



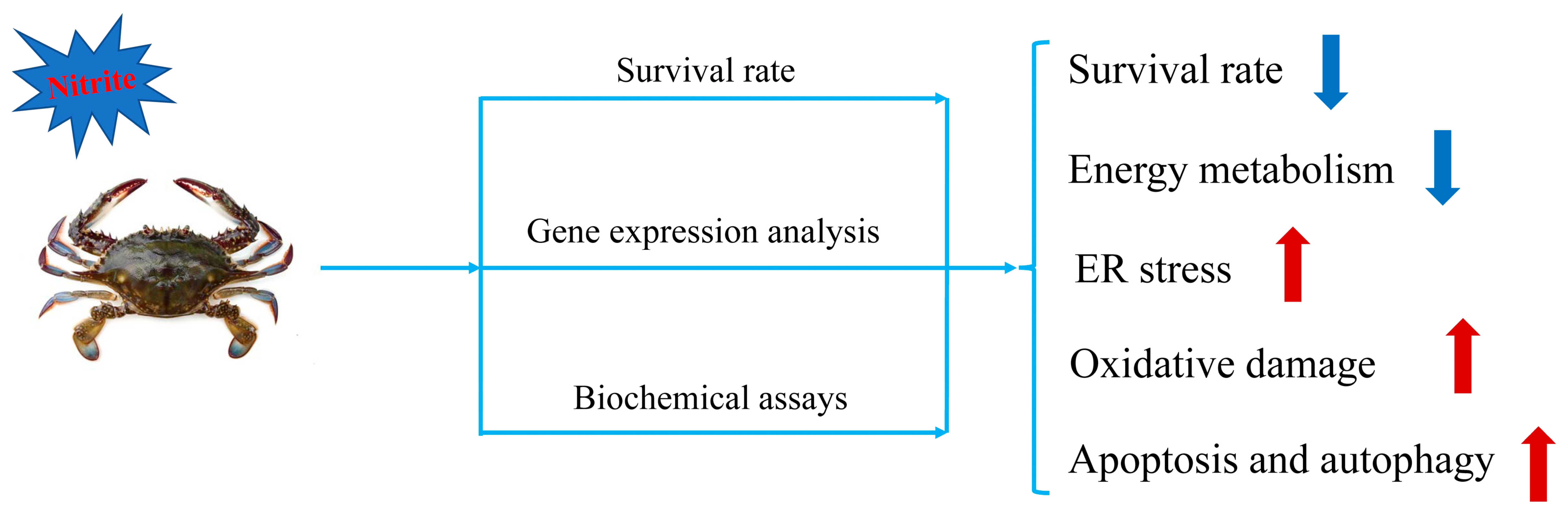
Survival, Energy Status, and Cellular Stress Responses of the Juvenile Swimming Crab Portunus trituberculatus under Acute Nitrite Stress


Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Animal
2.2. Survival Experiment
2.3. Sample Collection
2.4. Gene Expression Analysis
2.5. Biochemical Assays
2.6. Statistical Analysis
3. Results
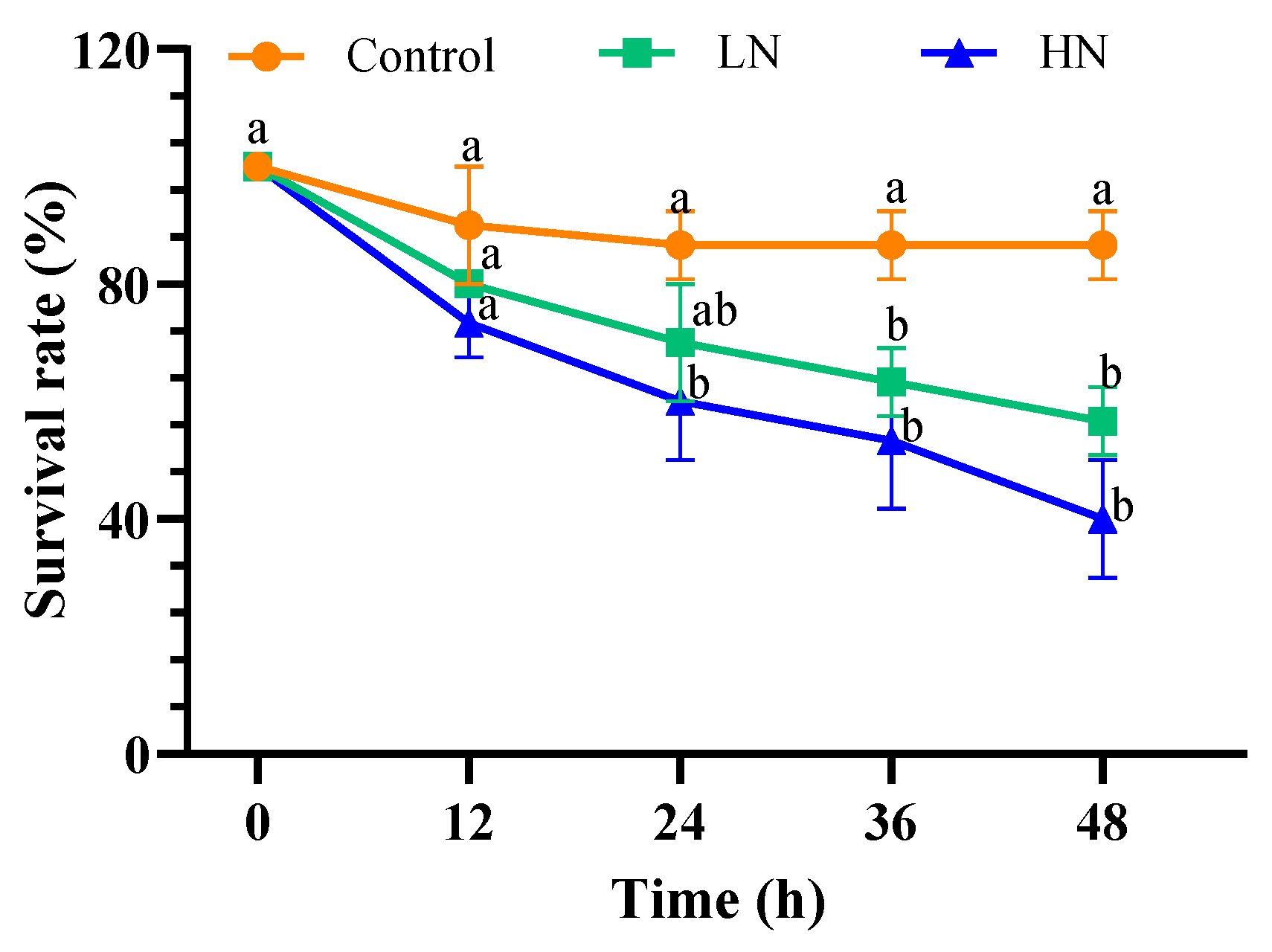
3.1. Survival Rate
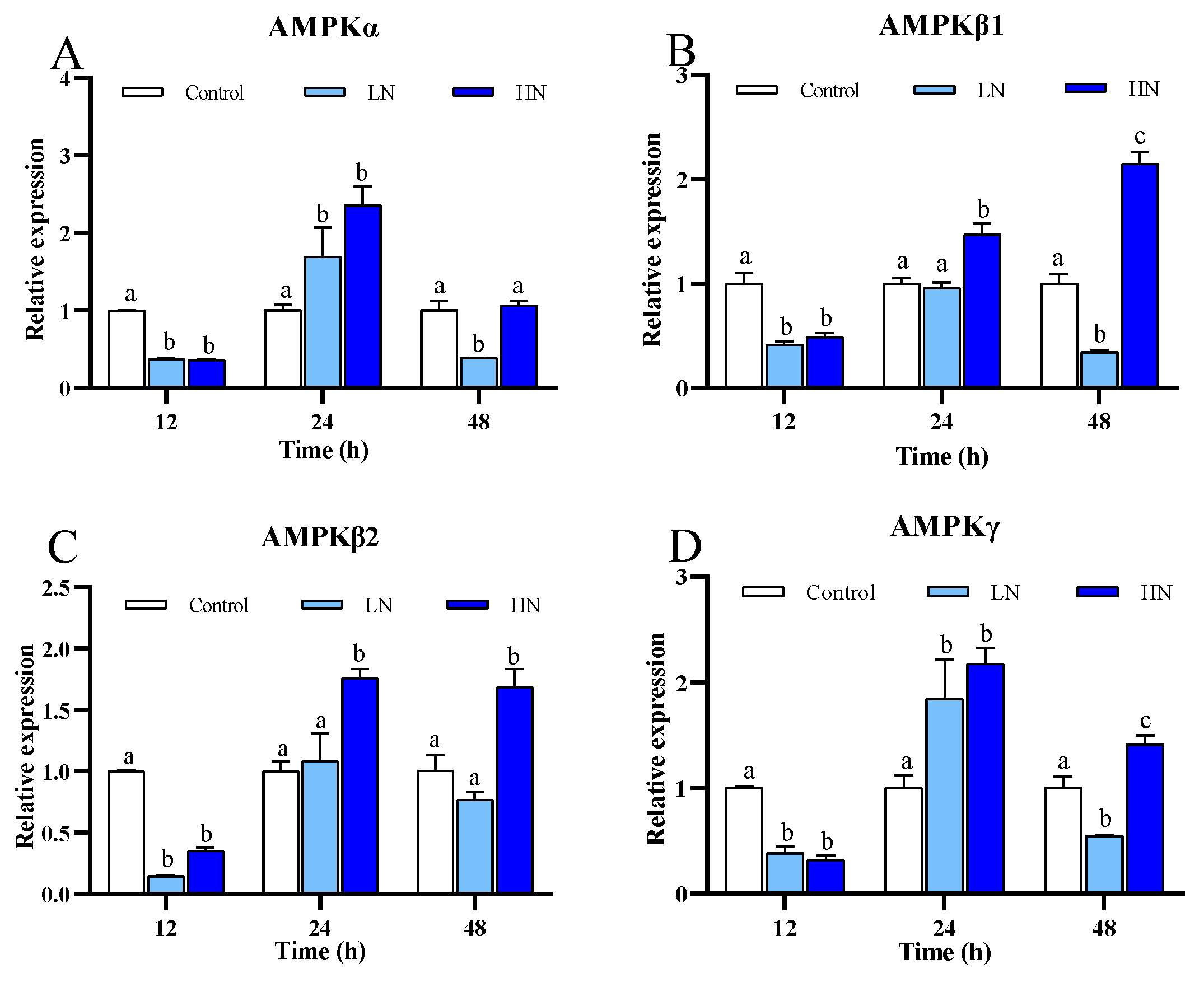
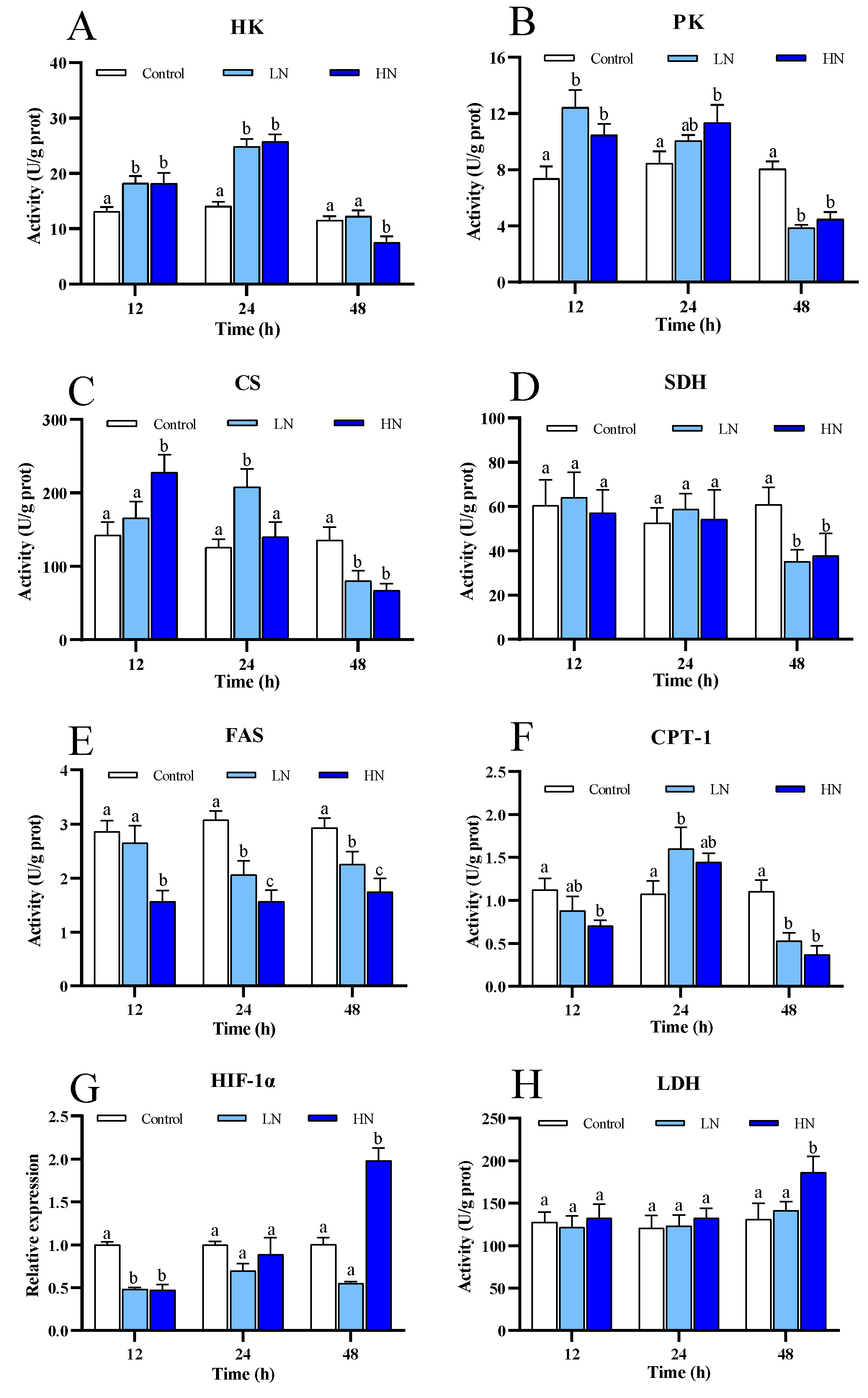
3.2. Energy Metabolism
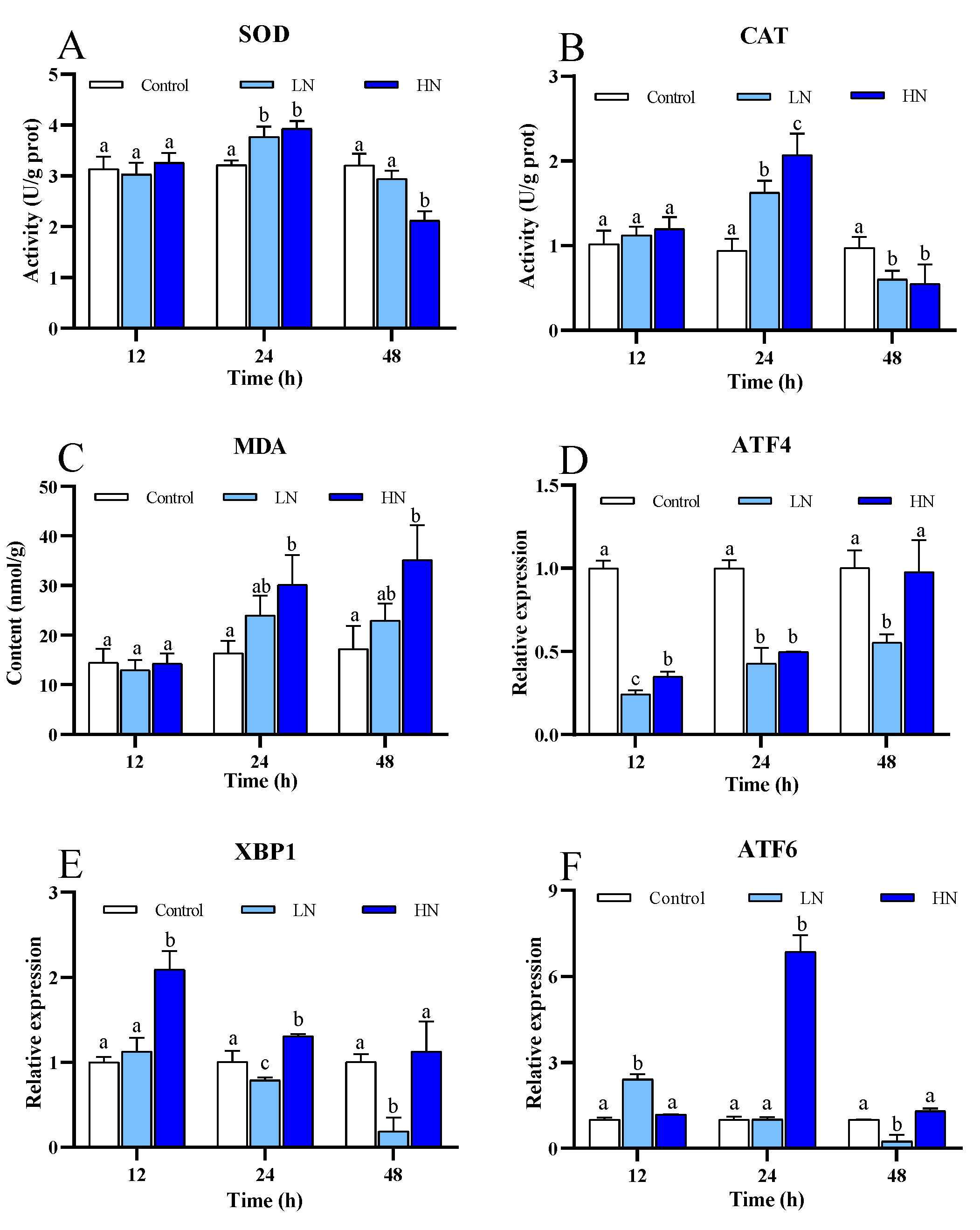
3.3. Antioxidant Defense and Unfolded Protein Response
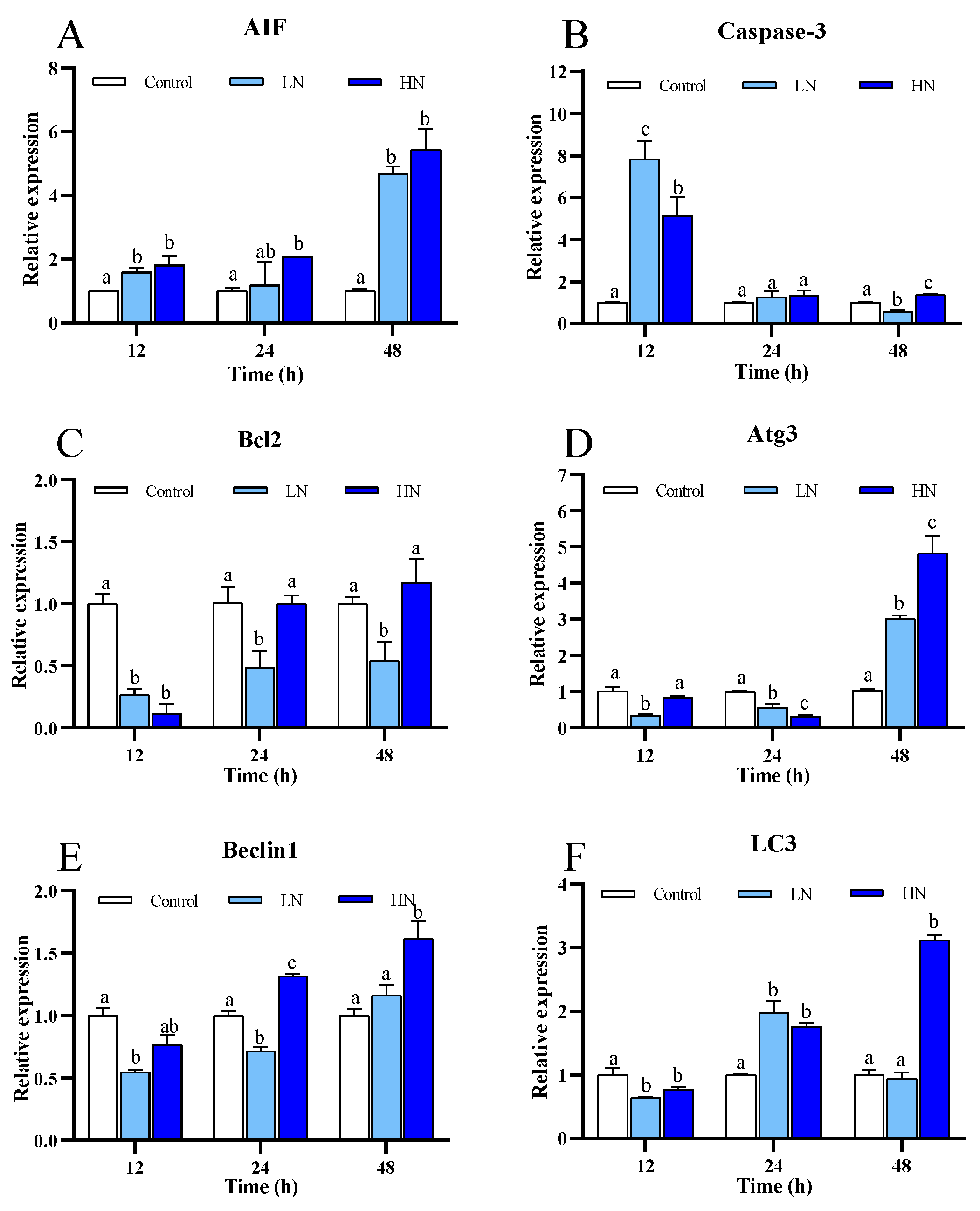
3.4. Apoptosis and Autophagy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fishery Department of Ministry of Agriculture and Rural Affairs of China. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2021. (In Chinese) [Google Scholar]
- Shi, C.; Zeng, T.; Li, R.; Wang, C.; Ye, Y.; Mu, C. Dynamic metabolite alterations of Portunus trituberculatus during larval development. J. Oceanol. Limnol. 2019, 37, 361–372. [Google Scholar] [CrossRef]
- Sun, P.; Jin, M.; Ding, L.; Lu, Y.; Ma, H.; Yuan, Y.; Zhou, Q. Dietary lipid levels could improve growth and intestinal microbiota of juvenile swimming crab, Portunus trituberculatus. Aquaculture 2018, 490, 208–216. [Google Scholar] [CrossRef]
- Liao, Y.Y.; Wang, H.H.; Lin, Z.G. Effect of ammonia and nitrite on vigour, survival rate, moulting rate of the blue swimming crab Portunus pelagicus zoea. Aquacult. Int. 2011, 19, 339–350. [Google Scholar] [CrossRef]
- Romano, N.; Zeng, C. Toxic Effects of Ammonia, Nitrite, and Nitrate to Decapod Crustaceans, A Review on Factors Influencing their Toxicity, Physiological Consequences, and Coping Mechanisms. Rev. Fish. Sci. 2013, 21, 1–21. [Google Scholar]
- Meng, X.; Jayasundara, N.; Zhang, J.; Ren, X.; Gao, B.; Li, J.; Liu, P. Integrated physiological, transcriptome and metabolome analyses of the hepatopancreas of the female swimming crab Portunus trituberculatus under ammonia exposure. Ecotoxicol. Environ. Saf. 2021, 228, 113026. [Google Scholar] [CrossRef]
- Kültz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef]
- Guo, H.; Xian, J.A.; Li, B.; Ye, C.X.; Wang, A.L.; Miao, Y.T.; Liao, S.A. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of white shrimp Litopenaeus vannamei under nitrite stress. Comp. Biochem. Physiol. C 2013, 157, 366–371. [Google Scholar] [CrossRef]
- Jiang, Q.; Zhang, W.; Tan, H.; Pan, D.; Yang, Y.; Ren, Q.; Yang, G. Analysis of gene expression changes, caused by exposure to nitrite, in metabolic and antioxidant enzymes in the red claw crayfish, Cherax quadricarinatus. Ecotox. Environ. Saf. 2014, 104, 423–428. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Jayasundara, N.; Ren, X.; Gao, B.; Liu, P.; Li, J.; Meng, X. Physiological and molecular responses in the gill of the swimming crab portunus trituberculatus during long-term ammonia stress. Front. Mar. Sci. 2021, 8, 1849. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Li, S.; Guo, X.; Fu, Y.; He, N.; Ruan, G.; Wang, Q.; Gao, W.; Liu, F. Impact of nitrite exposure on oxidative stress and antioxidative-related genes responses in the gills of Procambarus clarkii. Fish Shellfish Immunol. 2022, 131, 624–630. [Google Scholar] [CrossRef]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Chen, X.L.; Guo, Z.X. Nitrite-Induced Oxidative Stress, Histopathology, and Transcriptome Changes in the Mud Crab (Scylla paramamosain). Isr. J. Aquacult.-Bamid. IJA 2019, 71, 1626. [Google Scholar]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, Y.Q.; Guo, Z.X. Effect of nitrite exposure on oxidative stress, DNA damage and apoptosis in mud crab (Scylla paramamosain). Chemosphere 2019, 239, 124668. [Google Scholar] [CrossRef] [PubMed]
- Logue, S.E.; Cleary, P.; Saveljeva, S.; Samali, A. New directions in ER stress-induced cell death. Apoptosis 2013, 18, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, J.; Cao, J.; Liu, P.; Li, J.; Meng, X. Long-term Ammonia Toxicity in the Hepatopancreas of Swimming Crab Portunus Trituberculatus: Cellular Stress Response and Tissue Damage. Front. Mar. Sci. 2022, 8, 2105. [Google Scholar] [CrossRef]
- Ron, D. Translational control in the endoplasmic reticulum stress response. J. Clin. Investig. 2002, 110, 1383–1388. [Google Scholar] [CrossRef]
- Xiao, J.; Luo, S.S.; Du, J.H.; Liu, Q.Y.; Huang, Y.; Wang, W.F.; Wang, H.L. Transcriptomic analysis of gills in nitrite-tolerant and -sensitive families of Litopenaeus vannamei. Comp. Biochem. Physiol. C 2021, 253, 109212. [Google Scholar] [CrossRef]
- Fu, L.; Gao, T.; Jiang, H.; Zhang, Y.; Pan, J.L. Integrated miRNA-mRNA transcriptomic analysis of hepatopancreas reveals molecular strategies in Chinese mitten crab (Eriocheir sinensis) under acute nitrite stress. Aquacult. Int. 2021, 29, 1015–1030. [Google Scholar] [CrossRef]
- Li, Z.S.; Ma, S.; Shan, H.W.; Wang, T.; Xiao, W. Responses of hemocyanin and energy metabolism to acute nitrite stress in juveniles of the shrimp Litopenaeus vannamei. Ecotox. Environ. Saf. 2019, 186, 109753. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Xian, J.A.; Wang, A.L.; Hao, X.M.; Miao, Y.T.; Li, B.; Ye, C.X.; Liao, S.A. In vitro toxicity of nitrite on haemocytes of the tiger shrimp, Penaeus monodon, using flow cytometric analysis. Comp. Biochem. Physiol. C 2012, 156, 75–79. [Google Scholar] [CrossRef]
- Luzio, A.; Monteiro, S.M.; Fontaínhas-Fernandes, A.A.; Pinto-Carnide, O.; Matos, M.; Coimbra, A.M. Copper induced upregulation of apoptosis related genes in zebrafish (Danio rerio) gill. Aquat. Toxicol. 2013, 128–129, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ji, X.; Wang, X.; Li, T.; Wang, H.; Zeng, Q. Identification and characterization of differentially expressed genes in hepatopancreas of oriental river prawn Macrobrachium nipponense under nitrite stress. Fish. Shellfish Immunol. 2019, 87, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Luo, J.; Huang, Y.; Yuan, Y.; Cai, S. Effect of sub-lethal ammonia and nitrite stress on autophagy and apoptosis in hepatopancreas of Pacific whiteleg shrimp Litopenaeus vannamei. Fish. Shellfish Immunol. 2022, 130, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Shintani, T.; Klionsky, D.J. Autophagy in Health and Disease: A Double-Edged Sword. Science 2004, 306, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Dong, B.; Li, A.; Wu, L.; Zhang, Y.; Han, T.; Liu, X. scRNA-seq analysis reveals toxicity mechanisms in shrimp hemocytes subjected to nitrite stress. Chemosphere 2023, 316, 137853. [Google Scholar] [CrossRef]
- Chen, J.C.; Chin, T.S. Acute Toxicity of Nitrite to Tiger Prawn, Penaeus monodon, Larvae. Aquaculture 1988, 69, 253–262. [Google Scholar] [CrossRef]
- Chand, R.K.; Sahoo, P.K. Effect of nitrite on the immune response of freshwater prawn Macrobrachium malcolmsonii and its susceptibility to Aeromonas hydrophila. Aquaculture 2006, 258, 150–156. [Google Scholar] [CrossRef]
- Guo, H.; Xian, J.A.; Wang, A.L. Analysis of digital gene expression profiling in hemocytes of white shrimp Litopenaeus vannamei under nitrite stress. Fish. Shellfish Immunol. 2016, 56, 1–11. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, D.; Wang, F.; Dong, S. Hypothermal effects on survival, energy homeostasis and expression of energy-related genes of swimming crabs Portunus trituberculatus during air exposure. J. Therm. Biol. 2016, 60, 33–40. [Google Scholar] [CrossRef]
- Tian, W.X.; Hsu, R.Y.; Wang, Y.S. Studies on the reactivity of the essential sulfhydryl groups as a conformational probe for the fatty acid synthetase of chicken liver. Inactivation by 5,5’-dithiobis-(2-nitrobenzoic acid) and intersubunit cross-linking of the inactivated enzyme. J. Biol. Chem. 1985, 260, 11375–11387. [Google Scholar] [CrossRef] [PubMed]
- Bieber, L.L.; Fiol, C. Purification and assay of carnitine acyltransferases. Method. Enzynol. 1986, 123, 276–284. [Google Scholar]
- Mallasen, M.; Valenti, W.C. Effect of nitrite on larval development of giant river prawn Macrobrachium rosenbergii. Aquaculture 2006, 261, 1292–1298. [Google Scholar] [CrossRef]
- Lv, G.T.; Wang, Z.Z.; Wang, H.P.; Yuan, J.Y. Acute toxic efects of NaNO2 on Portunus trituberculatus larvas. J. Zhejiang Ocean Univ. (Nat. Sci. Ed.) 2006, 25, 244–248. [Google Scholar]
- Gomes, R.S.; De Lima, J.P.V.; Cavalli, O.; Correia, E.S. Acute Toxicity of Ammonia and Nitrite to Painted River Prawn, Macrobrachium carcinus, Larvae. J. World Aquacult. Soc. 2016, 47, 239–247. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Alexey, A.; Sukhotin, D. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Hong, M.; Chen, L.; Qin, J.G.; Sun, X.; Li, E.; Gu, S.; Yu, N. Acute tolerance and metabolic responses of Chinese mitten crab (Eriocheir sinensis) juveniles to ambient nitrite. Comp. Biochem. Physiol. C 2009, 149, 419–426. [Google Scholar] [CrossRef]
- Hardie, D.G.; Scott, J.W.; Pan, D.A.; Hudson, E.R. Management of cellular energy by the AMP-activated protein kinase system. FEBS Lett. 2003, 546, 113–120. [Google Scholar] [CrossRef]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef]
- Xu, Z.; Li, E.; Xu, C.; Gan, L.; Qin, J.G. Response of AMP-activated protein kinase and energy metabolism to acute nitrite exposure in the Nile tilapia Oreochromis niloticus. Aquat. Toxicol. 2016, 177, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Frederich, M.; O’Rourke, M.R.; Furey, N.B.; Jost, J.A. AMP-activated protein kinase (AMPK) in the rock crab, Cancer irroratus: An early indicator of temperature stress. J. Exp. Biol. 2009, 212, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Chen, J.C. Hemocyanin oxygen affinity, and the fractionation of oxyhemocyanin and deoxyhemocyanin for Penaeus monodon exposed to elevated nitrite. Aquat. Toxicol. 1999, 45, 35–46. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, Q.Y.; Du, J.H.; Zhu, W.L.; Wang, H.L. Integrated analysis of physiological, transcriptomic and metabolomic responses and tolerance mechanism of nitrite exposure in Litopenaeus vannamei. Sci. Total. Environ. 2019, 711, 134416. [Google Scholar] [CrossRef] [PubMed]
- Borgese, N.; Francolini, M.; Snapp, E. Endoplasmic reticulum architecture: Structures in flux. Curr. Opin. Cell Biol. 2006, 18, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Su, S.; Song, C.; Yu, F.; Zhou, J.; Li, J.; Jia, R.; Xu, P.; Tang, Y. Effects of Copper Exposure on Oxidative Stress, Apoptosis, Endoplasmic Reticulum Stress, Autophagy and Immune Response in Different Tissues of Chinese Mitten Crab (Eriocheir sinensis). Antioxidants 2022, 11, 2029. [Google Scholar] [CrossRef]
- Liu, H.J.; Dong, M.; Jiang, W.D.; Wu, P.; Liu, Y.; Jin, X.W.; Kuang, S.Y.; Tang, L.; Zhang, L.; Feng, L.; et al. Acute nitrite exposure-induced oxidative damage, endoplasmic reticulum stress, autophagy and apoptosis caused gill tissue damage of grass carp (Ctenopharyngodon idella): Relieved by dietary protein. Ecotoxicol. Environ. Saf. 2022, 243, 113994. [Google Scholar] [CrossRef]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Sign. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Candé, C.; Cohen, I.; Daugas, E.; Ravagnan, L.; Larochette, N.; Zamzami, N.; Kroemer, G. Apoptosis-inducing factor (AIF): A novel caspase-independent death effector released from mitochondria. Biochimie 2002, 84, 215–222. [Google Scholar] [CrossRef]
- Denton, D.; Nicolson, S.; Kumar, S. Cell death by autophagy: Facts and apparent artefacts. Cell Death. Differ. 2012, 19, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Zeh, H.J.; Lotze, M.T.; Tang, D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death. Differ. 2011, 18, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Menon, M.B.; Dhamija, S. Beclin 1 Phosphorylation—At the Center of Autophagy Regulation. Front. Cell. Dev. Biol. 2018, 6, 137. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Primer | Reserve Primer |
|---|---|---|
| AMPKα | GCACATTCTCACCGGCACTA | CGGAAGAGCTTGAGGTTGGT |
| AMPKβ1 | TTTGTGGCAATCGTGGACCT | ACGTTGTTCTGAGTGCCCAT |
| AMPKβ2 | GTAACCACAGCTCGCAGGAT | GCTTGGTGATGCTGGTGTTG |
| AMPKγ | TCAGCCCCTCAGATTCAAGC | CACTTTGCACGTTTCCCTGG |
| HIF-1α | CCAGCTGATGTGTAGGAGCA | GAGCTGGTGAATAGGGTGGA |
| ATF4 | AAGGCACCATCTTCCAACTG | TGCTGGAAGTGGACAGACAG |
| ATF6 | TACCACAGCTGACACACGC | GCAGCAGGTTCGTCTCCAT |
| XBP1 | GTGATGGACTCTGCACTGC | GGGTTCCAGGACTGTTGCT |
| Beclin1 | GGTGAGGAGGTGGAGTTTGA | GGGGCTGAGTCTTCTCTGTG |
| LC3 | CGGACGGGATCATGAAGTTA | ACACCACCACCGGAATCTTA |
| Atg3 | TGGTAGAGGACCCCACAGAC | TCTTGTCCACCACGTTCTTG |
| AIF | GAATGAACTGCCAAAACCAC | AGTCCAAAATCTACCAATCCAC |
| Caspase-3 | TGGCAGTGGTGGCTTGTCT | CGTGGCTTGTTGAGCAGTG |
| Bcl-2 | AGCTTACAACTGGATGCGCT | TCGAGAGTGATTTAGGCGGC |
| β-actin | CGAAACCTTCAACACTCCCG | GGGACAGTGTGTGAAACGCC |
| Time | Group | ATP | ADP | AMP | TAN | ADP/ATP | AMP/ATP | AEC |
|---|---|---|---|---|---|---|---|---|
| 12 h | Control | 3.38 a | 1.38 a | 0.44 a | 5.20 a | 0.41 a | 0.13 a | 0.78 a |
| LN | 4.02 b | 1.36 a | 0.43 a | 5.81 b | 0.34 a | 0.11 a | 0.80 a | |
| HN | 4.34 b | 1.47 a | 0.49 a | 6.30 b | 0.34 a | 0.11 a | 0.81 a | |
| 24 h | Control | 3.35 a | 1.45 a | 0.50 a | 5.30 a | 0.44 a | 0.15 a | 0.77 a |
| LN | 2.49 b | 1.89 a | 0.65 a | 5.04 a | 0.76 b | 0.26 b | 0.68 b | |
| HN | 2.09 b | 1.64 a | 0.51 a | 4.24 b | 0.80 b | 0.25 b | 0.69 b | |
| 48 h | Control | 3.45 a | 1.47 a | 0.52 a | 5.43 a | 0.43 a | 0.15 a | 0.77 a |
| LN | 2.05 b | 1.95 a | 0.52 a | 4.52 b | 0.95 b | 0.25 b | 0.67 b | |
| HN | 2.06 b | 1.38 a | 0.44 a | 3.88 b | 0.70 ab | 0.22 b | 0.71 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wang, D.; Shang, Y.; Yu, X.; Gao, B.; Lv, J.; Li, J.; Liu, P.; Li, J.; Meng, X. Survival, Energy Status, and Cellular Stress Responses of the Juvenile Swimming Crab Portunus trituberculatus under Acute Nitrite Stress. Fishes 2023, 8, 215. https://doi.org/10.3390/fishes8040215
Liu X, Wang D, Shang Y, Yu X, Gao B, Lv J, Li J, Liu P, Li J, Meng X. Survival, Energy Status, and Cellular Stress Responses of the Juvenile Swimming Crab Portunus trituberculatus under Acute Nitrite Stress. Fishes. 2023; 8(4):215. https://doi.org/10.3390/fishes8040215
Chicago/Turabian StyleLiu, Xiaochen, Daixia Wang, Yan Shang, Xuee Yu, Baoquan Gao, Jianjian Lv, Jitao Li, Ping Liu, Jian Li, and Xianliang Meng. 2023. "Survival, Energy Status, and Cellular Stress Responses of the Juvenile Swimming Crab Portunus trituberculatus under Acute Nitrite Stress" Fishes 8, no. 4: 215. https://doi.org/10.3390/fishes8040215
APA StyleLiu, X., Wang, D., Shang, Y., Yu, X., Gao, B., Lv, J., Li, J., Liu, P., Li, J., & Meng, X. (2023). Survival, Energy Status, and Cellular Stress Responses of the Juvenile Swimming Crab Portunus trituberculatus under Acute Nitrite Stress. Fishes, 8(4), 215. https://doi.org/10.3390/fishes8040215