. Pereira
1,*,
Leandro Castello
1,
Donald J. Orth
1,
Fabrice Duponchelle
2 and
Eric M. Hallerman
1
1
Department of Fish & Wildlife Conservation, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USA
2
Institut de Recherche pour le Développement (IRD), MARBEC (Université Montpellier, CNRS, Ifremer, IRD), 34095 Montpellier, France
*
Author to whom correspondence should be addressed.
Fishes 2023, 8(6), 306; https://doi.org/10.3390/fishes8060306
Submission received: 28 April 2023
/
Revised: 26 May 2023
/
Accepted: 31 May 2023
/
Published: 7 June 2023
(This article belongs to the Section Biology and Ecology)
Abstract
:Species of the genus Pseudoplatystoma, the long-whiskered catfishes, are important in commercial and recreational fisheries in South America, and some species have become key to regional aquaculture. Some species of the genus are under pressure due to overfishing and the negative impacts of dams. Six questions are asked in this review: (i) What species are in the genus, and where are they distributed? (ii) What are the life histories and ecologies of Pseudoplatystoma species? (iii) What are the patterns of somatic growth for these species? (iv) What is known about the biomass, production, and population dynamics of Pseudoplatystoma? (v) What is the geographic distribution of genetic variation within Pseudoplatystoma species? (vi) What are the threats to the conservation of these species? The taxonomy of the genus currently includes eight species, respectively, distributed over the Orinoco, Amazon, Paraná, and São Francisco basins. Pseudoplatystoma catfishes typically exhibit longitudinal migrations for reproduction and lateral migration for feeding, but these patterns may vary among populations. The size of the first maturation of these catfishes varies between 57 cm to 82 cm in total length. Five of the eight species spawn during the rising water season. Pseudoplatystoma species can grow to about 130 cm in total length and 100 Kg in weight and live until 30 years of age, depending upon the species. Biomass production and population dynamics of these catfishes have not yet been fully described. Their life-history characteristics indicate that they are periodic strategists with associated population recruitment dynamics. Population genetic patterning varies among Pseudoplatystoma species, with some degree of homing behavior and genetic differentiation among populations, indicating the need for management by applying the Management Unit and perhaps Evolutionary Significant Unit concepts. The main threats to the persistence of these catfishes are overfishing and alterations in and obstruction of river flow due to the construction of hydropower dams. After synthesizing existing information on species of the genus Pseudoplatystoma, we offer suggestions for future research to fill critical gaps in the knowledge of this group.
Key Contribution: This review synthesizes existing information on the ecology of species of the genus Pseudoplatystoma and offers suggestions for fisheries management and for research to fill critical gaps in the knowledge of the group.

1. Introduction
Freshwater fishes include diverse lineages collectively summing to over 18,000 species, representing ~51% of all fishes and ~25% of all vertebrates [1,2,3,4]. Freshwater fishes are important economically, culturally, aesthetically, scientifically, and educationally [5]. They provide the basis for human nutrition, employment, and ecosystem services, including nutrient transport within aquatic ecosystems, disease vector control, and seed dispersal [5,6]. About one-third of all freshwater species occur in the Neotropics, with an estimated 4475 species within 71 families [7], with new species being recognized regularly. This assemblage, however, faces a high level of anthropogenic pressure and is threatened by overexploitation, water pollution, river flow modification, degradation of habitat, and invasive exotic species [5,6,8]. Studies of the ecology of many Neotropical species are relatively scarce in the peer-reviewed literature, limiting the basis for science-based actions for fisheries management and conservation.
Catfishes of the Order Siluriformes include 39 families with 498 accepted genera and 4123 species [4]. More specifically, members of the Family Pimelodidae, the long-whiskered catfishes, and the genus Pseudoplatystoma are distributed from South America to southernmost Mexico [9]. They are a highly distinctive group of catfishes, with three pairs of barbels, maxillary barbels the length of their bodies, no scales, and well-developed adipose fins [9] (Figure 1). Many long-whiskered catfishes grow to be over 1 m in total length. The Family Pimelodidae includes 30 genera and 116 described species [4], and many valid pimelodid species have been described only recently [7]. Lundberg et al. [10] suggested that there are at least 25 undescribed living species, as well as five described and many undescribed fossil species.
Fishes of the genus Pseudoplatystoma support important commercial and artisanal fisheries, and some species have become important to regional aquaculture [11]. Most species of the genus are under pressure from dam construction and overfishing [12,13,14,15,16,17,18]. Two species are of conservation concern: P. corruscans is classified as Critically Endangered [13] and P. magdaleniatum as Endangered by IUCN [19]. Relatively few studies have focused on species of the genus Pseudoplatystoma, and further information on their biology, ecology, and population dynamics is needed to inform fisheries management and conservation. This review synthesizes existing information on the species of the genus Pseudoplatystoma. Six questions are asked: (i) What species are in the genus, and where are they distributed? (ii) What are the life histories and ecologies of Pseudoplatystoma species? (iii) What are the patterns of somatic growth for these species? (iv) What is known about the biomass production and population dynamics of Pseudoplatystoma? (v) What is the geographic distribution of genetic variation within Pseudoplatystoma species? (vi) What are the threats to the conservation of these species? After establishing what is known about the genus, this review offers suggestions for future research to fill critical gaps in the knowledge of this group.
2. What Species Are in the Genus, and Where Are They Distributed?
2.1. Natural History
An understanding of the natural history of the lineage provides insights into the geographic distribution of Pseudoplatystoma species. The first and so-far only reported fossil record for the genus is a neurocranium recently discovered from late Pleistocene deposits of the Cacarana River of Argentina, which was assigned to the extant species Pseudoplatystoma corruscans [20]. The fossil record of freshwater fishes in South America is not as well studied as that of terrestrial mammals [21]. Lundberg et al. [10] present a map showing sites where fossils from the Neogene period, spanning from the end of the Paleogene Period 23 million years ago (MYA) to the beginning of the present Quaternary Period 2.6 MYA, have been found, suggesting that additional finds might be made with effort directed there.
The systematics and taxonomy of the genus Pseudoplatystoma can be better understood in the context of the biogeological history of South America. Pseudoplatystoma has diverse species because extensive geomorphological and physiographic processes have transformed South American drainages over time, allowing vicariance, divergence, and subsequent secondary contact of lineages [22,23]. Fishes of the genus Pseudoplatystoma are currently distributed across four of the eight zoogeographic regions of South American freshwater fishes described by Géry [24], from north to south: the Magdalenian, Orinoco, Guinea-Amazonian, and Paranean regions. Géry [24] emphasizes that there are or have been connections between some South American river systems, some permanent (e.g., the Cassiquiera Canal joining the Rio Negro of the Amazon and the Orinoco drainages), some temporary (due to flooding in the rainy season), and some historical but no longer in existence. Current patterns of species distribution reflect these biogeographical processes, including marine incursions and the uplift of paleoarches, or ancient ridges [25].
2.2. Systematics and Taxonomy
The isolation of freshwater refugia permitted allopatric speciation, and subsequent recolonization of the lowland allowed the mixing of the respective lineages. Hubert and Renno [25] showed seven putative dispersal routes that gave rise to different species’ distributions. Against this background, it becomes apparent that P. magdaleniatum, P. orinocense, and P. mataense persisted in northern refugia, P. corruscans and P. reticulatum in southern refugia, and other Pseudoplatystoma species in western refugia. The model proposed by Hubert and Renno [25] could explain how four species’ distributions now overlap in the Amazon basin. Pseudoplatystoma species are not shared between the now-connected Orinoco and Amazon basins, which may be explained by the species not dispersing readily through small river systems.
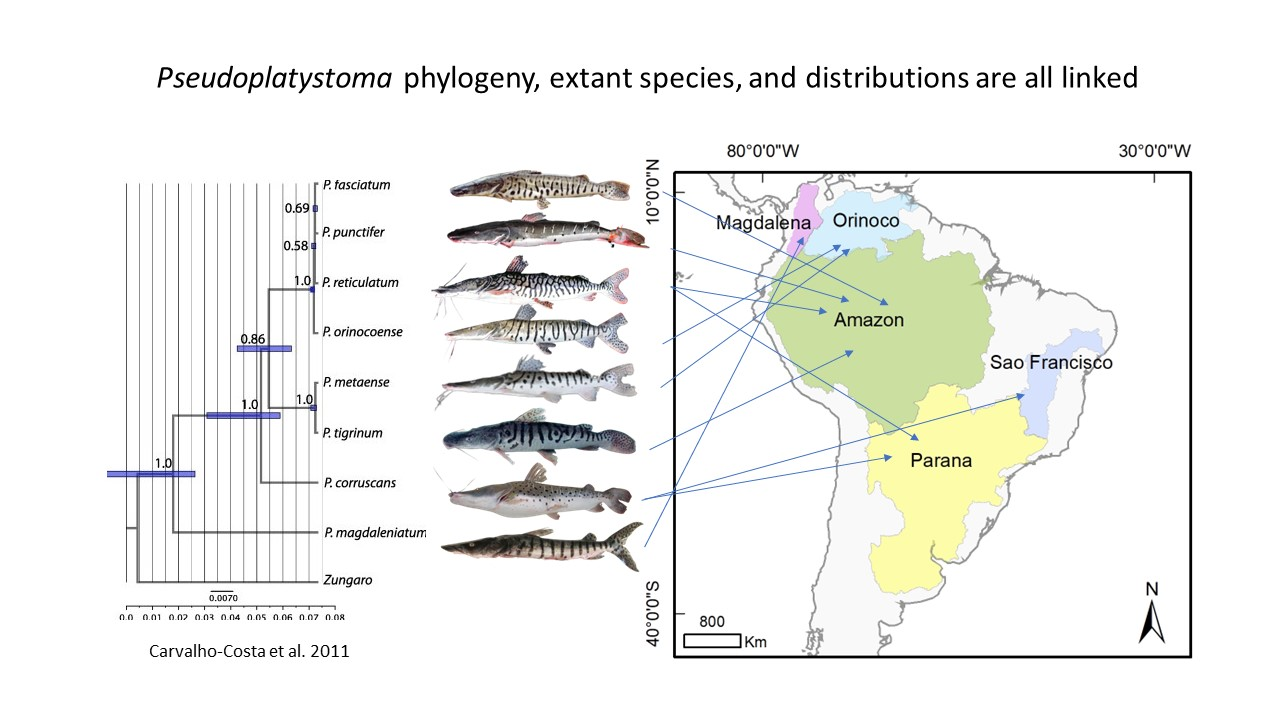
The phylogeny and distributions of species across the South American landmass are linked [10]. Until recently, three species of Pseudoplatystoma were recognized, P. fasciatum, P. tigrinum, and P. corruscans. Despite their apparent morphological homogeneity within species, the genus Pseudoplatystoma has cryptic species [26]. Based on morphological characteristics, Buitrago-Suárez and Burr [26] increased the number of known species to eight: P. punctifer and P. tigrinum in the Amazon basin, P. orinocense and P. mataense in the Orinoco basin, P. corruscans and P. reticulatum in the Paraná basin, P. magdaleniatum in the Magdalena basin, and P. fasciatum in the Guyana Shield. Later, using molecular genetic analyses, Torrico et al. [27] and Carvalho-Costa et al. [28] invalidated the distinction between P. fasciatum and P. punctifer made by Buitrago-Suárez and Burr [26]. Torrico et al. [27] evaluated species boundaries and biological and geographic patterns within the genus Pseudoplatystoma using a variation of mitochondrial cytochrome b and control region sequences to reconstruct phylogenetic relationships. They confirmed aspects of the morphological classification [26], but other elements were not supported. Morphology and molecular data supported the monophyly of the genus Pseudoplatystoma, i.e., their descent from a common ancestor. Molecular data also showed that P. tigrinum, P. corruscans, P. reticulatum, and P. magdaleniatum were highly supported clades, validating their taxonomic status. However, there were discrepancies between morphological and molecular genetic findings regarding P. mataense and P. orinocoense, suggesting either mtDNA introgression between the two species or misidentification of samples. The absence of phylogenetic and clear morphological differentiation between P. fasciatum and P. punctifer was such that Buitrago-Suárez and Burr [26] had erroneously separated P. fasciatum into two distinct species, thereby invalidating the taxonomic status of P. punctifer. On the other hand, Lundberg et al. [29] included four species in their study of the phylogenetics of Family Pimelodidae using nuclear and mitochondrial gene sequences, omitting the species P. punctifer. Moreover, the species P. fasciatum, P. tigrinum, P. magdaleniatum, and P. corruscans formed a well-supported, monophyletic clade embedded within the sorubimines, a lineage of ten genera. The position of P. tigrinum differed from that in the analysis of Buitrago-Suárez and Burr [26]. Moreover, García-Dávila et al. [30] showed the existence of a cryptic species within Pseudoplatystoma, with the new species resembling what Castelnau (1855) had named Platystoma punctifer, a species with no black stripes and a distinct mouth (Figure 1). While becoming clearer, details of the phylogeny of Pseudoplatystoma are yet undefined and will become more firmly established only with further study of suitably large collections of lineages and the application of suitable phylogenetic markers.
2.3. Distributions
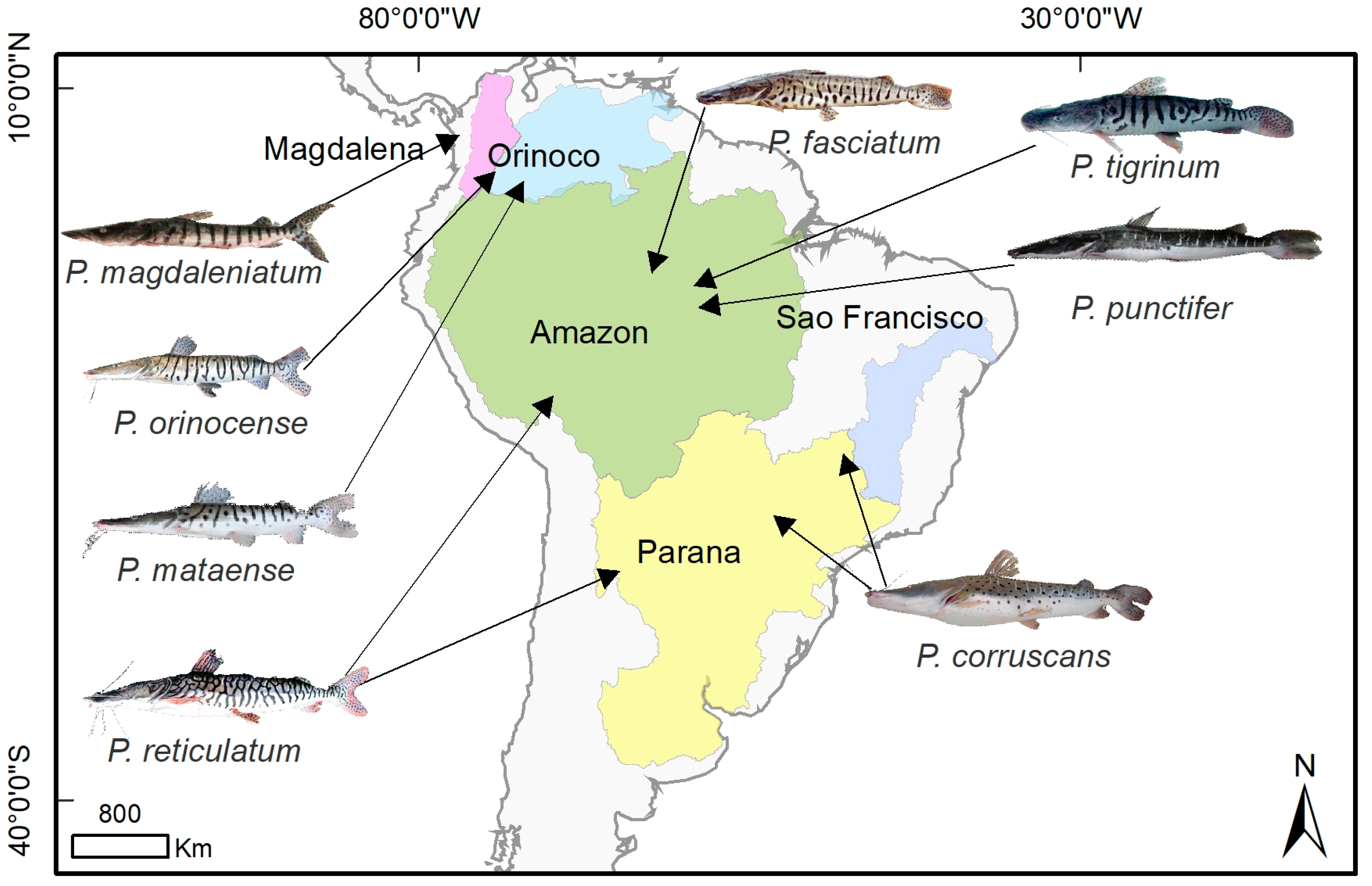
Developments in understanding the systematics of the genus Pseudoplatystoma have led to eight species being currently recognized in the taxonomy, and their distributions (Figure 2) are explained based on the natural history of South America. The Amazon basin contains three nominal species: P. tigrinum, P. fasciatum (named P. punctifer by Buitrago-Suárez et al. [27]), and P. reticulatum in the Amazon River, distributed in the mainstem Amazon and its Madeira, Solimões, Xingu, Tapajós, and Jurua tributaries. Additionally, there are the cryptic species believed to be what Castelnau (1855) described as P. punctifer: P. cf. punctifer [30]. Two species occur in the Orinoco basin: P. mataense, distributed in the Orinoco River in Venezuela and Colombia, and P. orinocoense in the Orinoco River in Venezuela. P. magdaleniatum is found in the Magdalena basin. P. fasciatum is found in the Guyana Shield in the Amazon, Corantijn, Essequibo, Orinoco, (Figure 1). Two species occur in the Paraná basin: P. corruscans and P. reticulatum in the central Amazon River and the Paraná River basin in Argentina, Bolivia, Brazil, Paraguay, and Uruguay. Only P. corruscans occurs in the São Francisco basin. However, because our understanding of the systematics of the respective species is still not fully defined, more studies are needed to clarify the systematics, refine the taxonomy, and explain the distributions of the respective lineages in the genus.
3. What Are the Life Histories and Ecologies of Pseudoplatystoma Species?
3.1. Movement Ecology
Most Pseudoplatystoma catfishes exhibit ontogenetic and feeding migrations, longitudinal migration for reproduction, and lateral migration for feeding [31,32]. However, the migration patterns of certain Pseudoplatystoma species need to be better understood, as there have been very few studies on the topic [32,33]. In the Colombian Amazon, it is believed that Pseudoplatystoma spp. migrate between 300–500 km [34], but no movement data are available. Patterns of movement in Pseudoplatystoma have been shown to vary by sex, size, time of year, and ability of fish to cross natural barriers such as waterfalls [18,31,33,35].
While there is only a limited understanding of the movement ecology of Pseudoplatystoma spp., especially regarding its potential importance in demographically and genetically structuring their populations, general patterns of migration of freshwater floodplains fishes suggest insights into species-level ecology of Pseudoplatystoma catfishes. The eggs and larvae of pimelodid catfishes have been found drifting in currents, in both white- and black-water rivers, and in both clear and turbid waters [36,37]. Catfish larvae drift mainly near the bottom of the water column, drifting distances of up to several thousand kilometers. Fast downstream movement (averaging 250 km/d) was suggested by Barthem et al. [38] for the early life stages of some pimelodids in rain-swollen currents.
Few studies have explicitly distinguished the movement ecology of sexes and life stages of Pseudoplatystoma catfishes. Male P. fasciatum and P. tigrinum usually inhabit rivers, while females inhabit riverside lakes during the low-water season [31]. Adults of both species migrate upstream for spawning during the beginning of the high-water season in the Mamore River in the Beni basin, Bolivia. At the same time, juvenile individuals believed to be non-migratory remain in the floodplains [31]. Adult P. fasciatum that possessed undeveloped gonads moved upstream and passed through river rapids during the low-water season in the Madeira River [35]; no fish were seen returning downstream. Juvenile P. fasciatum are always caught in the floodplains; other migratory behavior is unknown. Moreover, P. fasciatum and P. tigrinum migrate upstream across the rapids in the Madeira River, probably seeking prey. However, despite the incomplete and seemingly inconsistent information about the timing and the direction of migration in P. tigrinum and P. fasciatum [31,33], some studies have elucidated the migration of P. punctifer (P. fasciatum) and P. corruscans.
P. fasciatum performs movements both up and down river channels, and most movements occur between the dry season and the rising water period [39]. P. fasciatum individuals were tracked using telemetry methods in the Xingu River, where they migrated up to 164 km between the dry season and the rising water period. P. fasciatum was mainly detected moving upstream during the end of the rising-water period.
Migration and spawning patterns of P. corruscans in the São Francisco River were observed using radio telemetry [40]. Two migration patterns were exhibited, showing resident and migratory fish. According to the spawning stage, the migration was classified into pre-spawning staging and spawning migration. Female P. corruscans migrated among pre-spawning staging, spawning, and non-spawning sites. Godinho et al. [40] suggested that ripe females performed homing migration from feeding areas to the spawning sites, i.e., discrete sites where they returned multiple times. After spawning, most females migrated from the spawning site to other habitat units. The natural history of P. corruscans in the La Plata River in the Uruguay River basin was inferred using ratios of strontium isotopes (87Sr/86Sr) along a transect of the otolith [41]. P. corruscans was inferred to originate from the Paraná River, indicating a distinct nursery area. All P. corruscans individuals showed ontogenetic shifts in their isotopic signature, with strontium ratios corresponding to catchment sub-basins, suggesting that this species presents spatially bounded reproduction in the respective areas [41].
Given the lack of information about ontogenetic migration patterns for Pseudoplatystoma species, more studies are necessary to fully understand the life cycle of these species in the respective basins in which they are distributed. Questions that can and should be asked in future studies include, How far do the species migrate? Which habitats are used during their longitudinal and lateral migrations? Do juveniles and adults, and females and males, have the same migration patterns? Do all members of a species have the same migratory life history? Answering these questions would improve our understanding of the species’ life cycles and inform fisheries management and conservation for Pseudoplatystoma species.
3.2. Reproduction
The reproductive biology of Pseudoplatystoma species is not yet well studied. The current knowledge of the reproductive ecology of Pseudoplatystoma indicates that five of the eight species have common characteristics. Size-at-maturity varies among species between 57 cm to 82 cm, with all three species for which data are available spawning during the rising water season via total spawning, i.e., the release of all eggs in one batch (Table 1). Larvae of P. fasciatum were mostly observed drifting with currents during the end of the flood pulse, while larvae of P. tigrinum were detected flowing during the start of the flood pulse in the Peruvian Amazon [42].
Additionally, the reproductive morphology of Pseudoplatystoma is like that of other Siluriformes. Pseudoplatystoma species have double gonads, with elongated ovaries and testicles with digitiform projections all over their extent, features that are more evident during the reproductive period. The ovaries are covered by conjunctive and muscular tissues and blood vessels [43]. The gonads converge at the caudal region and join a duct. The female gonads are cistovaries, meaning that the lumen is connected to the oviduct, through which the oocytes are released into the water [44]. The male gonads are covered by conjunctive tissue that ramifies into septs filled with seminiferous tubules [44].
The reproduction of Pseudoplatystoma is closely related to their migratory ecology, as their reproduction is synchronized with the flood pulse and occurs during the period of rising and high waters [32,44,45,46,47,48] (Table 1). The synchronization of reproduction with the upstream migration during the period of rising water enables these species to spawn when the river’s water level is favorable for the small, semi-buoyant eggs and larvae to drift with the current until they reach the inundated floodplains, which are their nursery habitats [32,36]. Consequently, an important life-history characteristic of the genus is that its species do not seem to provide parental care, given knowledge about their biology and considering the egg size and number. Females exhibit total spawning of numerous ova and high fecundity, which compensate for high early mortality [31,49,50]; however, quantitative data about the fecundity of most of these species is lacking. Embryogenesis is rapid, lasting around 16 h at an average temperature of 23.0 ± 1.0 °C [51]. For P. fasciatum, the hatching takes place 14 h after fertilization at 27 °C [52].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Reproductive parameters of Pseudoplatystoma species.
| Species | Size at First Maturation (cm) | Spawning Season | Spawning Type | Oocyte Diameter (mm) | Ref. |
|---|---|---|---|---|---|
| P. corruscans | 62.92 | Rising water | Total | 0.9 | [32,48] |
| P. reticulatum | 57.84 | Rising water | Total | - | [48] |
| P. tigrinum | 69.4 | Rising water | Total | - | [45,53] |
| P. fasciatum | 53 | Rising water | Total | 0.64 | [31,53] |
| P.magdaleniatum | 82 | Rising water | - | 0.9 | [47] |
| P. tigrinum | 69.4 | High water | - | - | [42] |
| P. fasciatum | 53 | Rising water | - | 0.64 | [42] |
Current knowledge of the reproductive biology of some of the species allows managers and decision-makers to take action even in the absence of more detailed studies. Based on the current information for five species in the genus, the establishment of regulations for time and minimum size-at-capture for those species lacking information could be to restrict their fisheries during the rising and high-water periods. Managers could calculate a ratio L50/Lmax for the species with available information and recommend an average ratio for the others until new studies are done.
The reproductive biology of Pseudoplatystoma species is not yet well studied, so existing knowledge gaps represent opportunities for research. One question that remains unanswered in regard to their behavioral reproductive ecology is: What is the female behavior during spawning? Do females emit any pheromonal or behavioral signal prior to or during spawning? In addition to fecundity information available for P. fasciatum and P. reticulatum [54,55], another knowledge gap relates to the fecundity of these five species: what is the range of age- or size-specific fecundities? Additionally, knowledge of the characteristics of eggs and larvae is important for understanding the reproductive ecology of Pseudoplatystoma species. In particular, we must answer the questions: Do larvae perform vertical movement in the water column? What factors limit their recruitment to the juvenile stage? Are the respective species iteroparous or semelparous? If the former, how many spawning cycles occur during the lifetime? Do the species have broadcast spawning or other behaviors? Do juveniles have direct development or transformations? If the latter, how many inflection points? Answering these questions would enhance the current knowledge of reproductive biology, which is essential to managing fisheries and conserving these species.
3.3. Growth
The growth of six Pseudoplatystoma species has been studied. As noted above, these catfishes are large-bodied compared to other Neotropical species and can grow up to about 130 cm in total length and 100 kg in weight, depending on the species [56]. They can grow to 40 cm in their first year of life [14]. Females grow larger and faster than males. Von Bertalanffy growth parameters and Ø values for Pseudoplatystoma species are summarized in Table 2.
Most studies on the growth of Pseudoplatystoma found a similar pattern of growth rings in calcified structures such as otoliths and vertebrae, with opaque ring formation in the hard parts of Pseudoplatystoma, indicating fast growth occurring during the dry season [14,31,50,58,60]. Several studies showed that one opaque ring is formed annually during the dry season and the beginning of the rainy season for P. punctifer, P. tigrinum, and P. mataense in the Peruvian Amazon, Orinoco, and Mamore basins [14,31,57], probably because the dry season is when prey capture is more difficult due to competition with larger catfishes and dolphins and the beginning of the reproductive stage [14,31,33]. However, studies on central and eastern Amazon tributaries are necessary to determine whether the pattern of one ring per annual cycle is consistent for P. fasciatum and P. tigrinum. Other fish species—such as gilded catfish Brachyplatystoma rousseauxii (Castelnau, 1855) [58], mapara catfish Hypophthalmus marginatus (Valenciennes, 1840) [57], piaracatinga catfish Calophysus macropterus (Lichtenstein, 1819) [61], and pirarucu Arapaima spp. [59] in the central Amazon—form two growth rings per flood cycle [14]. Validation of ring formation has been done for P. punctifer, P. tigrinum, and P. fasciatum, but it is lacking for P. corruscans, P. reticulatum, and P. magdaleniatum [14,31]
Generally, species that are distributed in tropical and temperate regions have faster growth rates in the warmer tropical waters because food availability is the main constraint for growth in tropical regions, while temperature constrains the growth in temperate regions [62,63]. For example, all species of the genus Pseudoplatystoma that are distributed in the tropical areas of the Amazon, Orinoco, and Magdalena basins grow faster than the Pseudoplatystoma catfishes distributed in the Paraná basin (references in Table 2).
4. What Is Known about the Biomass Production and Population Dynamics of Pseudoplatystoma Species?
Biomass production and population dynamics of Pseudoplatystoma species have not been properly studied. However, current knowledge about their life-history and general patterns of biomass production and population dynamics of tropical species are expected to follow general patterns common to fishes in river floodplains [64,65]. This pattern involves seasonal changes in biomass via increases during high-water months due to active feeding and reproduction activities and decreases during low-water months due to natural mortality [66].
The population dynamics of Pseudoplatystoma species can also be understood based on the triangular model proposed by Winemiller and Rose [67], which classifies fishes in relation to their life-history trade-offs and the adaptative response to environmental variation. According to this classification, fishes can be classified within a continuum of three strategies: (1) periodic, in which fish have high values of both fecundity and age-of-maturity and a low value of juvenile survival; (2) opportunistic, in which fish have a low value of absolute fecundity but high productivity for their size, low age-at-maturity, and juvenile survival; and (3) equilibrium, in which fishes have low values of fecundity and high values in the age-at-maturity and juvenile survival (meaning that juveniles have relatively high survival rate, probably because of parental care). Winemiller [65] classified P. fasciatum as periodic because it has high absolute fecundity and intermediate size-at-maturity. The catfish has intermediate juvenile survival as it has planktonic eggs, and as soon as the yolk sac is reabsorbed, the larvae prey on items, including cannibalizing other larvae. P. fasciatum has a smaller size-at-maturity currently, compared to 20 to 30 years ago, mainly due to fishing which selectively removes the largest individuals from the populations, resulting in artificial selection for genotypes maturing at smaller sizes [68]. As periodic strategists, small differences in mortality at early life stages may have a huge influence on stock abundance [65].
Pseudoplatystoma species could be considered vulnerable to fishing pressures, as shown by fisheries stock assessments done for P. tigrinum and P. corruscans, which showed signs of growth of overfishing. Ruffino et al. [68] concluded that the stock of P. tigrinum was exploited very closely to maximum sustainable yields. Doria [67] showed that the fisheries landings of P. tigrinum are fewer and individual sizes are smaller than previously. Godinho [69] pointed out that some P. corruscans populations have been overfished to the point where they have disappeared.
5. What Is the Geographic Distribution of Genetic Variation within Pseudoplatystoma Species?
Maintaining the viability of valued populations is fundamental to fisheries management and conservation. Understanding of population’s genetic structure would inform the management and conservation of fishes of the genus Pseudoplatystoma. Studies identifying genetically distinct populations are needed to determine appropriate spatial units for managing demographically sustainable fisheries. In cases where genetically distinct gene pools could mix, as perhaps in large rivers, population genetic studies are needed to recognize that mixing and estimate the relative abundances of the respective stocks contributing to mixed-stock fisheries to protect the demographically less-robust populations. Genetic baseline information also is needed to design any hatchery-based programs that might be considered for supplementing wild fish populations [70]. A table of genetic concepts helpful for understanding key concepts of population genetics is presented in Table 3. While population genetic data are lacking for some species of this genus, a review of the existing literature shows that population genetic patterning varies among Pseudoplatystoma species.
Pereira et al. [71] investigated whether populations of P. corruscans were panmictic in the Paraná-Paraguay basin. They found highly significant population genetic differentiation among samples (FST = 0.034–0.163, AMOVA FST = 0.082%, and AMOVA RST = 0.097%). The patterning of individuals’ multilocus genotypes into inferred clusters using cluster analysis and Bayesian genotypic assignment was largely congruent with the branching topology of a population-level neighbor-joining phylogenetic tree. The results thus demonstrated genetic differentiation among populations. The documented long-distance movements of P. corruscans and the geographical scale of genetic differentiation found by Pereira et al. [71] result from a strong tendency for homing to natal nursery areas for reproduction [12]. This result supports the need for separate management of each spawning assemblage as an independent management unit, or MU. That is, fishery management must set sustainable exploitation rates for each MU, as each is demographically independent. We have no indication that these management strategies are being implemented for the fisheries.
Carvalho-Costa et al. [28] sampled P. corruscans at two sites in the São Francisco and the Paraná-Paraguay River basins in Brazil. They assessed the DNA sequence variation of a segment of the mitochondrial COI region and repeat-number variation at eight nuclear microsatellite DNA loci. Two mitochondrial DNA sequence haplotypes were detected in the São Francisco River basin and seven in the Paraná-Paraguay River basin. They found identical population tree topologies by neighbor-joining, maximum parsimony, and Bayesian analyses. They obtained high fixation index values between the São Francisco River basin and the Paraguay River basin for both nuclear and mitochondrial markers, i.e., FST of 0.18 (p = 0.01) and φST of 0.89 (p = 0.001). Several private microsatellite alleles were observed for each river basin. Bayesian cluster analysis of the entire dataset indicated the most likely number of genetic clusters to be K = 2, corresponding to river basins. Tajima’s D and Fu’s Fs indices both suggested population expansion of P. corruscans in the Paraguay River basin, the geologically recent river system. At the same time, no such indication was found for the São Francisco basin, the geologically older basin. Nuclear and matrilinear DNA divergence, combined with their historical separation into different hydrographical systems, strongly suggested that the two population groups evolved independently, perhaps warranting their recognition as distinct Evolutionary Significant Units, or ESUs. P. corruscans is considered abundant in the Pantanal wetlands region within the Paraguay River basin. Populations from the São Francisco River basin belong to a different ESU and hence should be managed as a separate unit. Some of these populations are critically affected by habitat fragmentation from several hydroelectric dams and overexploitation [13].
Hybrid P. fasciatum x P. corruscans are produced in aquaculture, and hybrids have been collected from the wild, presumably after escape from confinement. Dantas et al. [12] observed variation at five microsatellite DNA loci in three wild populations, and a captive group of P. corruscans was considered a potential source of broodstock for an augmentation program in the São Francisco River. FST and assignment test results showed low differentiation among three wild populations and the captive broodstock. The authors recommended that P. corruscans broodstock used in stocking-based augmentation programs for severely impacted river basins be genetically characterized for species purity before possible use as broodstock. Their results suggested that the captive population should not be used to stock areas where wild populations remain extant.
Telles et al. [72] used detailed spatial analyses to investigate the genetic variation of P. punctifer (P. fasciatum), which is widely distributed and commercially important in the Amazon River basin. They screened samples from fifteen localities along the Madeira and Solimoes rivers at seven microsatellite DNA loci. High genetic diversity was observed. Overall, FST was relatively low (0.057; p < 0.001), although pairwise FST values ranged from 0 to 0.21 among some pairs of populations. A Mantel correlogram of genetic distance as a function of geographic distance showed that populations up to 80 km apart tended to be more similar than expected by chance. A patch size of around 200 km is compatible with genetic isolation-by-distance and local constraints upon individual dispersal, resulting in spatially structured levels of gene flow. This pattern of genetic variation will inform strategies for maintaining genetic diversity in the species, especially considering threats due to the anthropogenic impacts of large dams constructed in this river system.
P. corruscans is the most-studied Pseudoplatystoma species in terms of population genetic structure. However, current knowledge of population genetic structure for catfishes of genus Pseudoplatystoma still needs to be more robust to inform spatially explicit fishery and conservation management. Hence, further studies are needed to understand the patterns of population genetic structuring of all Pseudoplatystoma species in their respective basins to delineate Management Units and possible Evolutionary Significant Units to manage fisheries and conserve these species properly. Over the longer term, the development of a large collection of single nucleotide polymorphism (SNP) markers could support the execution of genome-wide association studies to identify adaptive genetic variation. This knowledge would inform the conservation of the adaptive potential of the respective populations and species.
6. What Are the Threats to the Conservation of These Species?
The main threats to Pseudoplatystoma species are (1) overfishing and (2) alterations in and obstruction of river flow due to the construction of hydropower dams [73]. Other threats—such as industrial, domestic, and agricultural pollution, deforestation, introduced species, hybridization with farmed fish, and genetic erosion—may also affect populations of Pseudoplatystoma species, but in our view, they are not less pressing.
Dams interrupt streamflow and generate longitudinal and lateral hydrological changes in river ecosystems [74,75,76] that can be reflected subsequently in decreased fisheries yields. The most apparent effects of placing dams on rivers are the formation of a new lentic environment upstream from the dam and a tailwater environment downstream from the dam [75]. In general, six effects of dams on fisheries are (1) alteration of flood regime; (2) change in nutrient flow; (3) change in water temperature; (4) disruption of fish migration; (5) change in fish community composition; and (6) change of water chemistry [76]. For Pseudoplatystoma species, the alteration of the flood regime and disruption of migration are particularly important [32].
Alteration of the flood regime disrupts lateral connectivity, as dams cause the permanent inundation of upstream floodplains and restrict the seasonal movement of littoral zones that are part of the flood pulse [75]. Further, downstream of the dam, floodplains are not sufficiently inundated in terms of the depth, duration, and seasonality of flooding, which could result in a significant decline in Pseudoplatystoma populations and fisheries. That is, disruption of floodplain inundation is likely to affect the life cycle of Pseudoplatystoma, including disruption of migration, recruitment, and survival of cohorts. Dams blocking upstream and downstream migration can cause the decline and even extirpation of species that depend on longitudinal movements along the stream during certain phases of their life cycle [74,75]. Migratory fishes require different environments for the main stages of their life cycle: reproduction, juvenile production, growth, and sexual maturation (review in [73]). As a barrier to upstream migration, a dam prevents adults from reaching spawning grounds during the breeding season, resulting in recruitment failure and eventual extirpation of the population above the dam. Downstream migration past the dam may also be difficult or impossible for many fish species. Fish migrating into the reservoir from tributary streams may be unable to find their way to the dam and subsequently pass downstream through discharge structures. In particular, the modification of the downstream river-flow regime by an impoundment can have various adverse effects on Pseudoplatystoma fish species, including loss of stimuli for migration, loss of migration routes and spawning grounds, decreased survival of eggs and juveniles, decreased recruitment to the mature life stage and diminished fisheries production [76].
In addition to obstruction of river flow due to dams, overfishing is especially evident in parts of the São Francisco and Paraná rivers [32], particularly regarding P. corruscans stocks [32,77,78,79]. This species is subject to overfishing in Minas Gerais State, Brazil. P. corruscans harvest in the São Francisco River has manifested indications of collapse [79,80]. In the 1950s, the capture of thousands of P. corruscans with low fishery effort was the norm [81]. More recently, however, the yield of P. corruscans caught in a vital fishing area has declined from 10.3 kg fish/day in 1987 to 0.8 kg fish/day in 1999 [40].
In the Amazon basin, some authors have suggested that P. tigrinum and P. fasciatum are declining in mean catches and captured at small sizes [68,82,83,84]. Published landing data are low. Pseudoplatystoma species are essential catfishes in the landings of Amazonas State, Brazil. There were landings of 700 tons/y of P. fasciatum in seven fish-packing plants near Manaus in 1995 and 1996 [85]. In Tefe, Brazil, P. fasciatum landings averaged 18 tons/y between 1991–1994 [86]. In the Magdalena basin, the main threat to P. magdaleniatum is also overfishing. Estimates from landing indicate that the population there experienced a reduction of 72.7% between 1995 and 2007 [19]. The reasons for the decline are that the established minimum capture size and the ban on commercialization during the reproductive periods are not respected [87]. The species is not considered to reach maturity until sizes of 65 and 89 cm (total length) for males and females, respectively. Still, the average size-at-capture has declined from 92 cm in 1973 for the entire Magdalena basin to 42 cm in 2010 [88].
7. Conclusions
Although many essential studies have been made to understand the biology of Pseudoplatystoma species, many aspects require further investigation. In the absence of information, we have synthesized currently available knowledge to tentatively fill knowledge gaps for some species and to identify priority research topics to inform the sustainable fisheries management and conservation of Pseudoplatystoma species (Table 4).
To effectively manage Pseudoplatystoma species, knowledge of several key ecological aspects is still lacking (Table 4). The best-studied aspect of the ecology of all Pseudoplatystoma species regards the migration of all eight species. However, information on migration timing and distance, reproduction, and adult and juvenile growth is still missing for most species, which impedes fisheries management, including the delimiting of management units and fisheries areas, as well as the planning of dam construction. Based on the migratory life histories of P. fasciatum, P. tigrinum, P. punctifer, and P. magdaleniatum, we recommend avoiding the construction of dams, and when unavoidable, planning should include spatially explicit prioritization methods that trade-off electric power production and biodiversity loss under different scenarios in order to lower environmental costs [73].
Reproduction data is unavailable for P. punctifer, P. mataense, and P. orinocense, which challenges the development of effective management strategies for these species, including the delimitation of the period when fishing could be prohibited. In the absence of information on the reproduction of the species, we recommend that fishing be prohibited during the flood season for all species within their respective distributions.
Regarding growth, data is totally absent for P. orinocense and P. magdaleniatum. Moreover, validation of growth rings is missing for P. reticulatum, P. mataense, and P. corruscans. Growth is the main indicator of change in biomass which is essential to determine population production strategies for stock assessment and science-based management. Based on the growth information available for P. fasciatum, P. tigrinum, and P. punctifer, management strategies addressing the minimum size-at-capture should be developed and implemented.
Temporal data on changes in biomass and studies of the population dynamics of all Pseudoplatystoma species are few. This is the main ecological aspect impeding the management of the eight species of the genus. However, based on the current knowledge of general patterns of population dynamics of tropical species and the life-history traits of Pseudoplatystoma species, we suggest that recruitment of these species be classified as periodic. The biomass of tropical fishes can decrease by around 50% from the flood season to the dry season [63,64]. These considerations are such that harvest should be conservative, allowing populations to have a demographic buffer.
Finally, genetic data is missing for P. tigrinum, P. mataense, and P. orinocense. Therefore, without more studies on population genetics, the management of these species will be not well supported with data informing the delineation of management units. Until data are amassed and analyzed, we suggest that the management of Pseudoplatystoma species be done according to the basin in which the respective species are distributed. For instance, the management of P. corruscans in the Paraná-Uruguay basins should be done within the respective MUs. Fishery management must set sustainable exploitation rates for each MU, as each is demographically independent. Management of P. reticulatum and P. corruscans in the São Francisco and Paraná basins, respectively, should be done independently; these populations putatively belong to different MUs or even ESUs. Populations of P. corruscans in the Paraguay River basin are currently expanding, in contrast with those of P. reticulatum in the São Francisco River basin. Finally, in the Amazon basin, the management of P. fasciatum should be done within distributional patches that are separated from each other by at least 200 km. However, more studies are necessary to guide the management of P. fasciatum in the Amazon basin.
8. Future Directions: Knowledge Gaps
We have synthesized current understanding and identified the key knowledge gaps in interconnected areas of study of Pseudoplatystoma species. This body of information, complemented with future studies, would help amass the information needed for effective fisheries management and conservation of these species. Our understanding of the systematics and phylogeny of the genus Pseudoplatystoma is not yet settled. Further studies are needed to clarify the systematics, taxonomy, and distributions of the respective lineages in the genus. These studies will need suitably broad collections of study material from across the range of the species and well-chosen phylogenetic markers to address subtle patterns of evolutionary change. Historical interbreeding of lineages may complicate the inference of evolutionary history. In particular, the distinctions between P. fasciatum and P. punctifer and the introgression among P. mataense and P. mataense are of interest for both basic understanding and practical application in conservation and fishery management.
The growth of Pseudoplatystoma species has been better studied than other biological aspects. In general, previous studies indicate that the annulus formation in the hard parts (otoliths and vertebrae) of Pseudoplatystoma occurs during the dry season, with slower growth during the low-water season and faster growth during the high-water season. Secondly, we conclude that Pseudoplatystoma species have relatively slow growth rates and high longevity compared to many other Neotropical fish species. However, more studies about somatic growth plasticity and validation of annulus formation for all species are needed to understand patterns of growth of Pseudoplatystoma species throughout the distribution of the respective species.
The population dynamics of Pseudoplatystoma species have not yet been adequately described. Practical knowledge gaps regarding the population dynamics of the Pseudoplatystoma species still need to be filled. Population dynamics studies are missing for P. fasciatum, P. punctifer, P. reticulatum, P. magdaleniatum, and P. orinocense. However, available studies on sustainable fisheries landings suggest that most Pseudoplatystoma populations and species are overexploited. Some Pseudoplatystoma species seem to be imperiled, and more studies are necessary to understand the population dynamics and guide fisheries management actions to conserve these species.
Finally, regarding the population genetics of species within the genus Pseudoplatystoma, more studies of all eight species of the genus are needed. P. corruscans is the best-studied species of the genus. However, the same level of investigation should be applied to the other seven species. Ideally, the next steps in the study of molecular genetics of the genus should apply the same molecular markers so that results within and among species can be compared. The investigation of possible Management Units (MUs) and possible Evolutionary Significant Units (ESUs) is crucial for understanding the drivers shaping the population structure of these species and, thus, the appropriate planning for the fisheries management and conservation of the species.
Clearly, challenges in research of the biology of Pseudoplatystoma species persist. With continued research and collaboration among research groups, understanding the biology and ecology of this genus would fill these knowledge gaps and contribute to fisheries management and conservation of the eight species of the genus Pseudoplatystoma.
Author Contributions
Conceptualization, L.A.P. and L.C.; literature review, L.A.P.; maps and figures, L.A.P.; writing—original draft preparation, L.A.P.; writing—review and editing, L.A.P., E.M.H., L.C., D.J.O. and F.D.; supervision, E.M.H. and L.C.; funding acquisition, L.A.P., E.M.H. and L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NSF grant number 1852113 and Rufford Foundation, grant number 33099-1.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Gabriel Borba, Monalisa Silva, Fernanda Oliveira, and Mahatub Khan for their reading and contributions to the manuscript. Thank you to those who shared pictures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arthington, A.H.; Dulvy, N.K.; Gladstone, W.; Winfield, I.J. Fish Conservation in Freshwater and Marine Realms: Status, Threats and Management. Aquat. Conserv. 2016, 26, 838–857. [Google Scholar] [CrossRef] [Green Version]
- Pelayo-Villamil, P.; Astor Guisande, C.; Vari, R.P.; Manjarr Es-Hern Andez, A.; Garc Ia-Rosell, E.; Gonz Alez-Dacosta, J.; Urgen Heine, J.; Gonz Alez Vilas, L.; Patti, B.; Mar Ia Quinci, E.; et al. Global Diversity Patterns of Freshwater Fishes-Potential Victims of Their Own Success Diversity and Distributions. Divers. Distrib. 2015, 21, 345–356. [Google Scholar] [CrossRef]
- Pinder, A.C.; Britton, J.R.; Harrison, A.J.; Nautiyal, P.; Bower, S.D.; Cooke, S.J.; Lockett, S.; Everard, M.; Katwate, U.; Ranjeet, K.; et al. Mahseer. Fishes of the World: Status, Challenges and Opportunities for Conservation. Rev. Fish Biol. Fish. 2019, 29, 417–452. [Google Scholar] [CrossRef] [Green Version]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 16 May 2023).
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater Biodiversity: Importance, Threats, Status and Conservation Challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.J.; Paukert, C.; Hogan, Z. Endangered River Fish: Factors Hindering Conservation and Restoration. Endanger. Species Res. 2012, 17, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Foresti, A.F.; Hilsdorf, A.A.W.S. Genetics of Neotropical Fish: From Chromosomes to Populations. Fish. Physiol. Biochem. 2009, 35, 81–100. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Kumar, P.S.; Kabir, M.; Zuhara, F.T.; Mehjabin, A.; Tasannum, N.; Hoang, A.T.; Kabir, Z.; Mofijur, M. Threats, Challenges and Sustainable Conservation Strategies for Freshwater Biodiversity. Environ. Res. 2022, 214, 113808. [Google Scholar] [CrossRef]
- Nelson, J.S. Fishes of the World, 3rd ed.; J. Wiley and Sons: New York, NY, USA, 1994; ISBN 0471547131. [Google Scholar]
- Lundberg, J.G.; Pérez, M.H.S.; Dahdul, W.M.; Aguilera, O.A. The Amazonian Neogene Fish Fauna. In Amazonia, Landscape and Species Evolution: A Look into the Past; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 281–301. [Google Scholar] [CrossRef]
- Nuñez, J.; Dugué, R.; Corcuy Arana, N.; Duponchelle, F.; Renno, J.F.; Raynaud, T.; Hubert, N.; Legendre, M. Induced Breeding and Larval Rearing of Surubí, Pseudoplatystoma fasciatum (Linnaeus, 1766), from the Bolivian Amazon. Aquac. Res. 2008, 39, 764–776. [Google Scholar] [CrossRef]
- Dantas, H.L.; Arcanjo, M.; Neto, S.; Kelly, K.; Oliveira, C.; Severi, W.; Mendonça Diniz, F.; Raquel, M.; Coimbra, M.; Moura, M.R. Genetic Diversity of Captive and Wild Threatened Catfish Pseudoplatystoma corruscans in the São Francisco River. Rev. Fish. Sci. 2013, 21, 237–246. [Google Scholar] [CrossRef]
- Mello, P.H.; Venturieri, R.L.L.; Honji, R.M.; Moreira, R.G. Threatened Fishes of the World: Pseudoplatystoma corruscans (Agassiz, 1829) (Siluriformes: Pimelodidae). Environ. Biol. Fish. 2009, 85, 359–360. [Google Scholar] [CrossRef]
- Armas, M.; Ortega, H.; García-Vasquez, A.; García-Dávila, C.; Vargas, G.; Nuñez, J.; Renno, J.-F.; Duponchelle, F. Age Validation and Contrasted Growth Performances of Pseudoplatystoma punctifer (Siluriformes: Pimelodidae) in Two River Systems of the Western Amazon. Neotrop. Ichthyol. 2022, 20, e210099. [Google Scholar] [CrossRef]
- Santos, R.E.; Pinto-Coelho, R.M.; Fonseca, R.; Simões, N.R.; Zanchi, F.B. The Decline of Fisheries on the Madeira River, Brazil: The High Cost of the Hydroelectric Dams in the Amazon Basin. Fish. Manag. Ecol. 2018, 25, 380–391. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Pelicice, F.M.; Petry, A.C.; Gomes, L.C.; Júlio, H.F. Fish Diversity in the Upper Paraná River Basin: Habitats, Fisheries, Management and Conservation. Aquat. Ecosyst. Health Manag. 2007, 10, 174–186. [Google Scholar] [CrossRef]
- Peluso, L.M.; Mateus, L.; Penha, J.; Bailly, D.; Cassemiro, F.; Suárez, Y.; Fantin-Cruz, I.; Kashiwaqui, E.; Lemes, P. Climate Change Negative Effects on the Neotropical Fishery Resources May Be Exacerbated by Hydroelectric Dams. Sci. Total Environ. 2022, 828, 154485. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Machado, A.S.; Farias, I.P.; Hrbek, T.; Lizarazo, M.D.E.; Alves-Gomes, J.A.; Formiga, K.; Da Silva Batista, J. Genetic Differentiation and Gene Flow of the Amazonian Catfish Pseudoplatystoma punctifer across the Madeira River Rapids Prior to the Construction of Hydroelectric Dams. Hydrobiologia 2022, 849, 29–46. [Google Scholar] [CrossRef]
- Mojica, J.; Valderrama, M.; Jimenez-Segura, L.; Alonso, J.C. Pseudoplatystoma magdaleniatum. IUCN Red List Threatened Species 2016, e.T58439165A61474168. [Google Scholar] [CrossRef]
- Brambilla, L.; Toledo, M.J.; Ibarra, D.A. First Fossil Record of Pseudoplatystoma corruscans (Siluriformes, Pimelodidae) from the Late Pleistocene, Santa Fe, Argentina. J. S. Am. Earth Sci. 2021, 105, 102987. [Google Scholar] [CrossRef]
- Vallone, E.; Ignacio Vezzosi, R.; Luis Cione, A. First Fossil Fish (Teleostei, Siluriformes) from the Late Pleistocene of Santa Fe Province, Argentina Mammal Paleoneurology View Project Cranio-Appendicular Anatomy of American Fossil and Living Deers (Mammalia, Cervidae): Systematic and Phylogenetic Implications of the Most Representative Taxa from the Litoral Region View Project. Alcheringa Austral J. Palaeontol. 2017, 41, 369–377. [Google Scholar] [CrossRef]
- Hilsdorf, A.W.S.; Hallerman, E.M. Genetic Resources of Freshwater Neotropical Fishes. In Genetic Resources of Neotropical Fishes; Springer International Publishing: Cham, Switzerland, 2017; pp. 119–210. [Google Scholar]
- Cassemiro, F.A.S.; Albert, J.S.; Antonelli, A.; Menegotto, A.; Wüest, R.O.; Cerezer, F.; Coelho, M.T.P.; Reis, R.E.; Tan, M.; Tagliacollo, V.; et al. Landscape Dynamics and Diversification of the Megadiverse South American Freshwater Fish Fauna. Proc. Natl. Acad. Sci. USA 2023, 120, e2211974120. [Google Scholar] [CrossRef]
- Géry, J. The Fresh-Water Fishes of South. Am. Biogeogr. Ecol. S. Am. 1969, 2, 828–848. [Google Scholar]
- Hubert, N.; Renno, J.F. Historical Biogeography of South American Freshwater Fishes. J. Biogeogr. 2006, 33, 1414–1436. [Google Scholar] [CrossRef]
- Buitrago-Suárez, U.A.; Burr, B.M. Taxonomy of the Catfish Genus Pseudoplatystoma Bleeker (Siluriformes: Pimelodidae) with Recognition of Eight Species. Zootaxa 2007, 1512, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Torrico, J.P.; Hubert, N.; Desmarais, E.; Duponchelle, F.; Nuñez Rodriguez, J.; Montoya-Burgos, J.; Garcia Davila, C.; Carvajal-Vallejos, F.M.; Grajales, A.A.; Bonhomme, F.; et al. Molecular Phylogeny of the Genus Pseudoplatystoma (Bleeker, 1862): Biogeographic and Evolutionary Implications. Mol. Phylogenet. Evol. 2009, 51, 588–594. [Google Scholar] [CrossRef]
- Carvalho-Costa, L.F.; Piorski, N.M.; Willis, S.C.; Galetti, P.M.; Ortí, G. Molecular Systematics of the Neotropical Shovelnose Catfish Genus Pseudoplatystoma Bleeker 1862 Based on Nuclear and MtDNA Markers. Mol. Phylogenet. Evol. 2011, 59, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.G.; Sullivan, J.P.; Hardman, M. Phylogenetics of the South American Catfish Family Pimelodidae (Teleostei: Siluriformes) Using Nuclear and Mitochondrial Gene Sequences. Proc. Acad. Nat. Sci. Phila. 2011, 161, 153–189. [Google Scholar] [CrossRef]
- García-Dávila, C.; Duponchelle, F.; Castro-Ruiz, D.; Villacorta, J.; Quérouil, S.; Chota-Macuyama, W.; Nú, J.; Römer, U.; Carvajal-Vallejos, F.; Renno, J.-F.; et al. Molecular Identification of a Cryptic Species in the Amazonian Predatory Catfish Genus Pseudoplatystoma (Bleeker, 1962) from Peru. Genetica 2013, 141, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Loubens, G.; Panfili, J. Biologie de Pseudoplatystoma fasciatum et P. tigrinum (Teleostei: Pimelodidae) Dans Le Bassin Du Mamoré (Amazonie Bolivienne). Ichthyol. Explor. Freshw. 2000, 11, 13–34. [Google Scholar]
- Carolsfeld, J.; Harvey, B.; Ross, C.; Baer, A. (Eds.) Migratory Fishes of South. America; World Fisheries Trust: Victoria, BC, USA, 2003; ISBN 9780444536433. [Google Scholar]
- Barthem, R.; Goulding, M. Os Bagres Balizadores: Ecologia, Migração e Conservação de Peixes Amazônicos; Tefé: Sociedade Civil Mamirauá; CNPq: Tefe, Brazil, 1997. [Google Scholar]
- Diaz-Sarmineto, J.A.; Alvarez-León, R. Migratory Fishes of the Colombian Amazon. In Migratory Fishes of South America Chapter 7; Carolsfeld, J., Harvey, B., Ross, C., Baer, A., Eds.; World Fisheries Trust: Victoria, BC, USA, 2003; pp. 303–344. [Google Scholar]
- Goulding, M.; Lowe-McConnell, R.H. (Eds.) The Fishes and the Forest; University of California Press: Berkeley, CA, USA, 1980. [Google Scholar]
- Araujo-Lima, C.A.R.M.; Oliveira, E.C. Transport of Larval Fish in the Amazon. J. Fish. Biol. 1998, 53, 297–306. [Google Scholar] [CrossRef]
- Pavlov, S.; Nezdoliy, V.; Urteaga, A.; Sanches, O. Downstream Migration of Juvenile Fishes in the Rivers. J. Ichthyol./Vopr. Ikhtiologii 1995, 35, 227–248. [Google Scholar]
- Barthem, R.B.; Goulding, M.; Leite, R.G.; Cañas, C.; Forsberg, B.; Venticinque, E.; Petry, P.; Ribeiro, M.L.D.B.; Chuctaya, J.; Mercado, A. Goliath Catfish Spawning in the Far Western Amazon Confirmed by the Distribution of Mature Adults, Drifting Larvae and Migrating Juveniles. Sci. Rep. 2017, 7, 41784. [Google Scholar] [CrossRef] [Green Version]
- Hahn, L.; Martins, E.G.; Nunes, L.D.; da Câmara, L.F.; Machado, L.S.; Garrone-Neto, D. Biotelemetry Reveals Migratory Behaviour of Large Catfish in the Xingu River, Eastern Amazon. Sci. Rep. 2019, 9, 8464. [Google Scholar] [CrossRef] [Green Version]
- Godinho, A.L.; Kynard, B.; Godinho, H.P. Migration and Spawning of Female Surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environ. Biol. Fish. 2007, 80, 421–433. [Google Scholar] [CrossRef]
- Avigliano, E.; Pouilly, M.; Bouchez, J.; Domanico, A.; Sánchez, S.; Llamazares Vegh, S.; Clavijo, C.; Scarabotti, P.; Facetti, J.F.; Caffetti, J.D.; et al. Strontium Isotopes (87Sr/86Sr) Reveal the Life History of Freshwater Migratory Fishes in the La Plata Basin. River Res. Appl. 2020, 36, 1985–2000. [Google Scholar] [CrossRef]
- Mariac, C.; Renno, J.-F.; Garcia-Davila, C.; Vigouroux, Y.; Mejia, E.; Angulo, C.; Ruiz, D.C.; Estivals, G.; Nolorbe, C.; Vasquez, A.G.; et al. Species-Level Ichthyoplankton Dynamics for 97 Fishes in Two Major River Basins of the Amazon Using Quantitative Metabarcoding. Mol. Ecol. 2021, 31, 1627–1648. [Google Scholar] [CrossRef] [PubMed]
- Crepaldi, D.V.; Faria, P.M.C.; Teixeira, E.D.A.; Ribeiro, L.P.; Augusto, Â.; Costa, P.; Chemim De Melo, D.; Paula, A.; Cintra, R.; Prado, S.D.A.; et al. Estructura Histologica de los ovarios de Pseudoplatystoma corruscans (Agassiz, 1829) Pimelodidae, Siluriformes. Boletim do Instituto de Pesca 1996, 23, 203–212. [Google Scholar]
- Brito, M.F.G.; Bazzoli, N. Reproduction of the Surubim Catfish (Pisces, Pimelodidae) in the São Francisco River, Pirapora Region, Minas Gerais, Brazil. Arq. Bras. Med. Vet. Zootec. 2003, 55, 624–633. [Google Scholar] [CrossRef]
- Pérez, A.; Castillo, O.; Barbarino, A.; Fabré, N. Aspectos Reproductivos Del Bagre Rayado Pseudoplatystoma tigrinum (Siluriformes, Pimelodidae) En La Cuenca Del Río Apure, Venezuela. Zootecnia Trop. 2012, 30, 251–262. [Google Scholar]
- Jiménez-Segura, L.F.; Palacio, J.; López, R. Caracteristicas Biologicas del Blanquillo Sorobim cuspicaudas Littmann, Burr y Nass, 2000 y Bagre rayado Pseudoplatystoma magdaleniatum Buitrago-Suarez y Burr 2007 (Siluriformes: Pimelodidae) Relacionadas con su Reproducction en la Cuenca Media del Rio Magdalena, Colombia. Actual. Biol. 2009, 31, 53–66. [Google Scholar]
- Barzotto, E.; Oliveira, M.; Mateus, L. Reproductive Biology of Pseudoplatystoma corruscans (Spix and Agassiz, 1829) and Pseudoplatystoma reticulatum (Eigenmann and Eigenmann, 1889), Two Species of Fisheries Importance in the Cuiabá River Basin, Brazil. J. Appl. Ichthyol. 2017, 33, 29–36. [Google Scholar] [CrossRef]
- Rizzo, E.; Godinho, H.P. Superfície de Ovos de Peixes Characiformes e Siluriformes. In Águas, Peixes e Pescadores do São Francisco das Minas Gerais; Godinho, H.P., Godinho, A.L., Eds.; PUC Minas: Belo Horizonte, Brazil, 2003; pp. 115–132. [Google Scholar]
- Reid, J. La Biología de Los Bagres Rayados Pseudoplatystoma fasciatum y P. tigrinum En La Cuenca Del Río Apure, Venezuela. Revista Unellez de Ciencia y Tecnología 1983, 1.1, 13–41. [Google Scholar]
- Resende, A.; Kawakami, E.; Catella, C.; Lima, F.; Shirley Da, N.; Palmeira, S.; Aparecida, R.; Pereira, C.; De, M.; Lima, S. Biologia Do Curimbatá (Prochilodus lineatus), Pintado (Pseudoplatystoma corruscans) e Cachara (Pseudoplatystoma fasciatum) Na Bacia Hidrográfica Do Rio Miranda. Infoteca.Cnptia.Embrapa.br, 1995; 75p, (EMBRAPA-CPAP. Boletim de Pesquisa, 02). [Google Scholar]
- Padilla Pérez, P.P.; Alcántara Bocanegra, F.; Orbe, R.I. Reproducción Inducida de La Doncella Pseudoplatystoma fasciatum y Desarrollo Embrionario-Larval. Folia amazónica 2001, 12, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Garcia, V.A.; Montreuil, V. Rodriguez R Talla de Primera Maduracion y Epoca de Desove de La “Doncella”, Pseudoplatystoma fasciatum (Linnaeus), y Del “Tigre Zungaro”, Pseudoplatystoma tigrinum (Valenciennes), En La Amazonia Peruana. Boletin del Museo Para Emilio Goeldi Séria Zoologia 2001, 17, 3–13. [Google Scholar]
- Núñez, J.; Castro, D.; Fernández, C.; Dugué, R.; Chu-Koo, F.; Duponchelle, F.; García, C.; Renno, J.F. Hatching Rate and Larval Growth Variations in Pseudoplatystoma punctifer: Maternal and Paternal Effects. Aquac. Res. 2011, 42, 764–775. [Google Scholar] [CrossRef]
- Andrade, F.F.; Lima, A.F.; Assumpção, L.; Makrakis, S.; Kasai, R.I.D.; Makrakis, M.C. Characterization of the Early Development of Pseudoplatystoma reticulatum Eigenmann & Eigenmann, 1889 (Siluriformes: Pimelodidae) from the Paraguay River Basin. Neotrop. Ichthyol. 2016, 14. [Google Scholar] [CrossRef] [Green Version]
- Erard, C.; Keith, P.; Le Bail, P.Y.; Planquette, P. Atlas Des Poissons d’eau Douce de Guyane. Revue d’Écologie (La Terre et La Vie) 2002, 57, 185–187. [Google Scholar]
- Pérez, A.; Fabré, N.N. Life-History Characteristics of Pseudoplatystoma Metaense (Teleostei: Siluriformes: Pimelodidae) from the Northwestern Orinoco River Basin. Neotrop. Ichthyol. 2018, 16, 160140. [Google Scholar] [CrossRef] [Green Version]
- Cutrim, L.; Batista, V.d.S. Determinação de Idade e Crescimento Do Mapará (Hypophthalmus marginatus) Na Amazônia Central. Acta Amazon. 2005, 35, 85–92. [Google Scholar] [CrossRef]
- Hauser, M.; Doria, C.R.C.; Melo, L.R.C.; Santos, A.R.; Ayala, D.M.; Nogueira, L.D.; Amadio, S.; Fabré, N.; Torrente-Vilara, G.; García-Vásquez, Á.; et al. Age and Growth of the Amazonian Migratory Catfish Brachyplatystoma russeauxii in the Madeira River Basin before the Construction of Dams. Neotrop. Ichthyol. 2018, 16, 1127. [Google Scholar] [CrossRef] [Green Version]
- Arantes, C.C.; Castello, L.; Stewart, D.J.; Cetra, M.; Queiroz, H.L. Population Density, Growth and Reproduction of Arapaima in an Amazonian River-Floodplain. Ecol. Freshw. Fish. 2010, 19, 455–465. [Google Scholar] [CrossRef]
- González, S.Á.R.; Mendoza, J.; Arocha, F.; Márquez, A. Edad y Crecimiento Del Bagre Rayado Pseudoplatystoma orinocoense Del Orinoco Medio En Venezuela. Zootec. Trop. 2010, 28, 283–293. [Google Scholar]
- Pérez, A.; Fabré, N.N. Seasonal Growth and Life History of the Catfish Calophysus macropterus (Lichtenstein, 1819) (Siluriformes: Pimelodidae) from the Amazon Floodplain. J. Appl. Ichthyol. 2009, 25, 343–349. [Google Scholar] [CrossRef]
- Henderson, P.A. The Growth of Tropical Fishes. Fish. Physiol. 2005, 21, 85–100. [Google Scholar] [CrossRef]
- Lagler, K.F.; Kapetsky, J.M.; Stewart, D.J. The Fisheries of the Kafue River Flats, Zambia, in Relation to the Kafue Gorge Dam; University of Michigan, School of Natural Resources: Ann Arbor, MI, USA, 1971. [Google Scholar]
- Welcomme, R.L. River Fisheries; Food and Agriculture Organization of the United Nations: Rome, Italy, 1985; ISBN 9251022992. [Google Scholar]
- Winemiller, K.O. Patterns of Variation in Life History among South. American Fishes in Seasonal Environments. Oecologia 1989, 81, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Winemiller, K.O.; Rose, K.A. Patterns of Life-History Diversification in North American Fishes: Implications for Population Regulation. Canad J. Fish. Aquat. Sci. 1992, 49, 2196–2218. [Google Scholar] [CrossRef]
- Doria, C.R.C.; Duponchelle, F.; Lima, M.A.L.; Garcia, A.; Carvajal-Vallejos, F.M.; Méndez, C.C.; Catarino, M.F.; Freitas, C.E.D.C.; Vega, B.; Miranda-Chumacero, G.; et al. Review of Fisheries Resource Use and Status in the Madeira River Basin (Brazil, Bolivia, and Peru) Before Hydroelectric Dam Completion. Rev. Fish. Sci. Aquacult 2018, 26, 494–514. [Google Scholar] [CrossRef]
- Ruffino, M.L.; Isaac, V.J. Dinamica Populacional de Surubim-Tigre, Pseudoplatystoma. Acta Amazon. 1999, 29, 463–476. [Google Scholar] [CrossRef]
- Godinho, H.P.; Miranda, M.O.T.; Godinho, A.L.; Santos, J.E. Pesca e Biologia Do Surubim Pseudoplatystoma corruscans No Rio São Francisco. In Colecão Meio Ambiente. Série de Estudos: Pesca; Miranda, M.O.T., Ed.; Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis: Belo Horizonte, Brazil, 1997; Volume 19, pp. 27–42. [Google Scholar]
- Fullerton, A.H.; Burnett, K.M.; Steel, E.A.; Flitcroft, R.L.; Pess, G.R.; Feist, B.E.; Torgersen, C.E.; Miller, D.J.; Sanderson, B.L. Hydrological Connectivity for Riverine Fish: Measurement Challenges and Research Opportunities. Freshw. Biol. 2010, 55, 2215–2237. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.H.G.; Foresti, F.; Oliveira, C. Genetic Structure of the Migratory Catfish Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) Suggests Homing Behaviour. Ecol. Freshw. Fish. 2009, 18, 215–225. [Google Scholar] [CrossRef]
- Telles, M.P.C.; Collevatti, R.G.; Braga, R.S.; Guedes, L.B.S.; Castro, T.G.; Costa, M.C.; Silva-Júnior, N.J.; Barthem, R.B.; Diniz-Filho, J.A.F. Geographical Genetics of Pseudoplatystoma punctifer (Castelnau, 1855) (Siluriformes, Pimelodidae) in the Amazon Basin. Genet. Mol. Res. 2014, 13, 3656–3666. [Google Scholar] [CrossRef]
- Duponchelle, F.; Isaac, V.J.; Doria, C.; Van Damme, P.A.; Herrera-R, G.A.; Anderson, E.P.; Cruz, R.E.A.; Hauser, M.; Hermann, T.W.; Agudelo, E.; et al. Conservation of Migratory Fishes in the Amazon Basin. Aquat. Conserv. 2021, 31, 1087–1105. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Gomes, L.C. Dams and the Fish Fauna of the Neotropical Region: Impacts and Management Related to Diversity and Fisheries. Braz. J. Biol. 2008, 68, 1119–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poff, L.N.; Hart, D.D. How Dams Vary and Why It Matters for the Emerging Science of Dam Removal. Bioscience 2002, 52, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.; Marmulla, G. Dams, Fish and Fisheries. Opportunities, Challenges and Conflict Resolution. FAO Fish. Tech. Paper 2001, 419, 166. [Google Scholar]
- Sato, Y. Reprodução de Peixes Da Bacia Do Rio São Francisco: Indução e Caracterização de Padrões; 179f. Tese (Doutorado); Centro de Ciências Biológicas e da Saúde, Universidade Federal de São Carlos: São Carlos, SP, Brazil, 1999. [Google Scholar]
- Sato, Y.; Godinho, H.P. Migratory Fishes of the São Francisco River. In Migrotory Fish of South America; Carolsfeld, J., Harvey, B., Ross, C., Eds.; World Fisheries Trust: Victoria, BC, USA, 2003; pp. 195–232. [Google Scholar]
- Godinho, H.P.; Miranda, M.O.T.; Godinho, A.L.; Santos, J.E. Pesca e Biologia Do Surubim Pseudoplatystoma Coruscans No Rio São Francisco; Puc Minas: Belo Horizonte, Brazil, 1997. [Google Scholar]
- Godinho, A.L.; Godinho, H.P. Breve Visão Do São Francisco. Belo Horizonte: PUC Minas 468 (2003): 15-23. In Águas, Peixes e Pescadores do São Francisco das Minas Gerais; PUC Minas: Belo Horizonte, Brazil, 2003; Volume 468, pp. 15–23. [Google Scholar]
- Menezes, R.S. Pesca e Piscicultura No Vale Do São Francisco. Boletim da Secretaria da Agricultura Indústria e Comércio do Estado de Pernambuco 1956, 23, 43–105. [Google Scholar]
- Isaac, V.; Ruffino, M.; McGrath, D. In Search of a New Approach to Fisheries Management in the Middle Amazon Region. In Fishery Stock Assessment Models; Alaska Sea Grant, University of Alaska Fairbanks: Fairbanks, AK, USA, 1998; pp. 889–902. [Google Scholar]
- Castello, L.; McGrath, D.G.; Beck, P.S.A. Resource Sustainability in Small-Scale Fisheries in the Lower Amazon Floodplains. Fish. Res. 2011, 110, 356–364. [Google Scholar] [CrossRef]
- Agudelo, E.A.; Acosta-Santos, G.; Gómez, B.D.; Gil, R.E.; Ajiaco-Martínez; Ramírez-Gil, H. Pseudoplatystoma punctifer (Siluriformes, Pimelodidae). In I. Catálogo de los Recursos Pesqueros Continentales de Colombia. Serie Editorial Recursos Hidro-biológicos y Pesqueros continentales de Colombia; Lasso, C.A., Agudelo Córdoba, E., Jiménez-Segura, L.F., Ramírez-Gil, H., Morales-Betancourt, M., Ajiaco-Martínez, R.E., Gutiérrez, F.d.P., Usma Oviedo, J.S., Muñoz Torres, S.E., Sanabria Ochoa, A.I., Eds.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH): Bogotá, Colombia, 2011; Capitulo 7; pp. 509–512. [Google Scholar]
- Araujo-Lima; Carlos, A.; Ruffino, M.L. Migratory Fishes of the Brazilian Amazon. In Migratory Fishes of South America; Carolsfeld, J., Harvey, B., Ross, C., Ross, C., Eds.; World Fisheries Trust: Victoria, BC, USA, 2003; pp. 233–301. [Google Scholar]
- Barthem, R.B. Várzea: Diversity, Development, and Conservation of Amazonia’s Whitewater Floodplains. Adv. Eco Bot. 1999, 13, 7–28. [Google Scholar]
- Lasso, C.A.; Morales-Betancourt, M.A. (Eds.) Catálogo de Los. Recursos Pesqueros Continentales de Colombia; Ministerio de Ambiente, Vivienda y Desarrollo Territorial-Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogota, Colombia, 2011.
- Valderrama, M.; Zarate, M.; Vera, G.; Moreno, C.; Caraballo, P.; Martinez, J. Determinación de La Talla Media de Madurez y Análisis de La Problemática Con Referencia a Las Tallas Medias de Captura Del Bagre Rayado (Pseudoplatystoma fasciatum) Linnaeus 1766 (Pisces: Pimelodidae), En La Cuenca Del Río Magdalena, Colombia. Trianea. Act. Cient. Tec.-INDERENA 1988, 2, 537–550. [Google Scholar]
Figure 1.
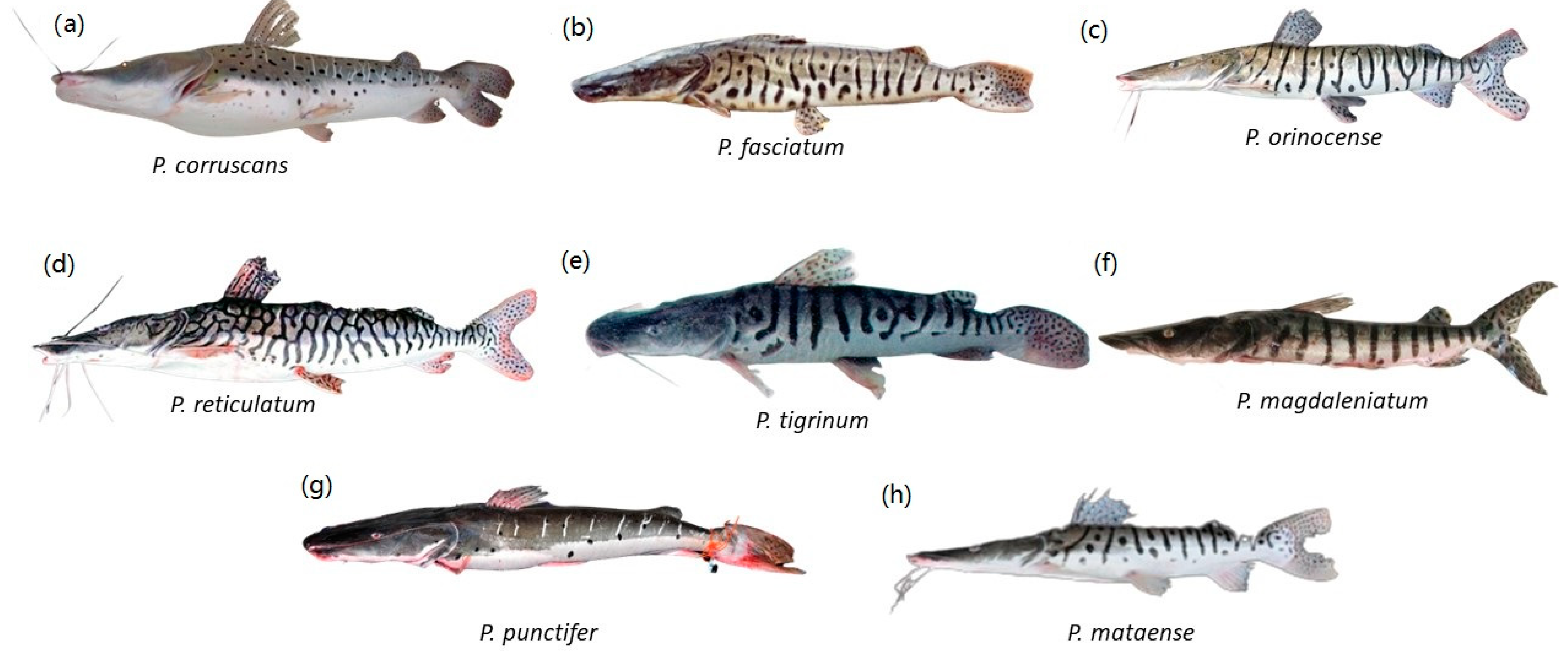
Eight Pseudoplatystoma species, members of the Family Pimelodidae; (a) P. corruscans; (b) P. fasciatum; (c) P. orinocense; (d) P. reticulatum; (e) P. tigrinum; (f) P. magdaleniatum; (g) P. punctifer; (h) P. mataense. Illustrations courtesy of Timm, C.D.; Magalhães, K.; Alvarez, F.; Sabaj Pérez; and F. Duponchelle.
Figure 1.
Eight Pseudoplatystoma species, members of the Family Pimelodidae; (a) P. corruscans; (b) P. fasciatum; (c) P. orinocense; (d) P. reticulatum; (e) P. tigrinum; (f) P. magdaleniatum; (g) P. punctifer; (h) P. mataense. Illustrations courtesy of Timm, C.D.; Magalhães, K.; Alvarez, F.; Sabaj Pérez; and F. Duponchelle.
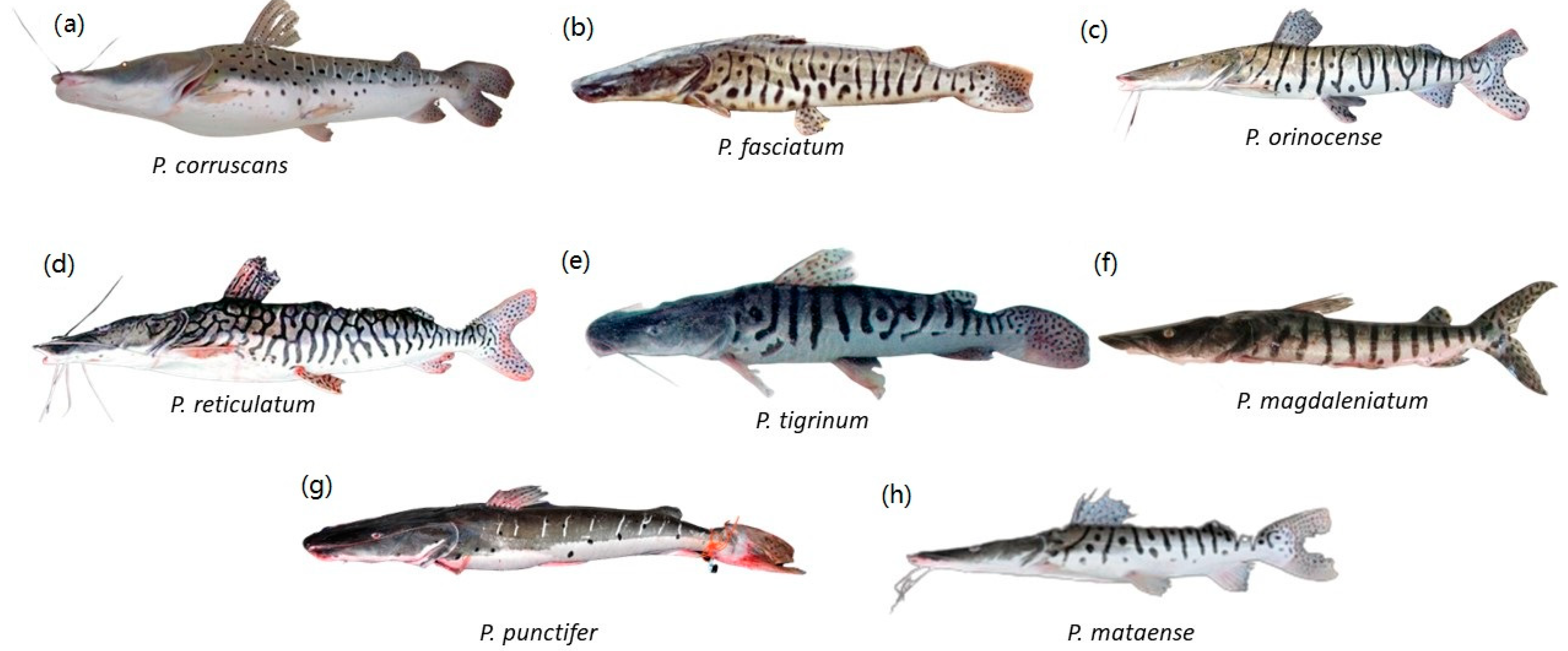
Figure 2.
Distributions of species of genus Pseudoplatystoma from the literature review data. The Magdalena basin is (represented in purple) where P. magdaleniatum is distributed. The Orinoco basin (light blue) encompasses the P. orinocense distribution. The Amazon basin (green) where P. fasciatum, P. tigrinum, P. reticulatum, and P. punctifer are distributed. P. corruscans is distributed in the São Francisco basin (blue), and P. corruscans and P. reticulatum are distributed in the Paraná basin (yellow). Illustrations courtesy of Timm, C.D.; Magalhães, K.; Alvarez, F.; Sabaj Pérez, and F. Duponchelle.
Figure 2.
Distributions of species of genus Pseudoplatystoma from the literature review data. The Magdalena basin is (represented in purple) where P. magdaleniatum is distributed. The Orinoco basin (light blue) encompasses the P. orinocense distribution. The Amazon basin (green) where P. fasciatum, P. tigrinum, P. reticulatum, and P. punctifer are distributed. P. corruscans is distributed in the São Francisco basin (blue), and P. corruscans and P. reticulatum are distributed in the Paraná basin (yellow). Illustrations courtesy of Timm, C.D.; Magalhães, K.; Alvarez, F.; Sabaj Pérez, and F. Duponchelle.
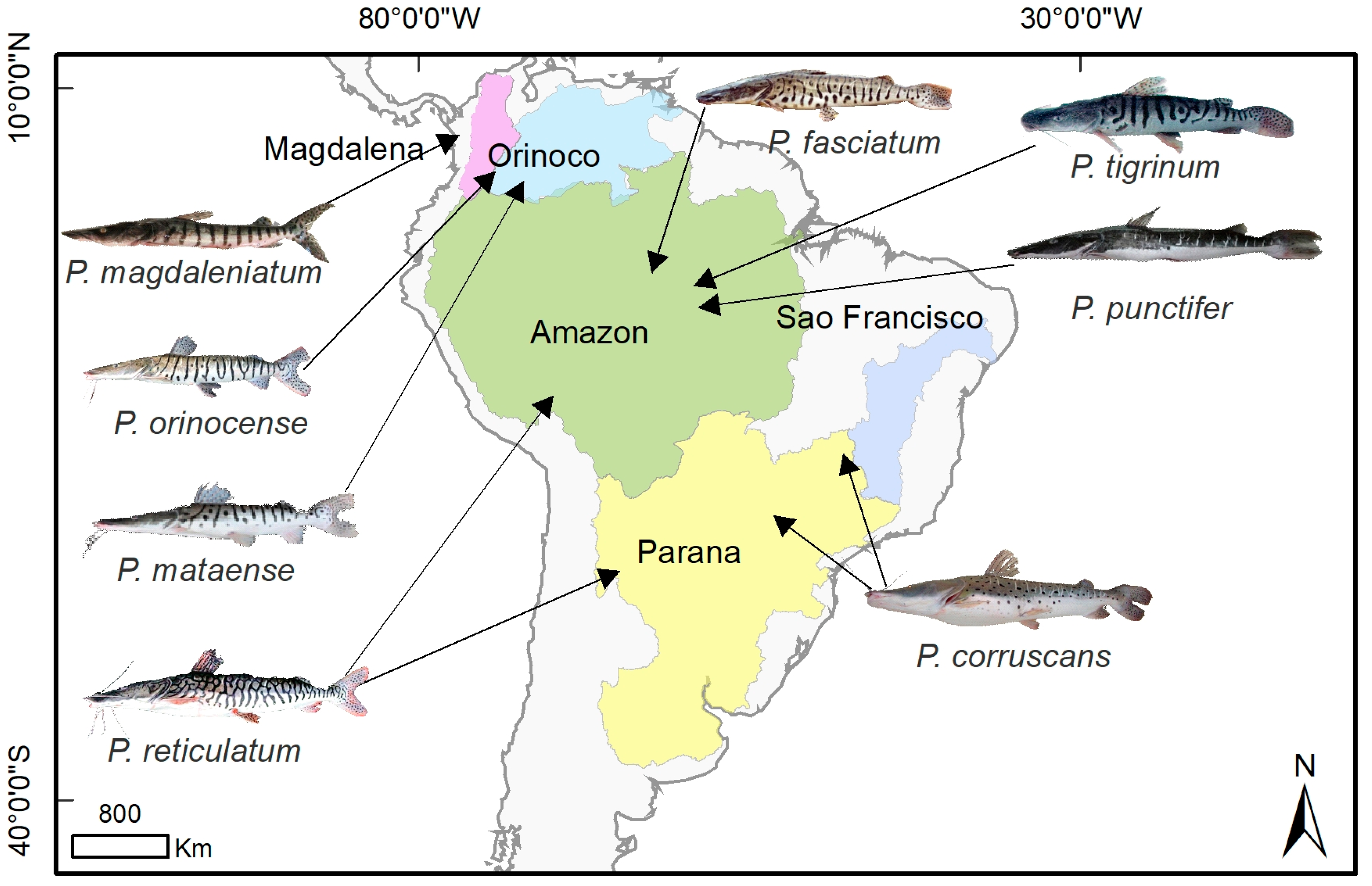
Table 2.
Summary of von Bertalanffy growth parameters and Ø values for Pseudoplatystoma species.
| Species | L∞ (cm) | K | Longevity (Years) | Ø | Ref. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L∞ | L∞m | L∞f | k | km | kf | Longevity | Male | Female | Ø | Øm | Øf | ||
| P. tigrinum | 180 | - | - | 0.29 | - | - | - | - | - | 0.97 | - | - | [56] |
| P. tigrinum | - | 146 | 170 | - | 0.11 | 0.01 | 25–30 | - | - | - | 0.49 | −0.55 | [51] |
| P. tigrinum | - | 119.8 | 131.8 | - | 0.17 | 0.15 | - | - | - | - | 0.63 | 0.59 | [32] |
| P. fasciatum | - | 77 | 103.1 | - | 0.44 | 0.24 | 15 | - | - | - | 0.91 | 0.72 | [32] |
| P. corruscans | 200 | - | - | 0.08 | - | - | 10 | - | - | 0.47 | - | - | [57] |
| P. corruscans | - | 99.2 | 131.8 | 0.51 | 0.34 | - | - | - | - | 1.05 | 0.95 | [58] | |
| P. reticulatum | - | 72.7 | 82.5 | - | 0.44 | 0.33 | - | - | - | - | 0.89 | 0.81 | [59] |
| P. punctifer | - | 99.5 | 97.3 | - | 0.16 | 0.2 | - | - | - | - | 0.54 | 0.63 | [15] |
| P. mataense | - | 108.6 | 119.9 | - | 0.19 | 0.17 | - | 12.37 | 15.7 | - | 0.64 | 0.62 | [60] |
Table 3.
Definitions of key genetic concepts.
| Concepts | Ecological or Genetic Meaning |
|---|---|
| Analysis of Molecular Variance (AMOVA) | An analytical algorithm that partitions genetic variation into within- and between-population components. |
| Bayesian methods | Methods of statistical inference in which Bayes’ theorem is used to update the probability for a hypothesis as more evidence or information becomes available. Bayesian approaches underlie some clustering and assignment tests and phylogenetic inference algorithms in population genetic analyses. |
| Evolutionary Significant Units (ESU) | A population or a group of populations that merits priority for conservation and separate management because of high genetic and ecological distinctiveness from other such units. |
| FST | The coefficient of departure of genotype frequencies from Hardy-Weinberg equilibrium due to differentiation among subpopulations within a larger population. |
| Fu’s Fs | A population genetics statistic helpful in assessing historical population dynamics. |
| Genetic differentiation | The condition of populations having contrasting genotype frequencies resulting from the dynamic interplay among mutation, gene flow, selection, and random genetic drift over space and time. |
| Management Unit (MU) | A population that is demographically independent from others and hence should be managed separately. |
| Maximum parsimony | In the context of phylogenetic inference, a method for evaluating relationships between lineages in which the evolutionary tree with the fewest common ancestors is regarded as the most likely. |
| Microsatellite | A genomic sequence with repeats of 1–6 nucleotide motifs, e.g., GT, CAC, or GACA. There is variation in the repeat number at a given locus among members of a chromosome pair in an individual, among individuals within a population, and often among populations. Such genetic markers are highly useful for population genetic studies. |
| Mitochondrial DNA COI region | Mitochondrial cytochrome oxidase subunit 3 (COI) gene. This genetic marker is often used for DNA barcoding to distinguish species. |
| Neighbor-likelihood joining | An agglomerative clustering method for the construction of phylogenetic trees. |
| Panmictic population (panmixia) | A collection of fish with the absence of genetic structure (the condition of being well mixed, as in a population where all individuals could interbreed). |
| Phylogenetic tree | A representation of the inferred evolutionary relationships among individuals, populations, or species that shows the branching from a known or hypothesized ancestor to its derived lineages, which outwardly resembles a tree and hence is referred to as this. |
| RST | A metric of genetic differentiation that incorporates the differences in the size of microsatellite alleles |
| Tajima’s D | The difference between two measures of genetic diversity: the mean number of pairwise differences and the number of segregating sites, each scaled so that they are expected to be the same in a neutrally evolving population of constant size. |
Table 4.
Summary of information available and knowledge gaps for eight species of genus Pseudoplatystoma.
Table 4.
Summary of information available and knowledge gaps for eight species of genus Pseudoplatystoma.
| Species | Genetics | Fisheries Status | Growth | Reproduction | Movement Ecology |
|---|---|---|---|---|---|
| P. fasciatum | Microsatellite DNA showed high genetic diversity with 200 km of genetic isolation by distance | No data | Validation, growth curves | Total spawning in the rising and high-water seasons 31, 52, 43 | Adults migrate, juveniles are nonmigratory 31~164 km, bidirectional movement during the dry season and the rising water period 39 |
| P. tigrinum | No data | No data; exploited very close to maximum sustainable 54 | Validation, growth curves | Total spawning in the rising water season 44, 52 | Adults migrate, juveniles are nonmigratory 31 |
| P. punctifer | No data | No data | Validation, growth curves | No data | No data |
| P. reticulatum | No data | No data | No validation data, growth curves | Total spawning in the rising water season 46 | Believed to be migratory |
| P. mataense | No data | No data | Validation, growth curves | No data | Believed to be migratory |
| P. orinocense | No data | No data | No data | No data | Believed to be migratory |
| P. magdaleniatum | No data | No data | No data | Spawning in the rising water season 45 | Believed to be migratory |
| P. corruscans | Mitochondrial COI region, microsatellite DNA analysis to delineate 2 ESUs | No data; strongly overfished 71 | No validation data, growth curves | Total spawning in the rising water season 32, 46 | Partial migration of females; homing migration 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pereira, L.A.; Castello, L.; Orth, D.J.; Duponchelle, F.; Hallerman, E.M. A Synthesis of the Ecology and Conservation of Pseudoplatystoma Catfishes in the Neotropics. Fishes 2023, 8, 306. https://doi.org/10.3390/fishes8060306
AMA Style
Pereira LA, Castello L, Orth DJ, Duponchelle F, Hallerman EM. A Synthesis of the Ecology and Conservation of Pseudoplatystoma Catfishes in the Neotropics. Fishes. 2023; 8(6):306. https://doi.org/10.3390/fishes8060306
Chicago/Turabian StylePereira, Luciana A., Leandro Castello, Donald J. Orth, Fabrice Duponchelle, and Eric M. Hallerman. 2023. "A Synthesis of the Ecology and Conservation of Pseudoplatystoma Catfishes in the Neotropics" Fishes 8, no. 6: 306. https://doi.org/10.3390/fishes8060306