
Piscine Orthoreovirus-1 (PRV-1) Has Been Present in Chilean Salmon Aquaculture since at Least 1994
 ,
,
Abstract
1. Introduction
2. Materials and Methodology
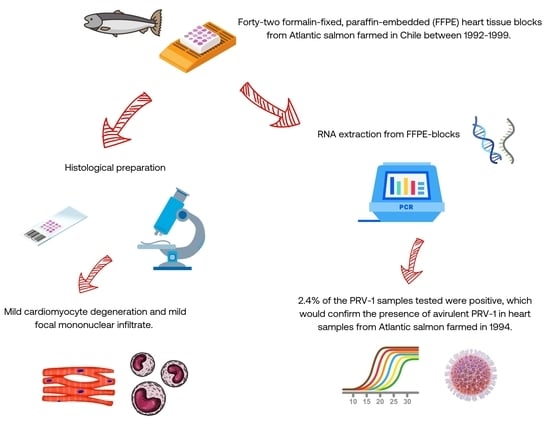
2.1. Fish and Tissue Samples
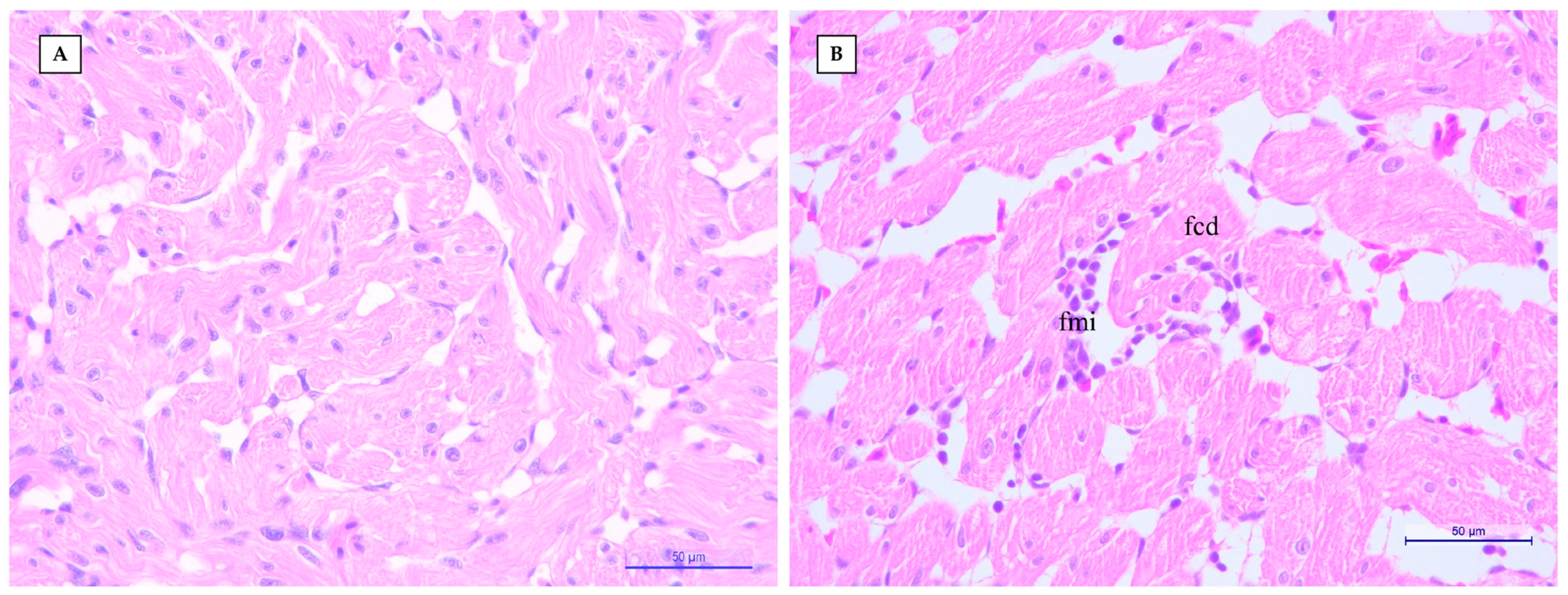
2.2. Histopathological and Immunohistochemical (IHC) Examination
2.3. RNA Extraction
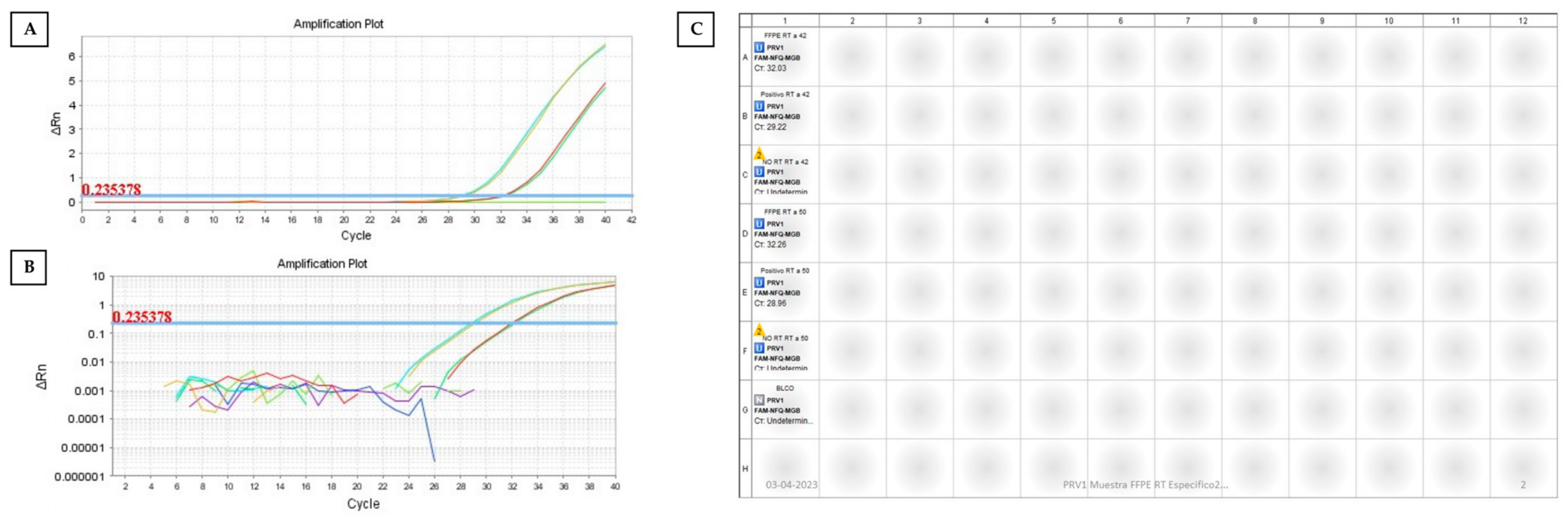
2.4. One-Step RT–qPCR
2.5. Two-Step RT–qPCR
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kongtorp, R.T.; Kjerstad, A.; Taksdal, T.; Guttvik, A.; Falk, K. Heart and skeletal muscle inflammation in Atlantic salmon, Salmo salar L.: A new infectious disease. J. Fish Dis. 2004, 27, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.-T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and Skeletal Muscle Inflammation of Farmed Salmon Is Associated with Infection with a Novel Reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, E.; Ferguson, H.W.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; Vanderstichel, R.; Wessel, Ø.; Rimstad, E.; Gardner, I.A.; Hammell, K.L.; et al. Heart and skeletal muscle inflammation (HSMI) disease diagnosed on a British Columbia salmon farm through a longitudinal farm study. PLoS ONE 2017, 12, e0171471. [Google Scholar]
- Godoy, M.G.; Kibenge, M.J.T.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S.B. First description of clinical presentation of piscine orthoreovirus (PRV) infections in salmonid aquaculture in Chile and identification of a second genotype (Genotype II) of PRV. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-Genome Sequencing and Confirmation of the Causative Agent of Erythrocytic Inclusion Body Syndrome in Coho Salmon Identifies a New Type of Piscine Orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [PubMed]
- Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, Ø.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Dahle, M.K.; Rimstad, E. Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses 2018, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, R.; Stoeckle, B.C.; Young, M.; Popp, L.; Taeubert, J.-E.; Pfaffl, M.W.; Geist, J. Identification of a piscine reovirus-related pathogen in proliferative darkening syndrome (PDS) infected brown trout (Salmo trutta fario) using a next-generation technology detection pipeline. PLoS ONE 2018, 13, e0206164. [Google Scholar] [CrossRef] [PubMed]
- Dhamotharan, K.; Tengs, T.; Wessel, Ø.; Braaen, S.; Nyman, I.B.; Hansen, E.F.; Christiansen, D.H.; Dahle, M.K.; Rimstad, E.; Markussen, T. Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses 2019, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Wessel, Ø.; Hansen, E.F.; Dahle, M.K.; Alarcon, M.; Vatne, N.A.; Nyman, I.B.; Soleim, K.B.; Dhamotharan, K.; Timmerhaus, G.; Markussen, T.; et al. Piscine Orthoreovirus-1 Isolates Differ in Their Ability to Induce Heart and Skeletal Muscle Inflammation in Atlantic Salmon (Salmo salar). Pathogens 2020, 9, 1050. [Google Scholar] [CrossRef] [PubMed]
- Bustos, P.; Rozas-Serri, M.; Bohle, H.; Ildefonso, R.; Sandoval, A.; Gaete, A.; Araya, C.; Grothusen, H.; Tapia, E.; Gallardo, A.; et al. Primer reporte de piscine reovirus en salmón del Atlántico, Salmo salar, cultivado en Chile. Versión Difer. 2011, 65–69. [Google Scholar]
- Sernapesca. Informe Sanitario De La Salmonicultura En Centros Agua Dulce y Mar Año 2021; Animal DdS: Valparaíso, Chile, 2022; p. 52. [Google Scholar]
- Dahle, M.K.; Wessel, Ø.; Rimstad, E. Immune Response Against Piscine orthoreovirus (PRV) in Salmonids. Princ. Fish Immunol. 2022, 445–461. [Google Scholar] [CrossRef]
- Haatveit, H.M.; Nyman, I.B.; Markussen, T.; Wessel, Ø.; Dahle, M.K.; Rimstad, E. The non-structural protein μNS of piscine orthoreovirus (PRV) forms viral factory-like structures. Veter. Res. 2016, 47, 5. [Google Scholar] [CrossRef] [PubMed]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Løvoll, M.; Finstad, Ø.W.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, S.; Robertsen, B.; Rimstad, E. Correction: Sequence Analysis of the Genome of Piscine Orthoreovirus (PRV) Associated with Heart and Skeletal Muscle Inflammation (HSMI) in Atlantic Salmon (Salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef]
- Karatas, S.; Mikalsen, J.; Steinum, T.M.; Taksdal, T.; Bordevik, M.; Colquhoun, D.J. Real time PCR detection of Piscirickettsia salmonis from formalin-fixed paraffin-embedded tissues. J. Fish Dis. 2008, 31, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Snow, M.; McKay, P.; McBeath, A.J.A.; Black, J.; Doig, F.; Kerr, R.; Cunningham, C.O.; Nylund, A.; Devold, M. Development, application and validation of a Taqman real-time RT-PCR assay for the detection of infectious salmon anaemia virus (ISAV) in Atlantic salmon (Salmo salar). Dev. Biol. 2006, 126, 133–145. [Google Scholar]
- Siah, A.; Knutsen, E.; Richmond, Z.; Mills, M.; Frisch, K.; Powell, J.F.; Brevik, Ø.; Duesund, H. Real-time RT-qPCR assay to detect sequences in the Piscine orthoreovirus-1 genome segment S1 associated with heart and skeletal muscle inflammation in Atlantic salmon. J. Fish Dis. 2020, 43, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Kibenge, M.J.; Iwamoto, T.; Wang, Y.; Morton, A.; Godoy, M.G.; Kibenge, F.S. Whole-genome analysis of piscine reovirus (PRV) shows PRV represents a new genus in family Reoviridae and its genome segment S1 sequences group it into two separate sub-genotypes. Virol. J. 2013, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Kashofer, K.; Viertler, C.; Pichler, M.; Zatloukal, K. Quality Control of RNA Preservation and Extraction from Paraffin-Embedded Tissue: Implications for RT-PCR and Microarray Analysis. PLoS ONE 2013, 8, e70714. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Histoscore | Atrium | Ventricle | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Epicardium | Stratum Compactum | Stratum Sponsiosum | Epicardium | ||||||
| Cell Degeneration | Mononuclear Cells Infiltrate | Mononuclear Cells Infiltrate | Cell Degeneration | Mononuclear Cells Infiltrate | Mononuclear Cells Infiltrate | Cell Degeneration | Mononuclear Cells Infiltrate | Thrombosis | |
| 0 | NHC | NIC | NIC | NIC | NHC | NIC | NHC | NHC | NHC |
| 1 | FCD (<10% TS) | MiMI (<10% TS) | MiMI (<10% TS) | MiMI (<10% TS) | FCD (<10% TS) | MiMI (<10% TS) | FCD (<10% TS) | <10% TS | <10% TS |
| 2 | FCD (10–50% TS) | MoMI (10–50% TS) | MoMI (10–50% TS) | MoMI (10–50% TS) | FCD (10–50% TS) | MoMI (10–50% TS) | FCD (10–50% TS) | 10–50% TS | 10–50% TS |
| 3 | DCD (>50% TS) | SMI (>10% TS) | SMI (>10% TS) | SMI (>10% TS) | DCD (>50% TS) | SMI (>10% TS) | DCD (>50% TS) | >50% TS | >50% TS |
| Relative weighting | 5 | 5 | 18.3 | 13.3 | 13.3 | 13.3 | 13.3 | 16.5 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozas-Serri, M.; Ildefonso, R.; Jaramillo, V.; Peñaloza, E.; Leiva, C.; Barrientos, S.; Coñuecar, D.; Maldonado, L.; Muñoz, A.; Peña, A.; et al. Piscine Orthoreovirus-1 (PRV-1) Has Been Present in Chilean Salmon Aquaculture since at Least 1994. Fishes 2023, 8, 229. https://doi.org/10.3390/fishes8050229
Rozas-Serri M, Ildefonso R, Jaramillo V, Peñaloza E, Leiva C, Barrientos S, Coñuecar D, Maldonado L, Muñoz A, Peña A, et al. Piscine Orthoreovirus-1 (PRV-1) Has Been Present in Chilean Salmon Aquaculture since at Least 1994. Fishes. 2023; 8(5):229. https://doi.org/10.3390/fishes8050229
Chicago/Turabian StyleRozas-Serri, Marco, Ricardo Ildefonso, Victoria Jaramillo, Estefanía Peñaloza, Camila Leiva, Soraya Barrientos, Darling Coñuecar, Lucerina Maldonado, Ariel Muñoz, Andrea Peña, and et al. 2023. "Piscine Orthoreovirus-1 (PRV-1) Has Been Present in Chilean Salmon Aquaculture since at Least 1994" Fishes 8, no. 5: 229. https://doi.org/10.3390/fishes8050229
APA StyleRozas-Serri, M., Ildefonso, R., Jaramillo, V., Peñaloza, E., Leiva, C., Barrientos, S., Coñuecar, D., Maldonado, L., Muñoz, A., Peña, A., Aranis, F., & Senn, C. (2023). Piscine Orthoreovirus-1 (PRV-1) Has Been Present in Chilean Salmon Aquaculture since at Least 1994. Fishes, 8(5), 229. https://doi.org/10.3390/fishes8050229