
Environmental and Genetic (vgll3) Effects on the Prevalence of Male Maturation Phenotypes in Domesticated Atlantic Salmon

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
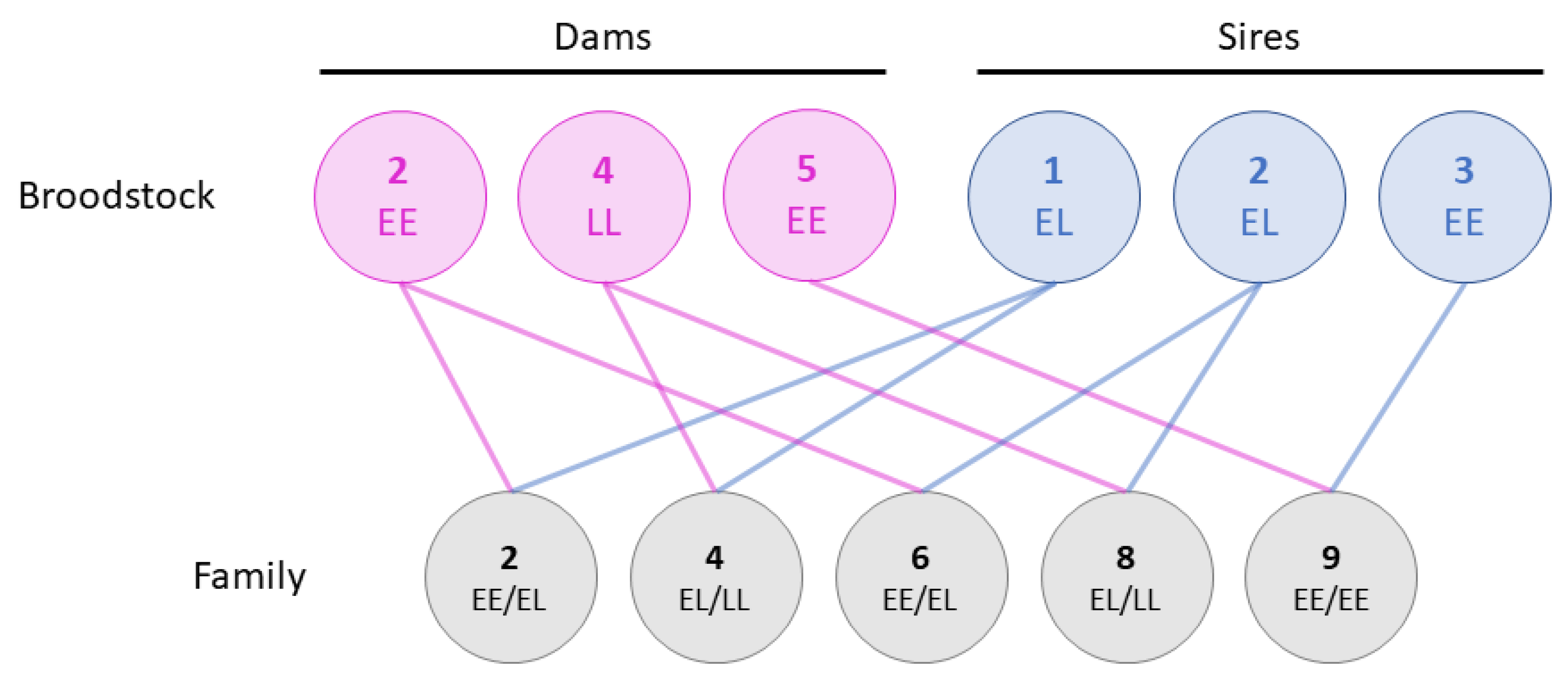
2.2. Family Material
2.3. Rearing Conditions
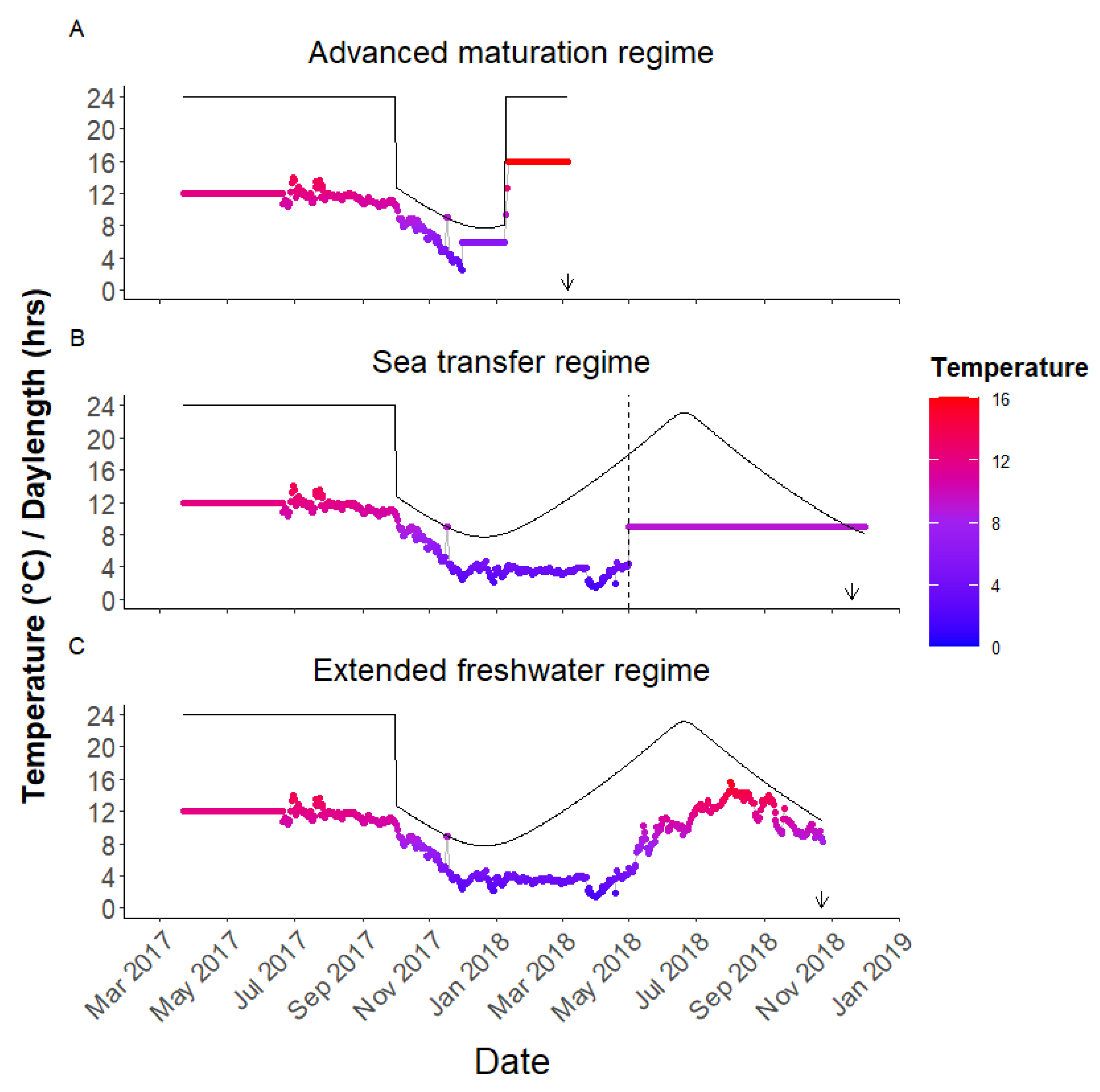
2.4. Advanced Maturation Regime
2.5. Sea Transfer Regime
2.6. Extended Freshwater Regime
2.7. Vgll3 Genotyping
2.8. Phenotypic Females
2.9. Statistical Analysis
3. Results
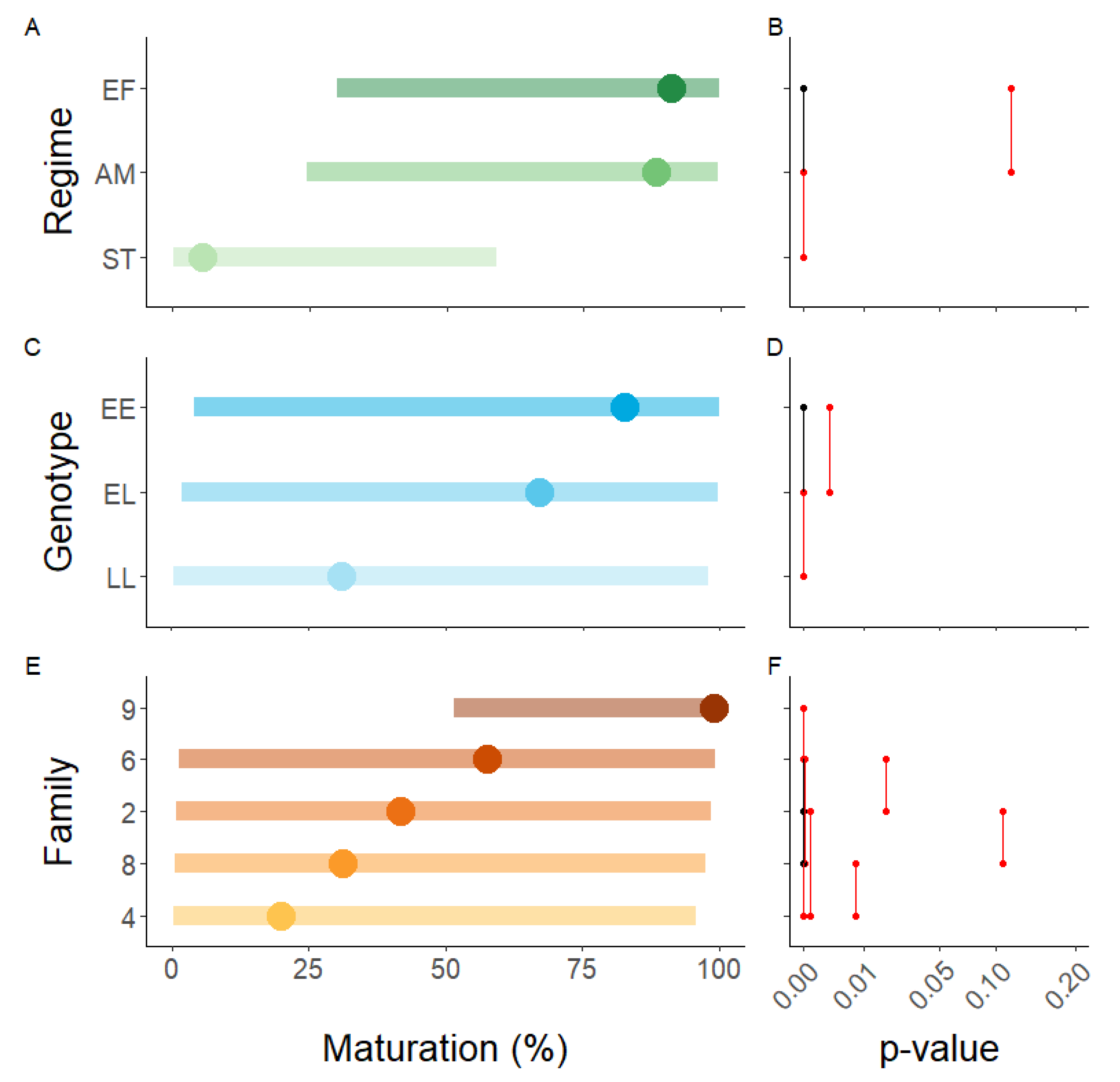
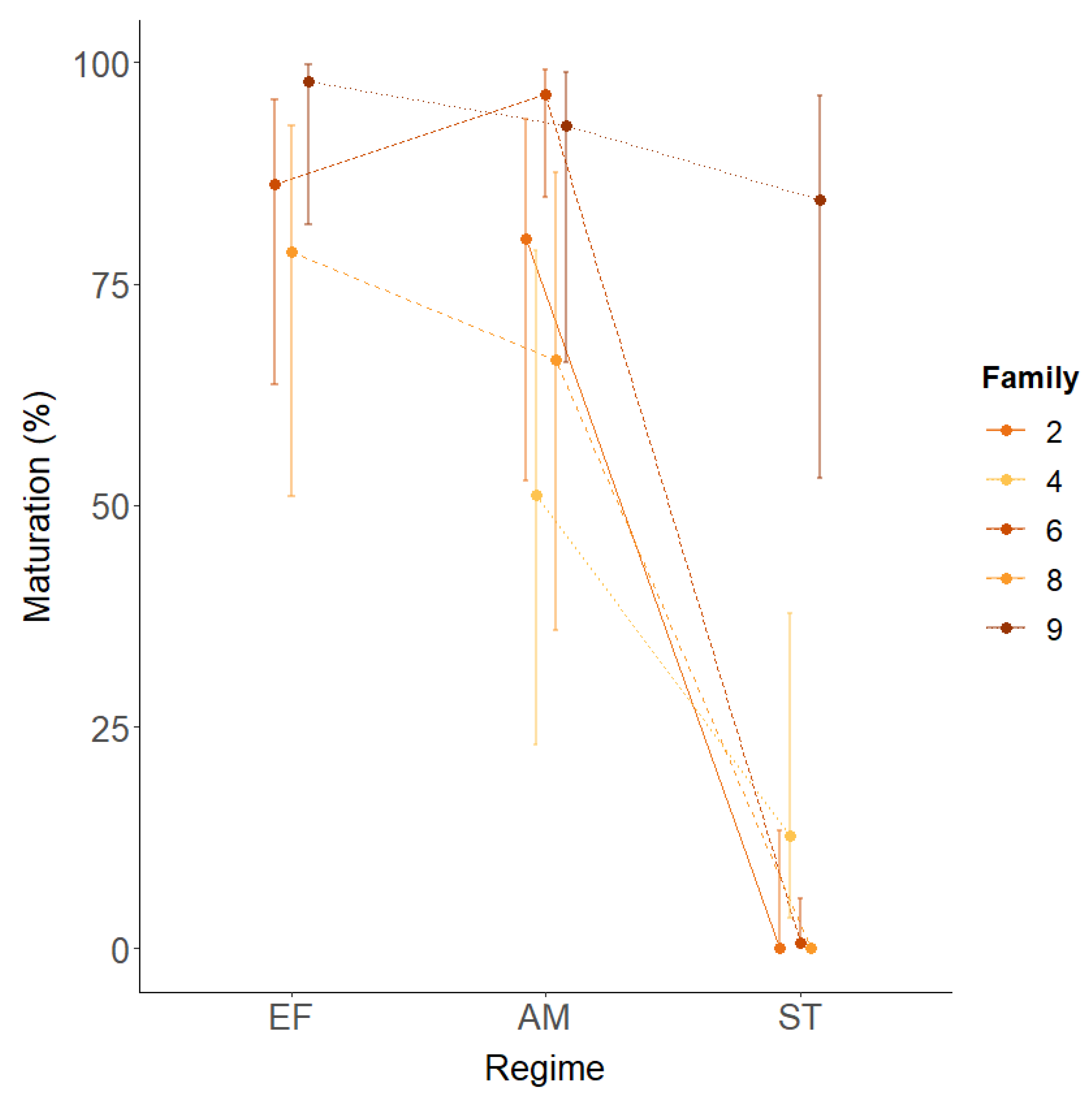
3.1. Post-Smolt Maturation across Three Environmental Regimes
3.2. Maturation over Time in Seawater
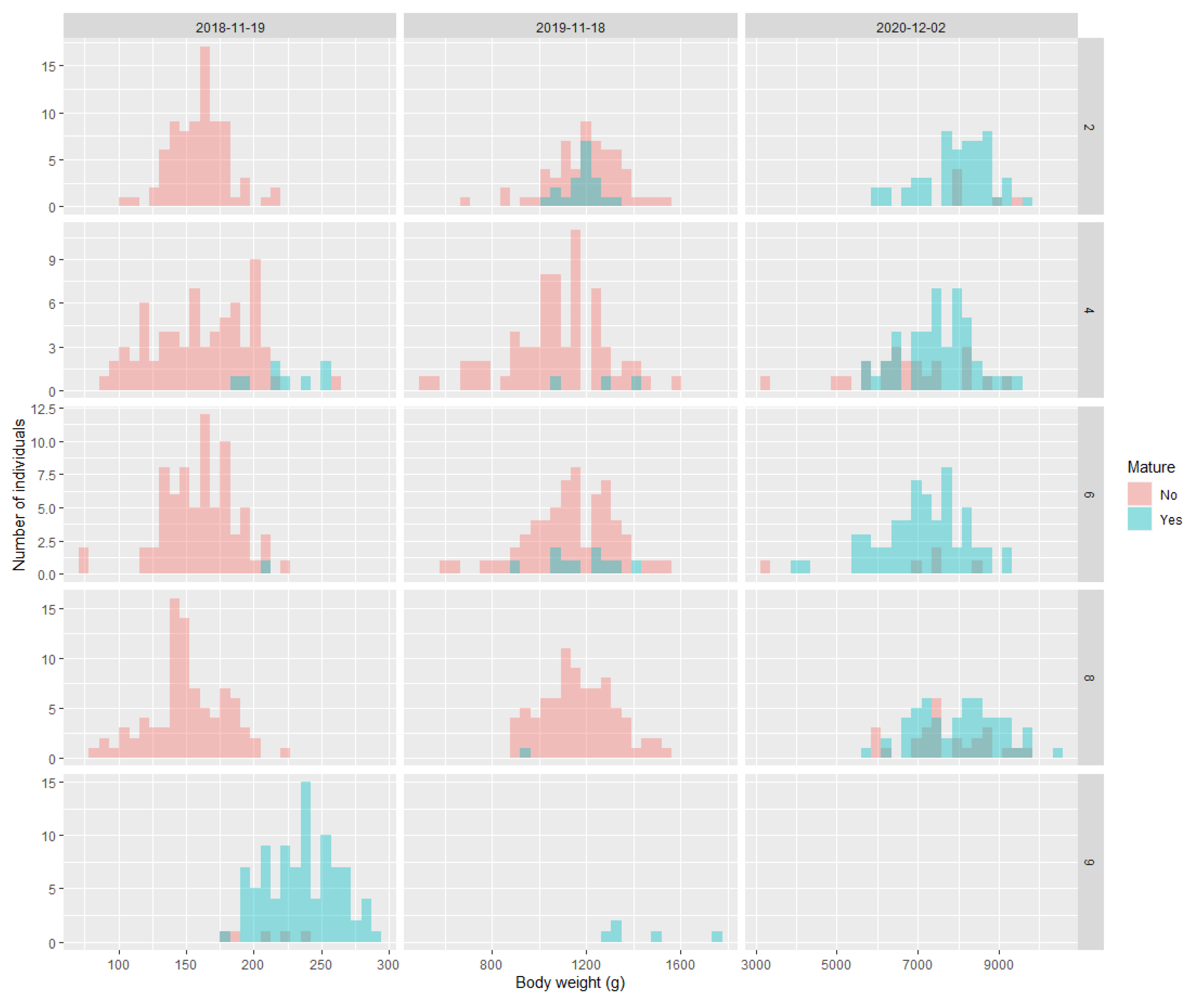
3.3. Timing of Maturation and Body Size
3.4. Vgll3 Genotype and Body Size
3.5. Genetic Sex of Phenotypic Females within Supposed “All-Male” Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO [FISHSTAT]. License: CC BY-NC-SA 3.0 IGO. Available online: https://www.fao.org/fishery/en/collection/aquaculture?lang=en (accessed on 23 January 2023).
- Taranger, G.L.; Carrillo, M.; Schulz, R.W.; Fontaine, P.; Zanuy, S.; Felip, A.; Weltzien, F.A.; Dufour, S.; Karlsen, Ø.; Norberg, B.; et al. Control of puberty in fish. Gen. Comp. Endocrinol. 2010, 165, 483–515. [Google Scholar] [CrossRef] [PubMed]
- Aksnes, A.; Gjerde, B.; Roald, S.O. Biological, chemical and organoleptic changes during maturation of farmed Atlantic salmon. Salmo Salar. Aquac. 1986, 53, 7–20. [Google Scholar] [CrossRef]
- Qui, D.; Xu, S.; Song, C.; Chi, L.; Li, X.; Sun, G.; Liu, Y. Effects of spectral composition, photoperiod and light intensity on the gonadal development of Atlantic salmon Salmo salar in recirculating aquaculture systems (RAS). Chin. J. Oceanol. Limnol. 2015, 33, 45–56. [Google Scholar]
- Bjørndal, T.; Tusvik, A. Economic analysis of on-growing of salmon post-smolts. Aquac. Econ. Manag. 2020, 24, 355–386. [Google Scholar] [CrossRef]
- Stefansson, S.O.; Hansen, T.J.; Taranger, G.L. Growth and parr-smolt transformation of Atlantic salmon (Salmo salar L.) under different light intensities and subsequent survival and growth in seawater. Aquac. Eng. 1993, 13, 231–243. [Google Scholar] [CrossRef]
- Klemetsen, A.; Amundsen, P.A.; Dempson, J.B.; Jonsson, B.; Jonsson, N.; O’Connell, M.F.; Mortensen, E. Atlantic salmon Salmo salar L., brown trout Salmo trutta L. and Arctic charr Salvelinus alpinus (L.): A review of aspects of their life histories. Ecol. Freshw. Fish 2003, 12, 1–59. [Google Scholar] [CrossRef]
- Crouse, C.; Davidson, J.; May, T.; Summerfelt, S.; Good, C. Production of market size European strain Atlantic salmon (Salmo salar) in land-based freshwater closed containment aquaculture systems. Aquac. Eng. 2021, 92, 102138. [Google Scholar] [CrossRef]
- Fjelldal, P.G.; Schulz, R.; Nilsen, T.O.; Andersson, E.; Norberg, B.; Hansen, T.J. Sexual maturation and smoltification in domesticated Atlantic salmon (Salmo salar L.)—Is there a developmental conflict? Physiol. Rep. 2018, 6, e13809. [Google Scholar] [CrossRef]
- Crouse, C.; Davidson, J.; Good, C. The effects of two water temperature regimes on Atlantic salmon (Salmo salar) growth performance and maturation in freshwater recirculating aquaculture systems. Aquaculture 2022, 553, 738063. [Google Scholar] [CrossRef]
- Martinez, E.P.; Balseiro, P.; Pedrosa, C.; Haugen, T.S.; Fleming, M.S.; Handeland, S.O. The effect of photoperiod manipulation on Atlantic salmon growth, smoltification and sexual maturation: A case study of a commercial RAS. Aquac. Res. 2021, 52, 2593–2608. [Google Scholar] [CrossRef]
- Ytrestøyl, T.; Hjelle, E.; Kolarevic, J.; Takle, H.; Rebl, A.; Afanasyev, S.; Krasnov, A.; Brunsvik, P.; Terjesen, B.F. Photoperiod in recirculation aquaculture systems and timing of seawater transfer affect seawater growth performance of Atlantic salmon (Salmo salar). J. World Aquac. Soc. 2023, 54, 73–95. [Google Scholar] [CrossRef]
- Good, C.; Davidson, J.; Iwanowicz, L.; Meyer, M.; Dietze, J.; Kolpin, D.W.; Marancik, D.; Birkett, J.; Williams, C.; Summerfelt, S. Investigating the influence of nitrate nitrogen on post-smolt Atlantic salmon Salmo salar reproductive physiology in freshwater recirculation aquaculture systems. Aquac. Eng. 2017, 78, 2–8. [Google Scholar] [CrossRef]
- Fjelldal, P.G.; Hansen, T.; Huang, T.-S. Continuous light and elevated temperature can trigger maturation both during and immediately after smoltification in male Atlantic salmon (Salmo salar). Aquaculture 2011, 321, 93–100. [Google Scholar] [CrossRef]
- Duston, J.; Saunders, R.L. Life histories of Atlantic salmon altered by winter temperature and summer rearing in fresh- or sea-water. Environ. Biol. Fishes 1997, 50, 149–166. [Google Scholar] [CrossRef]
- Atkinson, D. Temperature and organism size: A biological law for ectotherms? Adv. Ecol. Res. 1994, 25, 1–58. [Google Scholar]
- Fraser, T.W.K.; Hansen, T.; Skjæraasen, J.E.; Mayer, I.; Sambraus, F.; Fjelldal, P.G. The effect of triploidy on the culture performance, deformity prevalence, and heart morphology in Atlantic salmon. Aquaculture 2013, 416–417, 255–264. [Google Scholar] [CrossRef]
- Falconer, L.; Hjøllo, S.S.; Telfer, T.C.; McAdam, B.J.; Hermansen, Ø.; Ytteborg, E. The importance of calibrating climate change projections to local conditions at aquaculture sites. Aquaculture 2020, 514, 734487. [Google Scholar] [CrossRef]
- Nævdal, G.; Holm, M.; Ingebrigtsen, O.; Møller, D. Variation in age at fist spawning in Atlantic salmon (Salmo salar). J. Fish. Board Can. 1978, 35, 145–147. [Google Scholar] [CrossRef]
- Barson, N.J.; Aykanat, T.; Hindar, K.; Baranski, M.; Bolstad, G.H.; Fiske, P.; Jacq, C.; Jensen, A.J.; Johnston, S.E.; Karlsson, S.; et al. Sex-dependent dominance at a single locus maintains variation in age at maturity in salmon. Nature 2015, 528, 405–408. [Google Scholar] [CrossRef]
- Debes, P.V.; Piavchenko, N.; Ruokolainen, A.; Ovaskainen, O.; Moustakas-Verho, J.E.; Parre, N.; Aykanat, T.; Erkinaro, J.; Primmer, C.R. Polygenic and major-locus contributions to sexual maturation timing in Atlantic salmon. Mol. Ecol. 2021, 30, 4505–4519. [Google Scholar] [CrossRef]
- Fjelldal, P.G.; Hansen, T.J.; Wargelius, A.; Ayllon, F.; Glover, K.A.; Schulz, R.W.; Fraser, T.W.K. Development of supermale and all-male Atlantic salmon to research the vgll3 allele-puberty link. BMC Genet. 2020, 21, 123. [Google Scholar] [CrossRef]
- Ayllon, F.; Kjærner-Semb, E.; Furmanek, T.; Wennevik, V.; Solberg, M.F.; Dahle, G.; Taranger, G.L.; Glover, K.A.; Almén, S.; Rubin, C.J.; et al. The vgll3 locus controls age at maturity in wild and domesticated Atlantic salmon (Salmo salar L.) males. PLoS Genet. 2015, 11, e1005628. [Google Scholar] [CrossRef]
- Ayllon, F.; Solberg, M.F.; Glover, K.A.; Mohammadi, F.; Kjærner-Semb, E.; Fjelldal, P.G.; Andersson, E.; Hansen, T.; Edvardsen, E.; Wargelius, A. The influence of vgll3 genotypes on sea age at maturity is altered in farmed mowi strain Atlantic salmon. BMC Genet. 2019, 20, 44. [Google Scholar] [CrossRef]
- Crespo, D.; Skaftnesmo, K.O.; Kjærner-Semb, E.; Yilmaz, O.; Norberg, B.; Olausson, S.; Vogelsang, P.; Bogerd, J.; Kleppe, L.; Edvardsen, R.B.; et al. Pituitary gonadotropin gene expression during induced onset of postsmolt maturation in male Atlantic salmon: In vivo and tissue culture studies. Front. Endocrinol. 2022, 13, 826920. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, H.; Fridberg, G. Sexual maturation versus immaturity: Different tactics with adaptive values in Baltic salmon (Salmo salar L.) male smolts. Can. J. Zool. 1982, 60, 1822–1827. [Google Scholar] [CrossRef]
- Hansen, T.J.; Penman, D.; Glover, K.A.; Fraser, T.W.K.; Vågseth, T.; Thorsen, A.; Sørvik, A.G.E.; Fjelldal, P.G. Production and verification of the first Atlantic salmon (Salmo salar L.) clonal lines. BMC Genet. 2020, 21, 71. [Google Scholar] [CrossRef]
- Quillet, E.; Aubard, G.; Queau, I. Mutation in a sex-determining gene in rainbow trout: Detection and genetic analysis. J. Hered. 2002, 93, 91–99. [Google Scholar] [CrossRef]
- Komen, J.; De Boer, P.; Richter, C. Male sex reversal in gynogenetic XX females of common carp (Cyprinus carpio L.) by a recessive mutation in a sex-determining gene. J. Hered. 1992, 83, 431–434. [Google Scholar] [CrossRef]
- Mair, G.; Abucay, J.; Abella, T.; Beardmore, J.; Skibinski, D. Genetic manipulation of sex ratio for the large-scale production of all-male tilapia Oreochromis niloticus. Can. J. Fish Aquat. Sci. 1997, 54, 396–404. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 February 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1–155; R Core Team: Vienna, Austria, 2022; Available online: https://CRAN.R-project.org/package=nlme (accessed on 15 February 2021).
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means, R Package Version 1.7.0; R Core Team: Vienna, Austria, 2021; Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 February 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978–3–319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 15 February 2021).
- Duston, J.; Saunders, R.L. Effect of 6-, 12-, and 18-month photoperiod cycles on smolting and sexual maturation in juvenile Atlantic Salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 1992, 49, 2273–2280. [Google Scholar] [CrossRef]
- Duston, J. Effect of salinity on survival and growth of Atlantic salmon (Salmo salar) parr and smolts. Aquaculture 1994, 121, 115–124. [Google Scholar] [CrossRef]
- Thorpe, J.E. Maturation responses of salmonids to changing developmental opportunities. Mar. Ecol. Prog. Ser. 2007, 335, 285–288. [Google Scholar] [CrossRef]
- Thrush, M.A.; Duncan, N.J.; Bromage, N.R. The use of photoperiod in the production of out-of-season Atlantic salmon (Salmo salar) smolts. Aquaculture 1994, 121, 29–44. [Google Scholar] [CrossRef]
- Fraser, T.W.K.; Hansen, T.J.; Norberg, B.; Nilsen, T.O.; Schulz, R.W.; Fjelldal, P.G. Atlantic salmon male post-smolt maturation can be reduced by using a 3-hour scotophase when inducing smoltification. Aquaculture 2023, 562, 738772. [Google Scholar] [CrossRef]
- Fjelldal, P.G.; Fraser, T.W.K.; Hansen, T.J.; Karlsen, Ø.; Bui, S. Effects of laboratory salmon louse infection on mortality, growth and sexual maturation in Atlantic salmon. ICES J. Mar. Sci. 2022, 79, 1530–1583. [Google Scholar] [CrossRef]
- Lundqvist, H.; Borg, B.; Berglund, I. Androgens impair seawater adaptability in smolting Baltic salmon (Salmo salar). Can. J. Zool. 1989, 67, 1733–1736. [Google Scholar] [CrossRef]
- Melo, M.C.; Andersson, E.; Fjelldal, P.G.; Bogerd, J.; França, L.R.; Taranger, G.L.; Schulz, R.W. Salinity and photoperiod modulate pubertal development in Atlantic salmon (Salmo salar). J. Endocrinol. 2014, 220, 319–332. [Google Scholar] [CrossRef]
- Mohamed, A.R.; Verbyla, K.L.; Al-Mamun, H.A.; McWilliam, S.; Evans, B.; King, H.; Kube, P.; Kijas, J.W. Polygenic and sex specific architecture for two maturation traits in farmed Atlantic salmon. BMC Genom. 2019, 20, 139. [Google Scholar] [CrossRef]
- Bohlin, T.; Dellefors, C.; Faremo, U. Probability of first sexual maturation of male parr in wild sea-run brown trout (Salmo trutta) depends on condition factor 1 yr in advance. Can. J. Fish. Aquat. Sci. 1994, 51, 1920–1926. [Google Scholar] [CrossRef]
- Herbinger, C.M.; Friars, G.W. Correlation between condition factor and total lipid content in Atlantic salmon, Salmo salar L., parr. Aquac. Fish. Manag. 1991, 22, 527–529. [Google Scholar] [CrossRef]
- Jonsson, N.; Jonsson, B.; Hansen, L.P. Changes in proximate composition and estimates of energetic costs during upstream migration and spawning in Atlantic salmon Salmo salar. J. Anim. Ecol. 1997, 66, 425–436. [Google Scholar] [CrossRef]
- Trudel, M.; Tucker, S.; Morris, J.F.T.; Higgs, D.A.; Welch, D.W. Indicators of energetic status in juvenile coho salmon and chinook salmon. North Am. J. Fish. Manag. 2004, 25, 374–390. [Google Scholar] [CrossRef]
- Andersson, E.; Schulz, R.W.; Male, R.; Bogerd, J.; Patiña, P.; Benedet, S.; Norberg, B.; Taranger, G.L. Pituitary gonadotropin and ovarian gonadotropin receptor transcript levels: Seasonal and photoperiod-induced changes in the reproductive physiology of female Atlantic salmon (Salmo salar). Gen. Comp. Endocrinol. 2013, 191, 247–258. [Google Scholar] [CrossRef]
- Fraser, T.W.K.; Fjelldal, P.G.; Schulz, R.W.; Norberg, B.; Hansen, T.J. Termination of puberty in out-of-season male Atlantic salmon smolts. Comp. Biochem. Physiol. Part A 2019, 232, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Ahi, E.P.; Sinclair-Waters, M.; Moustakas-Verho, J.; Jansouz, S.; Primmer, C.R. Strong regulatory effects of vgll3 genotype on reproductive axis gene expression in juvenile male Atlantic salmon. Gen. Comp. Endocrinol. 2022, 325, 114055. [Google Scholar] [CrossRef]
- Åsheim, E.R.; Debes, P.V.; House, A.; Liljeström, P.; Niemelä, P.T.; Siren, P.T.; Erkinaro, J.; Primmer, C.R. Atlantic salmon (Salmo salar) age at maturity is strongly affected by temperature, population and age-at-maturity genotype. Conserv. Physiol. 2023, 11, coac086. [Google Scholar] [CrossRef]
- Barrett, L.T.; Oldham, T.; Kristiansen, T.S.; Oppedal, F.; Stien, L.H. Declining size-at-harvest in Norwegian salmon aquaculture: Lice, disease, and the role of stunboats. Aquaculture 2022, 559, 738440. [Google Scholar] [CrossRef]
- Borgstrøm, R.; Opdahl, J.; Svenning, M.-A.; Länsman, M.; Orell, P.; Niemelä, E.; Erkinaro, J.; Dempson, J.B. Temporal changes in ascendance and in-season exploitation of Atlantic salmon, Salmo salar, inferred by a video camera array. Fish. Manag. Ecol. 2010, 17, 454–463. [Google Scholar] [CrossRef]
- Besnier, F.; Ayllon, F.; Skaala, Ø.; Favnbøe, M.F.; Fjeldheim, P.T.; Anderson, K.; Knutar, S.; Glover, K.A. Introgression of domesticated salmon changes life history and phenology of a wild salmon population. Evol. Appl. 2022, 15, 853–864. [Google Scholar] [CrossRef]
- Besnier, F.; Skaala, Ø.; Wennevik, V.; Ayllon, A.; Utne, R.; Fjeldheim, P.T.; Andersen-Fjeldheim, K.; Knutar, S.; Glover, K.A. Overruled by nature: A plastic response to an ecological regime shift disconnects a gene and its trait. Mol. Ecol. 2022, in press. [Google Scholar]
- Strøm, J.F.; Thorstad, E.B.; Rikardsen, A.H. Thermal habitat of adult Atlantic salmon Salmo salar in a warming ocean. J. Fish Biol. 2020, 96, 327–336. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Model | Maturation (%) | Odds Ratio | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | df | p | EF | AM | ST | EF/AM | EF/ST | AM/ST | ||
| 2 | - | - | - | 91.6 (91.5–91.7) | - | 0.0 | - | - | - | |
| 4 | 18.5 | 1 | <0.001 | *** | 39.4 (31.1–48.3) a | - | 10.1 (4.9–19.6) b | - | - | 5.8 (2.4) |
| 6 | 50.2 | 2 | <0.001 | *** | 91.3 (87.7–94.5) b | 97.7 (93.8–99.2) a | 1.3 (0.2–9.1) c | 0.2 (0.1) | 766.8 (796.8) | 3087.6 (3508.7) |
| 8 | 7.0 | 1 | 0.008 | ** | 67.8 (33.4–89.8) a | 52.2 (20.7–82.1) b | 0.0 | 1.9 (0.5) | - | - |
| 9 | 5.3 | 2 | 0.071 | 99.4 (95.6–99.9) a | 97.8 (91.5–99.4) b | 94.8 (88.1–97.8) c | 3.5 (4.4) | 8.5 (9.3) | 2.4 (2.0) | |
| Year | Family | Model | Maturation (%) | Odds Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | df | p | EE | EL | LL | EE/EL | EL/LL | |||
| 2018 | 2 | 17.6 | 1 | <0.001 | *** | 97.5 (90.7–99.4) a | 77.7 (68.1–85.0) b | - | 11.4 (8.6) | - |
| 4 | 2.7 | 1 | 0.097 | - | 25.5 (8.7–55.2) | 17.3 (5.3–44.1) | - | 1.6 (0.5) | ||
| 6 | 2.4 | 1 | 0.124 | 72.0 (3.8–99.4) | 57.9 (2.1–98.9) | - | 1.9 (0.8) | - | ||
| 8 | 65.1 | 1 | <0.001 | *** | - | 80.8 (71.0–87.8) a | 35.6 (25.2–47.7)b | - | 7.6 (1.9) | |
| 2019 | 2 | 8.5 | 1 | 0.004 | ** | 38.5 (24.7–54.4) a | 10.5 (4.0–24.9) b | - | 5.3 (3.3) | - |
| 4 | - | - | - | - | 6.5 (2.1–18.4) | 0.0 | - | - | ||
| 6 | 6.47 | 1 | 0.011 | * | 22.5 (12.1–37.9) a | 3.1 (0.4–19.1)b | - | 9.0 (9.8) | - | |
| 8 | - | - | - | - | 2.3 (0.3–14.4) | 0.0 | - | - | ||
| 2020 | 2 | 1.87 | 1 | 0.171 | 95.8 (75.7–99.4) | 85.3 (69.2–93.7) | - | 4.0 (4.5) | - | |
| 4 | 24.9 | 1 | <0.001 | *** | - | 88.4 (74.9–95.1) a | 29.2 (14.6–49.8) b | - | 18.5 (12.1) | |
| 6 | 2.09 | 1 | 0.149 | 96.8 (80.4–99.5) | 87.1 (70.3–95.1) | - | 4.4 (5.1) | - | ||
| 8 | - | - | - | - | 100.0 | 35.9 (22.5–51.9) | - | - | ||
| Phenotype | Family | Model | Delayed from Earlier to Later Phenotype (%) | Odds Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | df | p | EE | EL | LL | EE/EL | EL/LL | |||
| 0 SW to 1 SW | 2 | - | - | - | 100.0 | 100.0 | - | - | - | |
| 4 | - | - | - | - | 33.3 (11.1–66.7) | 0.0 | - | - | ||
| 6 | - | - | - | 100.0 | 35.3 (5.9–94.1) | - | - | - | ||
| 8 | - | - | - | - | 100.0 | - | - | - | ||
| 1 SW to 2 SW | 2 | 7.1242 | 1 | 0.008 | ** | 60.5 (44.4–74.6) | 87.9 (71.8–95.4) | - | 0.21 (0.13) | - |
| 4 | 0.97931 | 1 | 0.322 | - | 92.6 (79.6–97.6) | 100.0 | - | - | ||
| 6 | 5.7051 | 1 | 0.017 | * | 76.9 (61.3–7.5) | 96.4 (78.6–99.5) | - | 0.12 (0.13) | - | |
| 8 | 0.55807 | 1 | 0.455 | - | 97.7 (85.6–99.6) | 100.0 | - | - | ||
| 2 SW to 2 SW+ | 2 | 1.8721 | 1 | 0.171 | 4.2 (0.6–24.3) | 14.7 (6.3–30.8) | - | 0.25 (0.29) | - | |
| 4 | 24.936 | 1 | <0.001 | *** | - | 11.6 (4.9–25.1) | 70.8 (50.2–85.4) | - | 0.05 (0.04) | |
| 6 | 2.0854 | 1 | 0.149 | 3.2 (0.5–19.6) | 12.9 (5.0–29.7) | - | 0.23 (0.26) | - | ||
| 8 | - | - | - | - | 0.0 | 64.1 (48.1–77.5) | - | - | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraser, T.W.K.; Hansen, T.J.; Fjelldal, P.G. Environmental and Genetic (vgll3) Effects on the Prevalence of Male Maturation Phenotypes in Domesticated Atlantic Salmon. Fishes 2023, 8, 275. https://doi.org/10.3390/fishes8050275
Fraser TWK, Hansen TJ, Fjelldal PG. Environmental and Genetic (vgll3) Effects on the Prevalence of Male Maturation Phenotypes in Domesticated Atlantic Salmon. Fishes. 2023; 8(5):275. https://doi.org/10.3390/fishes8050275
Chicago/Turabian StyleFraser, Thomas W. K., Tom J. Hansen, and Per Gunnar Fjelldal. 2023. "Environmental and Genetic (vgll3) Effects on the Prevalence of Male Maturation Phenotypes in Domesticated Atlantic Salmon" Fishes 8, no. 5: 275. https://doi.org/10.3390/fishes8050275
APA StyleFraser, T. W. K., Hansen, T. J., & Fjelldal, P. G. (2023). Environmental and Genetic (vgll3) Effects on the Prevalence of Male Maturation Phenotypes in Domesticated Atlantic Salmon. Fishes, 8(5), 275. https://doi.org/10.3390/fishes8050275