
A Cartilage Matrix Protein Regulates Collagen Synthesis in Mantle of Magallana gigas (Crassostrea gigas) under Ocean Acidification


Abstract
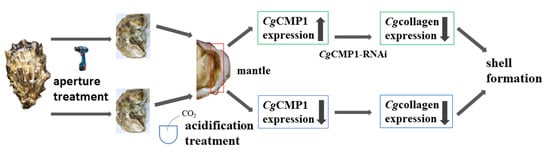
1. Introduction
2. Materials and Methods
2.1. Oyster Treatment, and Sample Collection
2.1.1. Obtaining and Maintenance of Oysters
2.1.2. Aperture and Acidification Treatment
2.1.3. Measurement of Length and Thickness of New-Formation Shells
2.1.4. Sample Collection
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Gene Cloning and Sequence Analysis
2.4. qRT-PCR Analysis of mRNA Expression
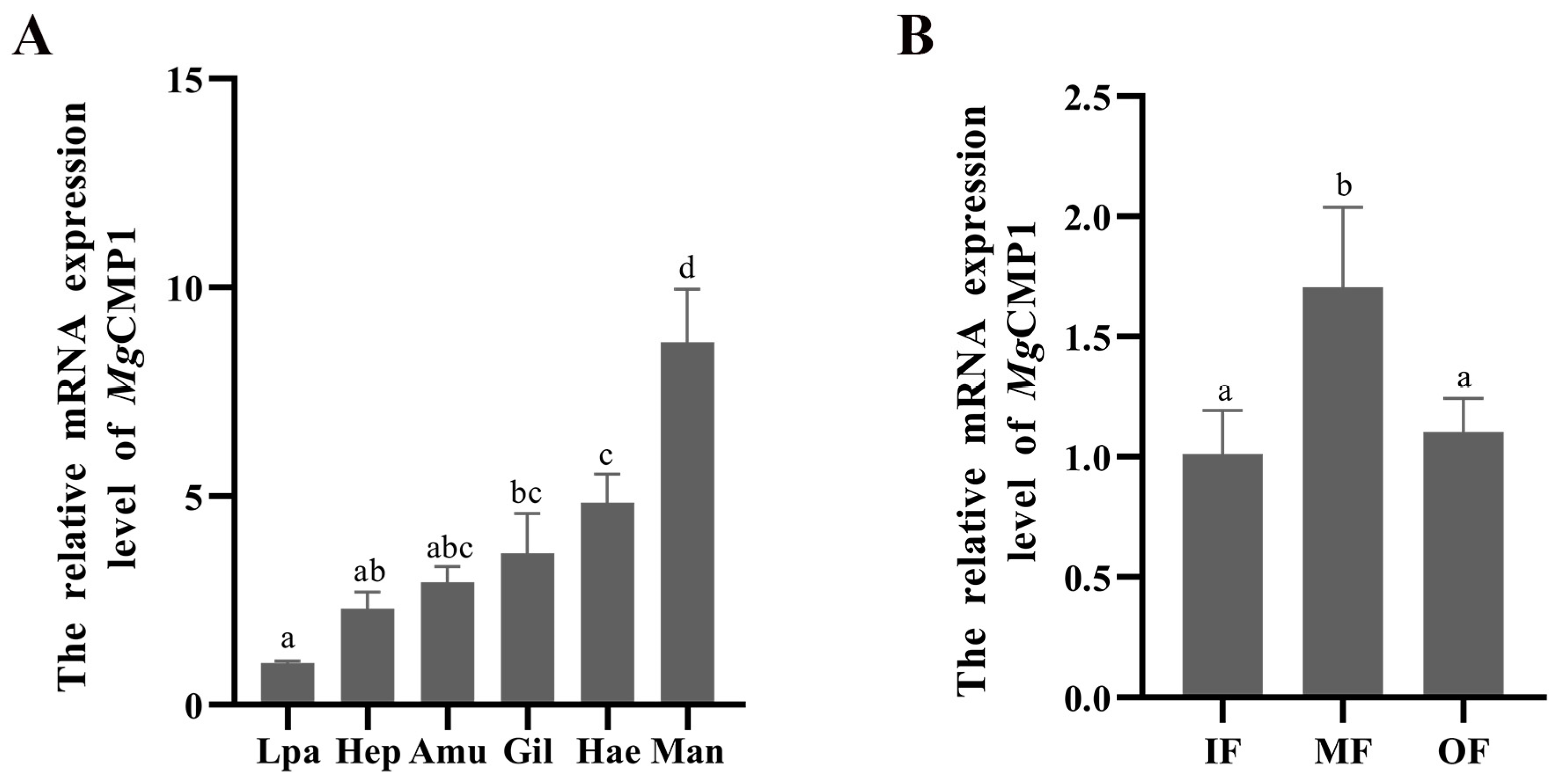
2.5. Recombinant Expression and Purification of MgCMP1
2.6. The Molecular Interaction between rMgCMP1 and rMgBMP7
2.7. In Vivo RNA Interference (RNAi) Treatment of Oysters
2.8. In Situ Hybridization
2.8.1. RNA Probe Preparation
2.8.2. ISH Techniques
2.9. Statistical Analysis
3. Results
3.1. The Sequence Characteristics and Phylogenetic Relationship of MgCMP1
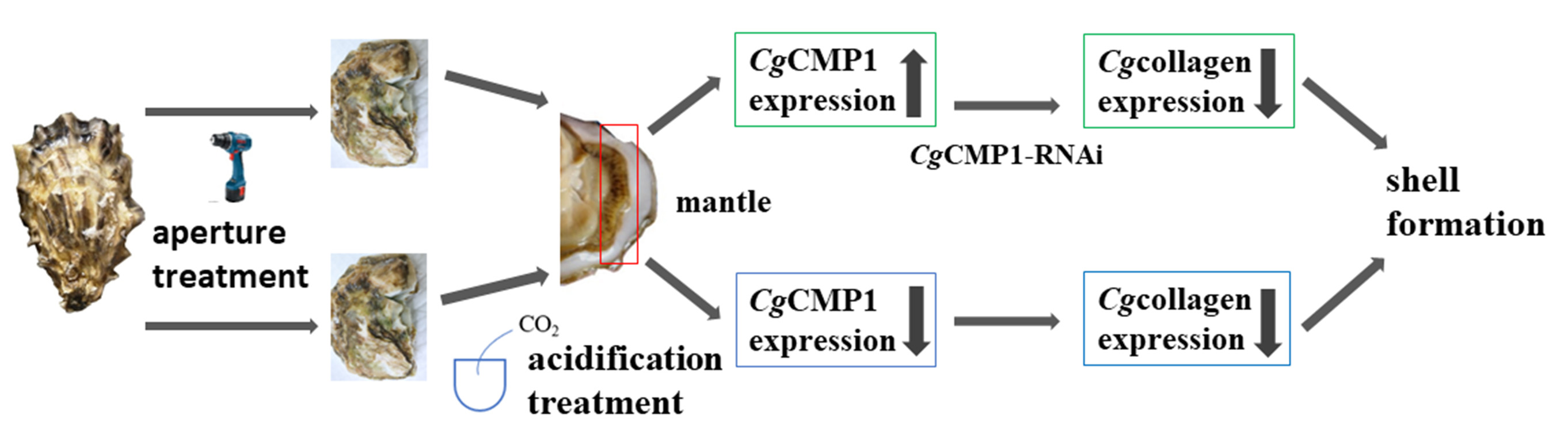
3.2. The Distribution of MgCMP1 mRNA in Different Tissues
3.3. Recombinant Protein of rMgCMP1 and Its Interaction with rMgBMP7 In Vitro
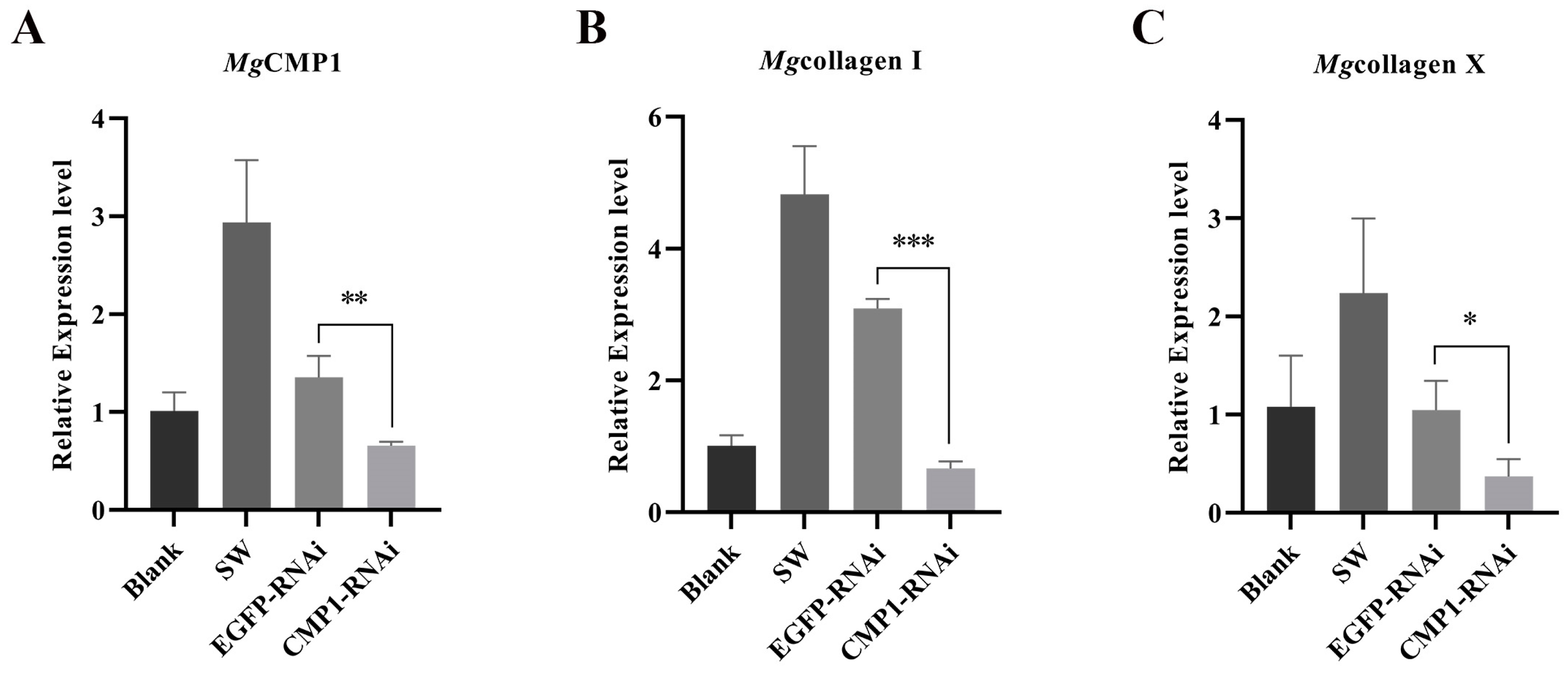
3.4. The mRNA Transcripts of Mgcollagen I and Mgcollagen X in MgCMP1-RNAi Oysters
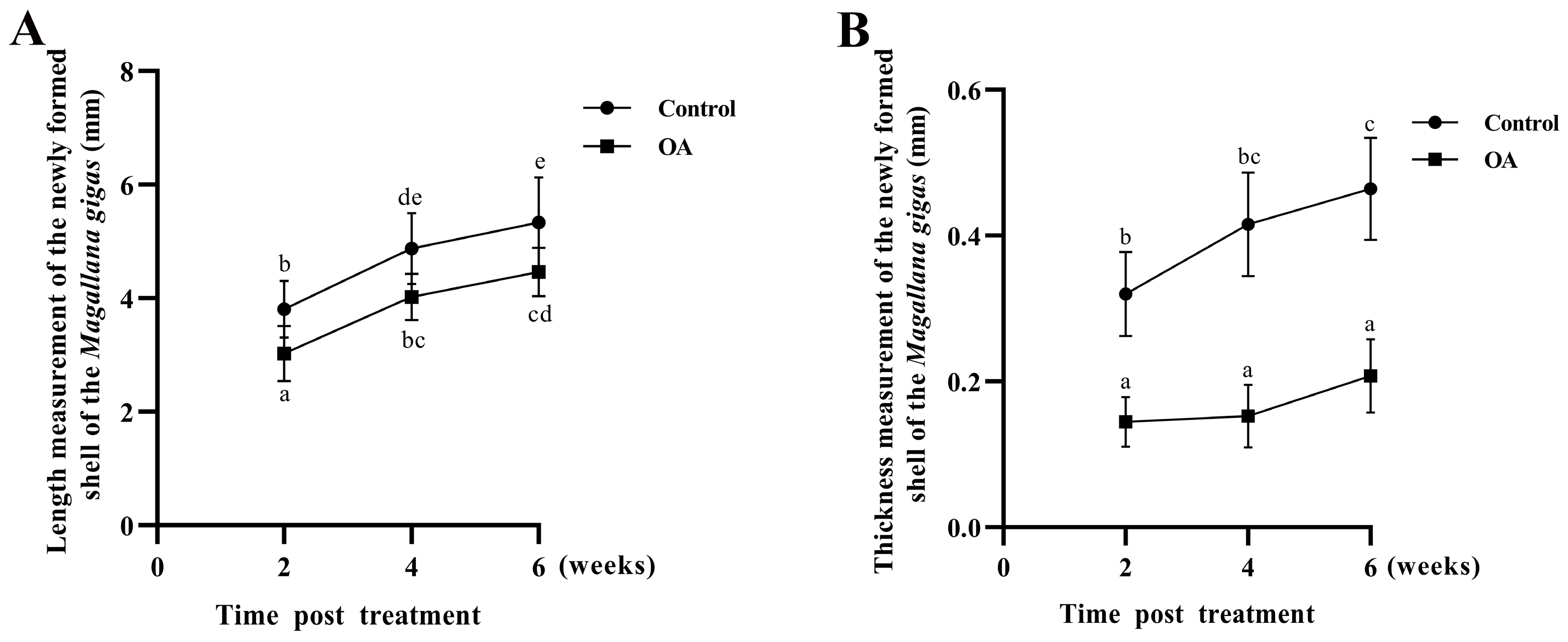
3.5. Morphologic Characteristics of New-Formed Shell in Punctured Oysters under Acidification Treatment
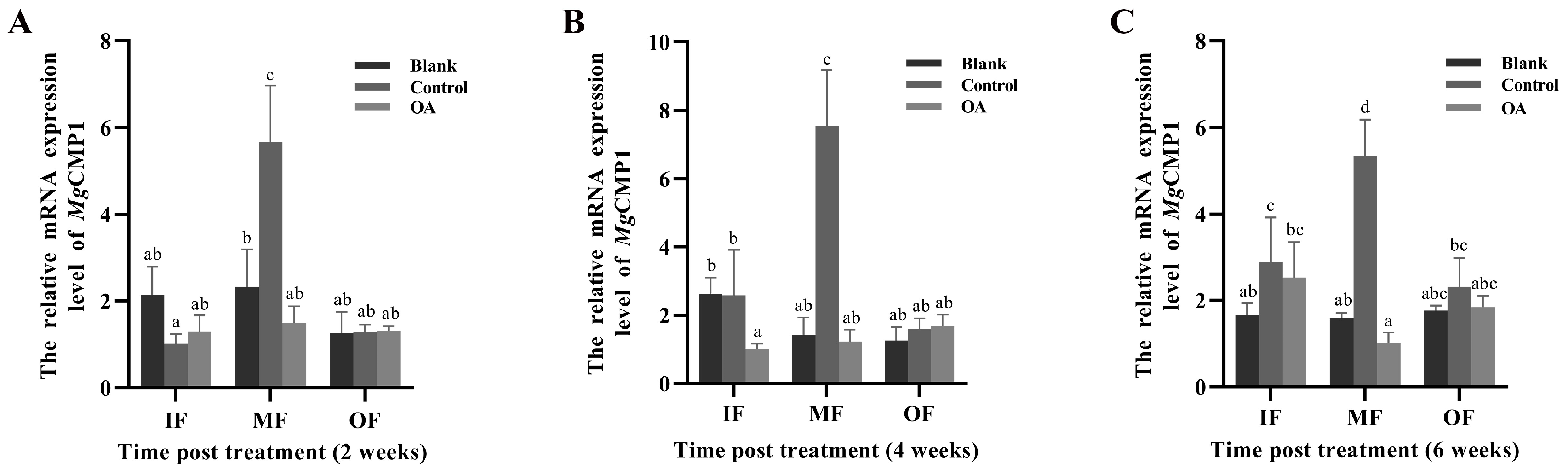
3.6. The mRNA Expression Levels of MgCMP1 in IF, MF and OF of Mantle of Punctured Oysters under Acidification Treatment
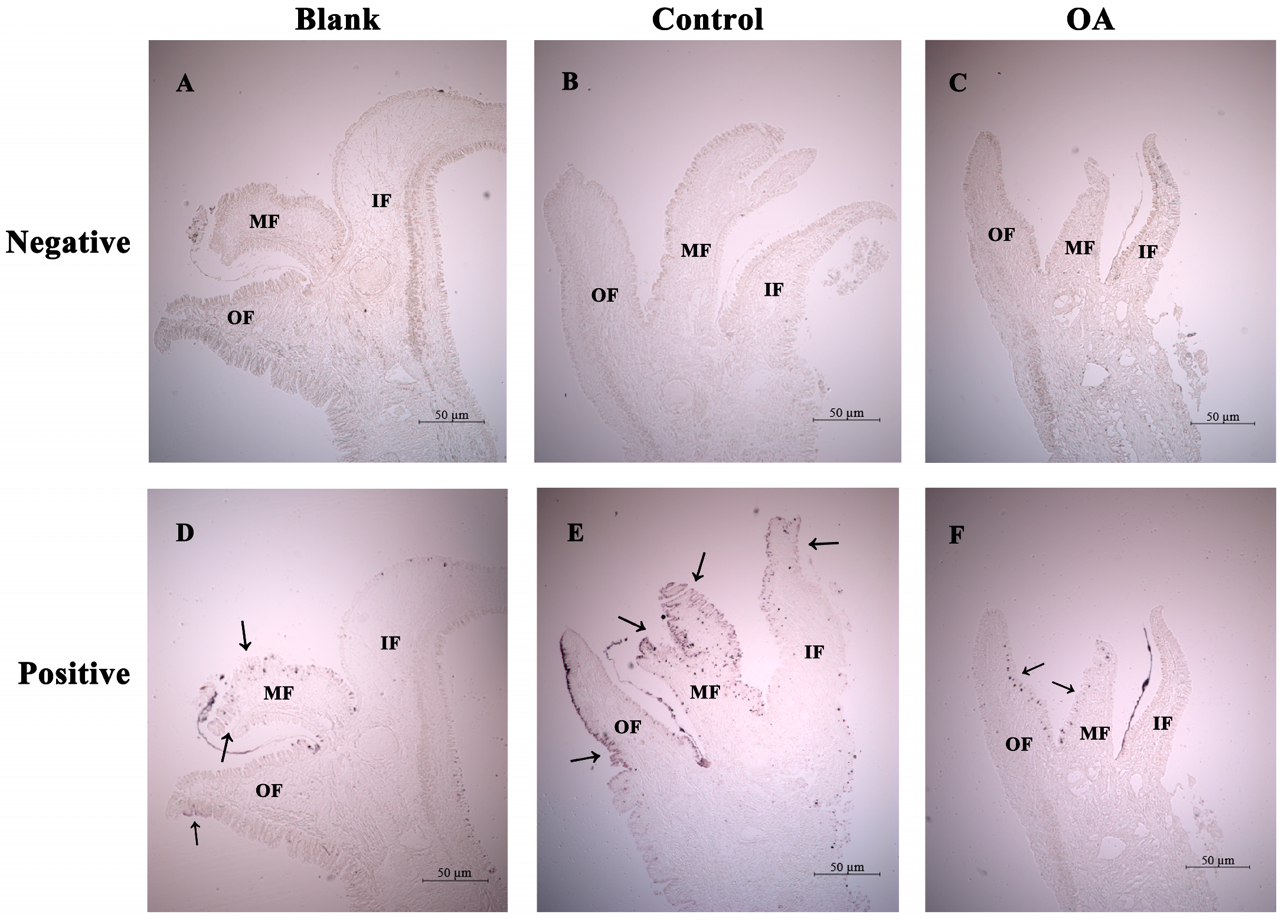
3.7. The Distribution of MgCMP1 mRNA Transcripts in IF, MF and OF in Mantle of Punctured Oysters under Acidification Treatment
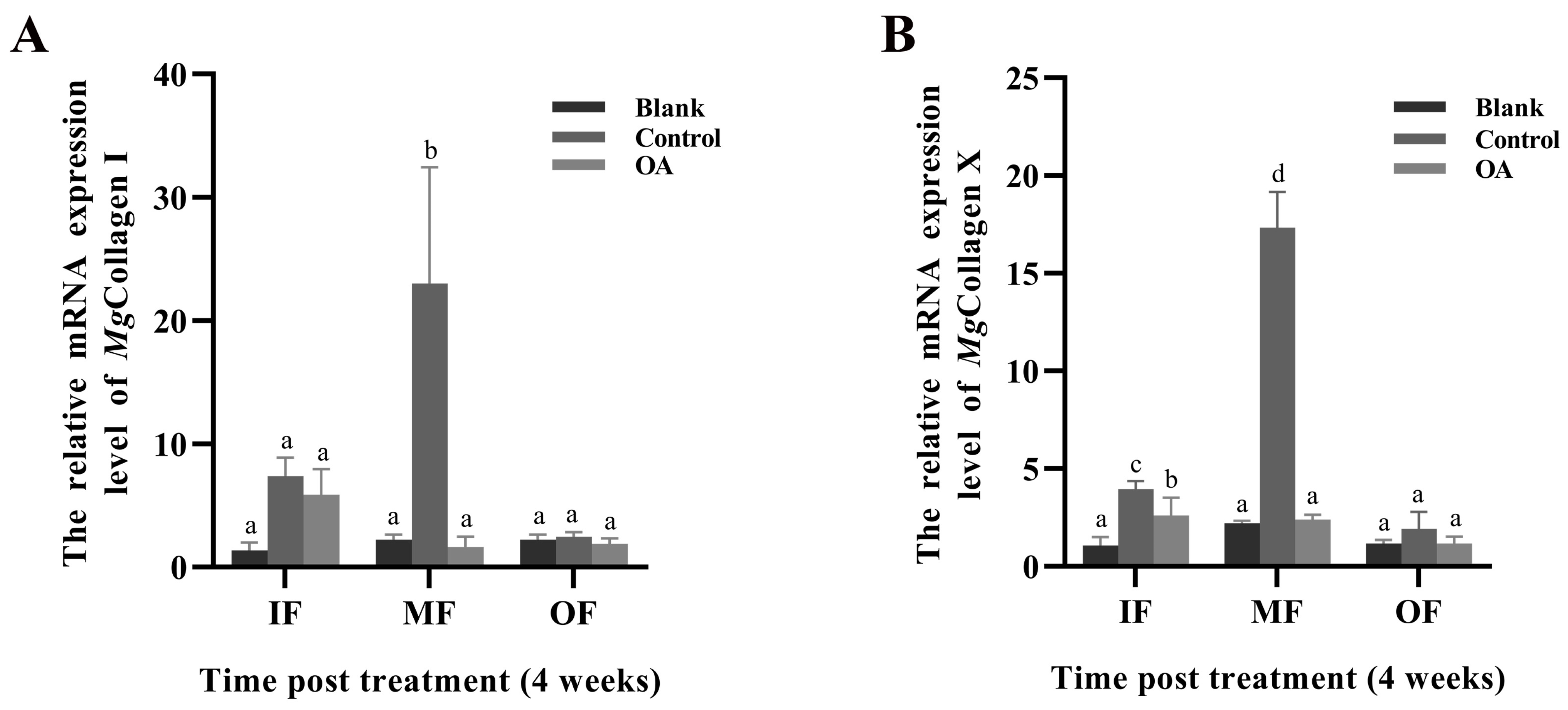
3.8. The mRNA Expression Levels of Mgcollagen I and Mgcollagen X in IF, MF and OF in Mantle of Punctured Oysters under Acidification Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michaelidis, B.; Ouzounis, C.; Paleras, A.; Pörtner, H.-O. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine mussels (Mytilus galloprovincialis). Mar. Ecol. Prog. Ser. 2005, 293, 109–118. [Google Scholar] [CrossRef]
- Berge, J.A.; Bjerkeng, B.; Pettersen, O.; Schaanning, M.T.; Øxnevad, S. Effects of increased sea water concentrations of CO2 on growth of the bivalve Mytilus edulis L. Chemosphere 2006, 62, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, K.; Wickett, M.E. Oceanography: Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed]
- Fitzer, S.C.; Phoenix, V.R.; Cusack, M.; Kamenos, N.A. Ocean acidification impacts mussel control on biomineralisation. Sci. Rep. 2014, 4, 6218. [Google Scholar] [CrossRef]
- Freer, A.; Bridgett, S.; Jiang, J.; Cusack, M. Biomineral Proteins from Mytilus edulis Mantle Tissue Transcriptome. Mar. Biotechnol. 2014, 16, 34–45. [Google Scholar] [CrossRef]
- Marin, F.; Luquet, G. Molluscan shell proteins. Comptes Rendus Palevol 2004, 3, 469–492. [Google Scholar] [CrossRef]
- Marin, F.; Luquet, G.; Marie, B.; Medakovic, D. Molluscan Shell Proteins: Primary Structure, Origin, and Evolution. Curr. Top. Dev. Biol. 2007, 80, 209–276. [Google Scholar] [CrossRef]
- Comeau, S.; Edmunds, P.; Spindel, N.; Carpenter, R. The responses of eight coral reef calcifiers to increasing partial pressure of CO2 do not exhibit a tipping point. Limnol. Oceanogr. 2013, 58, 388–398. [Google Scholar] [CrossRef]
- Gazeau, F.P.H.; Quiblier, C.M.L.; Jansen, J.M.; Gattuso, J.P.; Middelburg, J.J.; Heip, C.H.R. Impact of elevated CO2 on shellfish calcification. J. Geophys. Res. 2007, 34, L07603. [Google Scholar] [CrossRef]
- Orr, J.C. Recent and Future Changes in Ocean Carbonate Chemistry. In Ocean Acidification; Gattuso, J.-P., Hansson, L., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 41–66. [Google Scholar] [CrossRef]
- Huang, J.; Li, S.; Liu, Y.; Liu, C.; Xie, L.; Zhang, R. Hemocytes in the extrapallial space of Pinctada fucata are involved in immunity and biomineralization. Sci. Rep. 2018, 8, 4657. [Google Scholar] [CrossRef]
- Bubel, A. An electron-microscope study of periostracum formation in some marine bivalves. II. The cells lining the periostracal groove. Mar. Biol. 1973, 20, 222–234. [Google Scholar] [CrossRef]
- Nakayama, S.; Suzuki, M.; Endo, H.; Iimura, K.; Kinoshita, S.; Watabe, S.; Kogure, T.; Nagasawa, H. Identification and characterization of a matrix protein (PPP-10) in the periostracum of the pearl oyster, Pinctada fucata. FEBS Open Bio 2013, 3, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Feng, Q.; Chi, Y.; Xie, L.; Zhang, R. Investigation of cell proliferation and differentiation in the mantle of Pinctada fucata (Bivalve, Mollusca). Mar. Biol. 2008, 153, 745–754. [Google Scholar] [CrossRef]
- Mao, J.X.; Zhang, W.J.; Wang, X.B.; Song, J.; Yin, D.H.; Tian, Y.; Hao, Z.L.; Han, B.; Chang, Y.Q. Histological and Expression Differences Among Different Mantle Regions of the Yesso Scallop (Patinopecten yessoensis) Provide Insights into the Molecular Mechanisms of Biomineralization and Pigmentation. Mar. Biotechnol. 2019, 21, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Kádár, E. Haemocyte response associated with induction of shell regeneration in the deep-sea vent mussel Bathymodiolus azoricus (Bivalvia: Mytilidae). J. Exp. Mar. Biol. Ecol. 2008, 362, 71–78. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Zong, Y.; Zheng, Y.; Li, Y.; Han, Z.; Wang, L.; Song, L. The Increased Expression of an Engrailed to Sustain Shell Formation in Response to Ocean Acidification. Front. Physiol. 2020, 11, 530435. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, G.; Fan, S.; Liu, B.; Zhang, B.; Su, J.; Yu, D. Molecular cloning and expression profiles of matrilin-1 inpearl oyster (Pinctada fucata). S. China Fish. Sci. 2017, 13, 76–84. Available online: https://www.cabdirect.org/cabdirect/welcome/?target=%2fcabdirect%2fabstract%2f20173126376 (accessed on 1 May 2023).
- Mörgelin, M.; Heinegård, D.; Engel, J.; Paulsson, M. Electron microscopy of native cartilage oligomeric matrix protein purified from the Swarm rat chondrosarcoma reveals a five-armed structure. J. Biol. Chem. 1992, 267, 6137–6141. [Google Scholar] [CrossRef]
- Deák, F.; Wagener, R.; Kiss, I.; Paulsson, M. The matrilins: A novel family of oligomeric extracellular matrix proteins. Matrix Biol. 1999, 18, 55–64. [Google Scholar] [CrossRef]
- Klatt, A.R.; Becker, A.-K.A.; Neacsu, C.D.; Paulsson, M.; Wagener, R. The matrilins: Modulators of extracellular matrix assembly. Int. J. Biochem. Cell Biol. 2011, 43, 320–330. [Google Scholar] [CrossRef]
- Becker, A.-K.A.; Mikolajek, H.; Werner, J.M.; Paulsson, M.; Wagener, R. Characterization of recombinantly expressed matrilin VWA domains. Protein Expr. Purif. 2015, 107, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, X.; Wang, T.; Zhu, Q.; Miao, G.; Chen, Y.; Fang, X.; Que, H.; Li, L.; Zhang, G. Evolution and functional analysis of the Pif97 gene of the Pacific oyster Crassostrea gigas. Curr. Zool. 2013, 59, 109–115. [Google Scholar] [CrossRef]
- Posey, K.L.; Coustry, F.; Hecht, J.T. Cartilage oligomeric matrix protein: COMP opathies and beyond. Matrix Biol. 2018, 71–72, 161–173. [Google Scholar] [CrossRef]
- Ishida, K.; Acharya, C.; Christiansen, B.A.; Yik, J.H.N.; DiCesare, P.E.; Haudenschild, D.R. Cartilage oligomeric matrix protein enhances osteogenesis by directly binding and activating bone morphogenetic protein-2. Bone 2013, 55, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.; Karsai, A.; Fong, M.C.; Cai, W.; Yik, J.H.N.; Klineberg, E.; Haudenschild, D.R.; Liu, G.Y. Label-Free and Direct Visualization of Multivalent Binding of Bone Morphogenetic Protein-2 with Cartilage Oligomeric Matrix Protein. J. Phys. Chem. B 2019, 123, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Nicolae, C.; Ko, Y.P.; Miosge, N.; Niehoff, A.; Studer, D.; Enggist, L.; Hunziker, E.B.; Paulsson, M.; Wagener, R.; Aszodi, A. Abnormal Collagen Fibrils in Cartilage of Matrilin-1/Matrilin-3-deficient Mice*. J. Biol. Chem. 2007, 282, 22163–22175. [Google Scholar] [CrossRef] [PubMed]
- Zaucke, F.; Grässel, S. Genetic mouse models for the functional analysis of the perifibrillar components collagen IX, COMP and matrilin-3: Implications for growth cartilage differentiation and endochondral ossification. Histol. Histopathol. 2009, 24, 1067–1079. [Google Scholar] [CrossRef]
- Jayasuriya, C.T.; Goldring, M.B.; Terek, R.; Chen, Q. Matrilin-3 induction of IL-1 receptor antagonist is required for up-regulating collagen II and aggrecan and down-regulating ADAMTS-5 gene expression. Arthritis Res. Ther. 2012, 14, R197. [Google Scholar] [CrossRef]
- Muttigi, M.S.; Kim, B.J.; Choi, B.; Yoshie, A.; Kumar, H.; Han, I.; Park, H.; Lee, S.H. Matrilin-3 codelivery with adipose-derived mesenchymal stem cells promotes articular cartilage regeneration in a rat osteochondral defect model. J. Tissue Eng. Regen. Med. 2018, 12, 667–675. [Google Scholar] [CrossRef]
- Blumbach, K.; Niehoff, A.; Paulsson, M.; Zaucke, F. Ablation of collagen IX and COMP disrupts epiphyseal cartilage architecture. Matrix Biol. 2008, 27, 306–318. [Google Scholar] [CrossRef]
- Neacsu, C.D.; Ko, Y.P.; Tagariello, A.; Karlsen, R.K.; Neiss, W.F.; Paulsson, M.; Wagener, R. Matrilin-1 is essential for zebrafish development by facilitating collagen II secretion. J. Biol. Chem. 2014, 289, 1505–1518. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, Z.; Wang, L.; Song, L. Recent Advances of Shell Matrix Proteins and Cellular Orchestration in Marine Molluscan Shell Biomineralization. Front. Mar. Sci. 2019, 6, 41. [Google Scholar] [CrossRef]
- Bouchut, A.; Roger, E.; Coustau, C.; Gourbal, B.; Mitta, G. Compatibility in the Biomphalaria glabrata/Echinostoma caproni model: Potential involvement of adhesion genes. Int. J. Parasitol. 2006, 36, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.F.; Fang, X.D.; Guo, X.M.; Li, L.; Luo, R.B.; Xu, F.; Yang, P.C.; Zhang, L.L.; Wang, X.T.; Qi, H.G.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Iwashima, A.; Tsutsui, N.; Ohira, T.; Kogure, T.; Nagasawa, H. Identification and characterisation of a calcium carbonate-binding protein, blue mussel shell protein (BMSP), from the nacreous layer. Chembiochem 2011, 12, 2478–2487. [Google Scholar] [CrossRef]
- Kocot, K.M.; Cannon, J.T.; Todt, C.; Citarella, M.R.; Kohn, A.B.; Meyer, A.; Santos, S.R.; Schander, C.; Moroz, L.L.; Lieb, B.; et al. Phylogenomics reveals deep molluscan relationships. Nature 2011, 477, 452–456. [Google Scholar] [CrossRef]
- Wang, X.D.; Wang, M.Q.; Jia, Z.H.; Song, X.R.; Wang, L.L.; Song, L.S. A shell-formation related carbonic anhydrase in Crassostrea gigas modulates intracellular calcium against CO2 exposure: Implication for impacts of ocean acidification on mollusk calcification. Aquat. Toxicol. 2017, 189, 216–228. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Song, X.; Huang, S.; Wang, L.; Song, L. The Inhibition of Ocean Acidification on the Formation of Oyster Calcified Shell by Regulating the Expression of Cgchs1 and Cgchit4. Front. Physiol. 2019, 10, 1034. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, T.; Wang, L.; Xu, J.; Li, M.; Zhang, A.; Qiu, L.; Song, L. A new non-phagocytic TLR6 with broad recognition ligands from Pacific oyster Crassostrea gigas. Dev. Comp. Immunol. 2016, 65, 182–190. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, P.; Wang, W.; Dong, M.; Wang, M.; Gong, C.; Jia, Z.; Liu, Z.; Zhang, A.; Wang, L.; et al. A DM9-containing protein from oyster Crassostrea gigas (CgDM9CP-2) serves as a multipotent pattern recognition receptor. Dev. Comp. Immunol. 2018, 84, 315–326. [Google Scholar] [CrossRef]
- Jia, Z.; Tao, Z.; Jiang, S.; Wang, M.; Cheng, Q.; Sun, M.; Wang, L.; Song, L. An integrin from oyster Crassostrea gigas mediates the phagocytosis toward Vibrio splendidus through LPS binding activity. Dev. Comp. Immunol. 2015, 53, 253–264. [Google Scholar] [CrossRef]
- Li, Q.; Wang, W.L.; Wu, S.S.; Li, J.L.; Dong, M.R.; Wang, L.L.; Song, L.S. CgBlimp-1 inhibits granulocytes proliferation and interleukin production in the immune response of oyster Crassostrea gigas. Dev. Comp. Immunol. 2023, 142, 104652. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.L.; Lv, X.J.; Liu, Z.Q.; Song, X.R.; Yi, Q.L.; Wang, L.L.; Song, L.S. The sensing pattern and antitoxic response of Crassostrea gigas against extracellular products of Vibrio splendidus. Dev. Comp. Immunol. 2020, 102, 103467. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.Q.; Wang, W.L.; Fan, S.Q.; Li, J.L.; Li, Q.; Wu, S.S.; Wang, L.L.; Song, L.S. The receptor CgIL-17R1 expressed in granulocytes mediates the CgIL-17 induced haemocytes proliferation in Crassostrea gigas. Dev. Comp. Immunol. 2022, 131, 104376. [Google Scholar] [CrossRef]
- Shah, N.B.; Duncan, T.M. Bio-layer interferometry for measuring kinetics of protein-protein interactions and allosteric ligand effects. J. Vis. Exp. 2014, 84, e51383. [Google Scholar] [CrossRef]
- Xin, X.; Liu, C.; Liu, Z.; Zhang, Y.; Gao, Y.; Zhu, T.; Wang, L.; Song, L. Calmodulin regulates the calcium homeostasis in mantle of Crassostrea gigas under ocean acidification. Front. Mar. Sci. 2022, 9, 1050022. [Google Scholar] [CrossRef]
- Johnson, K.L.; Zhen, D.K.; Bianchi, D.W. The use of fluorescence in situ hybridization (FISH) on paraffin-embedded tissue sections for the study of microchimerism. Biotechniques 2000, 29, 1220–1224. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Huang, W.; Yao, B.; Sun, L.; Pu, R.; Wang, L.; Zhang, R. Immunohistochemical and in situ hybridization studies of gonadotropin releasing hormone (GnRH) and its receptor in rat digestive tract. Life Sci. 2001, 68, 1727–1734. [Google Scholar] [CrossRef]
- Hendriks, I.; Duarte, C.; Olsen, Y.; Steckbauer, A.; Ramajo, L.; Moore, T.; Trotter, J.; McCulloch, M. Biological mechanisms supporting adaptation to ocean acidification in coastal ecosystems. Estuar. Coast. Shelf Sci. 2015, 152, A1–A8. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Salisbury, J.E. Ocean acidification in the coastal zone from an organism′s perspective: Multiple system parameters, frequency domains, and habitats. Ann. Rev. Mar. Sci. 2014, 6, 221–247. [Google Scholar] [CrossRef] [PubMed]
- Mount, A.S.; Wheeler, A.P.; Paradkar, R.P.; Snider, D. Hemocyte-mediated shell mineralization in the eastern oyster. Science 2004, 304, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liang, J.; Sun, J.; Gao, J.; Zhang, S.r.; Liu, Y.j.; Xie, L.p.; Zhang, R.q. Influence of the Extrapallial Fluid of Pinctada fucata on the Crystallization of Calcium Carbonate and Shell Biomineralization. Cryst. Growth Des. 2016, 16, 672–680. [Google Scholar] [CrossRef]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk shell formation: A source of new concepts for understanding biomineralization processes. Chemistry 2006, 12, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, T.; Schwarzinger, C.; Miksik, I.; Smrz, M.; Beran, A. Molluscan shell evolution with review of shell calcification hypothesis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 351–371. [Google Scholar] [CrossRef]
- Watabe, N. Studies on shell formation: XI. Crystal—Matrix relationships in the inner layers of mollusk shells. J. Ultrastruct. Res. 1965, 12, 351–370. [Google Scholar] [CrossRef]
- Xu, W.; Faisal, M. Matrilin-like molecules produced by circulating hemocytes of the zebra mussel (Dreissena polymorpha) upon stimulation. Dev. Comp. Immunol. 2007, 31, 1205–1210. [Google Scholar] [CrossRef]
- Sun, Q.; Jiang, Y.; Fan, M.; Zhang, X.; Xu, H.; Liao, Z. Characterization of a novel shell matrix protein with vWA domain from Mytilus coruscus. Biosci. Biotechnol. Biochem. 2020, 84, 1629–1644. [Google Scholar] [CrossRef]
- Bjärnmark, N.A.; Yarra, T.; Churcher, A.M.; Felix, R.C.; Clark, M.S.; Power, D.M. Transcriptomics provides insight into Mytilus galloprovincialis (Mollusca: Bivalvia) mantle function and its role in biomineralisation. Mar. Genom. 2016, 27, 37–45. [Google Scholar] [CrossRef]
- Yarra, T.; Gharbi, K.; Blaxter, M.; Peck, L.S.; Clark, M.S. Characterization of the mantle transcriptome in bivalves: Pecten maximus, Mytilus edulis and Crassostrea gigas. Mar. Genom. 2016, 27, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Gardner, L.D.; Mills, D.; Wiegand, A.; Leavesley, D.; Elizur, A. Spatial analysis of biomineralization associated gene expression from the mantle organ of the pearl oyster Pinctada maxima. BMC Genom. 2011, 12, 455. [Google Scholar] [CrossRef]
- Fan, S.; Zhou, D.; Xu, Y.; Yu, D. Cloning and functional analysis of BMP3 in the pearl oyster (Pinctada fucata). J. Appl. Anim. Res. 2019, 47, 250–261. [Google Scholar] [CrossRef]
- Rosenberg, K.; Olsson, H.; Mörgelin, M.; Heinegård, D. Cartilage Oligomeric Matrix Protein Shows High Affinity Zinc-dependent Interaction with Triple Helical Collagen*. J. Biol. Chem. 1998, 273, 20397–20403. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Trehan, S.K.; Guan, Y.j.; Sun, C.q.; Moore, D.C.; Jayasuriya, C.T.; Chen, Q. Matrilin-3 Inhibits Chondrocyte Hypertrophy as a Bone Morphogenetic Protein-2 Antagonist*. J. Biol. Chem. 2014, 289, 34768–34779. [Google Scholar] [CrossRef]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Ann. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef]
- Emma, T.-S.; Coffey, W.D.; Hua, W.; Nunn, B.L.; Dickinson, G.H.; Roberts, S.B. Shotgun proteomics reveals physiological response to ocean acidification in Crassostrea gigas. BMC Genom. 2014, 15, 951. [Google Scholar] [CrossRef]
- Hofmann, G.E.; Barry, J.P.; Edmunds, P.J.; Gates, R.D.; Hutchins, D.A.; Klinger, T.; Sewell, M.A. The Effect of Ocean Acidification on Calcifying Organisms in Marine Ecosystems: An Organism to Ecosystem Perspective. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 127–147. [Google Scholar] [CrossRef]
- Liu, W.; Yu, Z.; Huang, X.; Shi, Y.; Lin, J.; Zhang, H.; Yi, X.; He, M. Effect of ocean acidification on growth, calcification, and gene expression in the pearl oyster, Pinctada fucata. Mar. Environ. Res. 2017, 130, 174–180. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) |
|---|---|
| Clone primers | |
| MgCMP1-KL-F | TGATGAACGACCCGCTTA |
| MgCMP1-KL-R | AAAATGTAGGCACGGCTGT |
| MgCMP1-M-F | CCGGAATTCATGCTGACTTTCTTAGTTTTGTG |
| MgCMP1-M-R | CCGCTCGAGATCAATATCAAAACACAGCTCGT |
| RT-PCR primers | |
| MgCMP1-RT-F | ATCGTGAGTGCGTTCGACAT |
| MgCMP1-RT-R | CACGTGACAGTCCATCCGTT |
| MgEF-RT-F | AGTCACCAAGGCTGCACAGAAAG |
| MgEF-RT-R | TCCGACGTATTTCTTTGCGATGT |
| Mgcollagen X-RT-Fi | CGACACCGTGGTGACCAATA |
| Mgcollagen X-RT-Ri | GCATTGCGCACTAACCTCAC |
| Mgcollagen I-RT-Fi | ACCTCCAGGACCTTCGTTTG |
| Mgcollagen I-RT-Ri | TATCCTTGCCGCTGGTGAC |
| MgCMP1-RT-Fi | CCCGGTCGATCTTGTGTTCA |
| MgCMP1-RT-Ri | ACTCCAATCGTGGCTCATCG |
| WEISH primer | |
| MgCMP1-WEISH-F | CTGGGTATCTTTGTGCTTGC |
| MgCMP1-WEISH-R | AAATCCAGTGTCGGTGCC |
| RNAi primer | |
| MgCMP1-RNAi-F1 | GATCACTAATACGACTCACTATAGGGGCTCGC TCTAATGTTGCC |
| MgCMP1-RNAi-R1 | GATCACTAATACGACTCACTATAGGGGGTTGA AAGGGAAAGTCGC |
| EGFP-Fi | GGATCCTAATACGACTCACTATAGGGATCCGAC GTAAACGGCCACAAGT |
| EGFP-Ri | GGATCCTAATACGACTCACTATAGGGATCCTTG TACAGCTCGTCCATGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, T.; Liu, C.; Liu, Z.; Gao, Y.; Xin, X.; Wang, L.; Song, L. A Cartilage Matrix Protein Regulates Collagen Synthesis in Mantle of Magallana gigas (Crassostrea gigas) under Ocean Acidification. Fishes 2023, 8, 290. https://doi.org/10.3390/fishes8060290
Zhu T, Liu C, Liu Z, Gao Y, Xin X, Wang L, Song L. A Cartilage Matrix Protein Regulates Collagen Synthesis in Mantle of Magallana gigas (Crassostrea gigas) under Ocean Acidification. Fishes. 2023; 8(6):290. https://doi.org/10.3390/fishes8060290
Chicago/Turabian StyleZhu, Ting, Chang Liu, Zhaoqun Liu, Yuqian Gao, Xiaoyu Xin, Lingling Wang, and Linsheng Song. 2023. "A Cartilage Matrix Protein Regulates Collagen Synthesis in Mantle of Magallana gigas (Crassostrea gigas) under Ocean Acidification" Fishes 8, no. 6: 290. https://doi.org/10.3390/fishes8060290
APA StyleZhu, T., Liu, C., Liu, Z., Gao, Y., Xin, X., Wang, L., & Song, L. (2023). A Cartilage Matrix Protein Regulates Collagen Synthesis in Mantle of Magallana gigas (Crassostrea gigas) under Ocean Acidification. Fishes, 8(6), 290. https://doi.org/10.3390/fishes8060290