
Environmental Impacts on Skin Microbiomes of Sympatric High Arctic Salmonids
 ,
,
Abstract
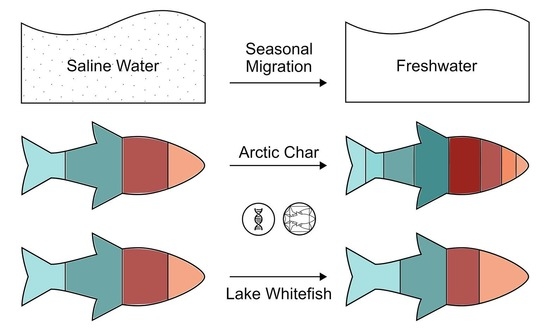
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Area, Fish, and Water Sampling
2.2. Fish Condition and Growth Curve Calculations
2.3. DNA Extractions and Sequencing
2.4. Statistical Analyses, Data Availability, and Efforts to Reduce Environmental Impact
3. Results
3.1. Condition Factors and Annual Incremental Growth
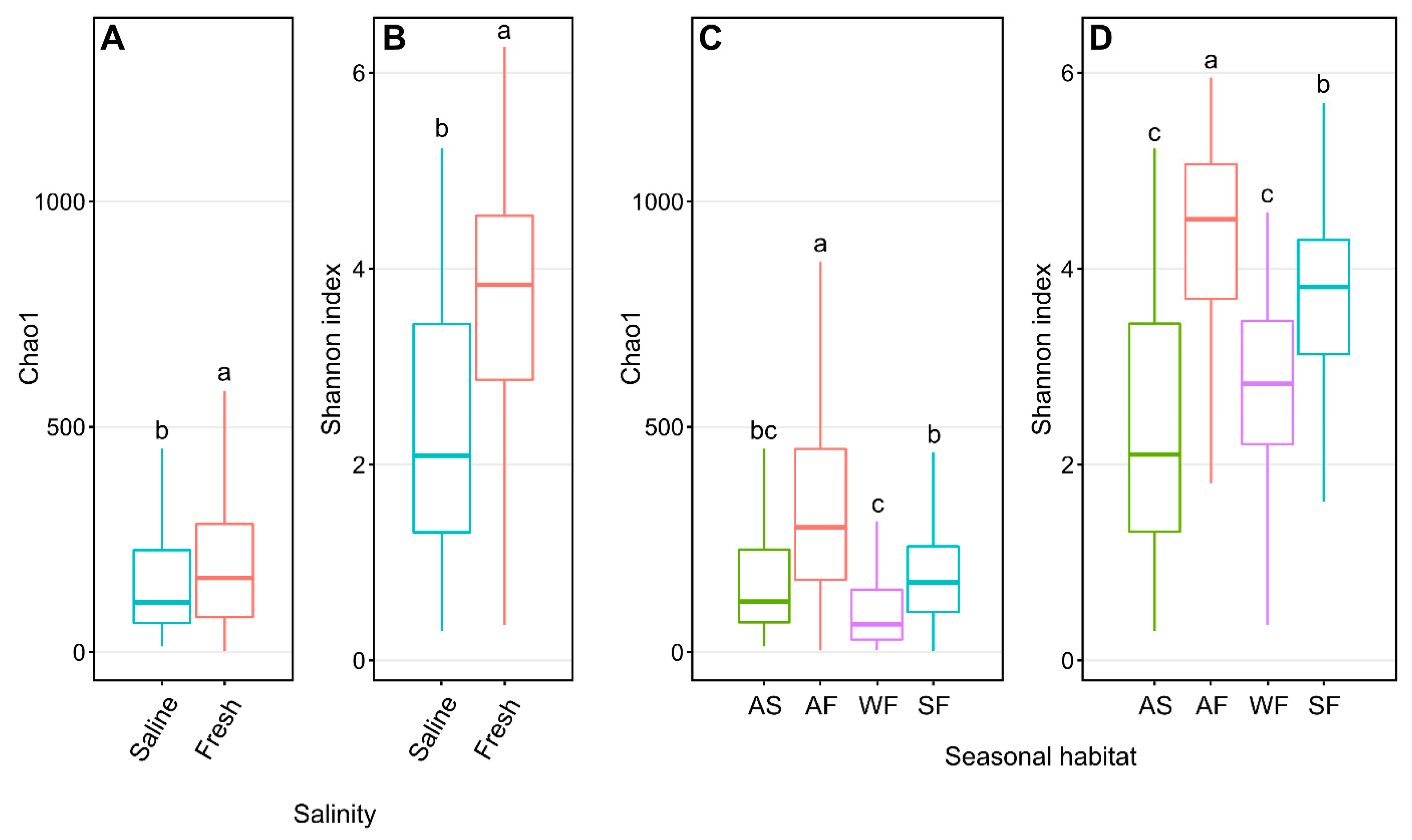
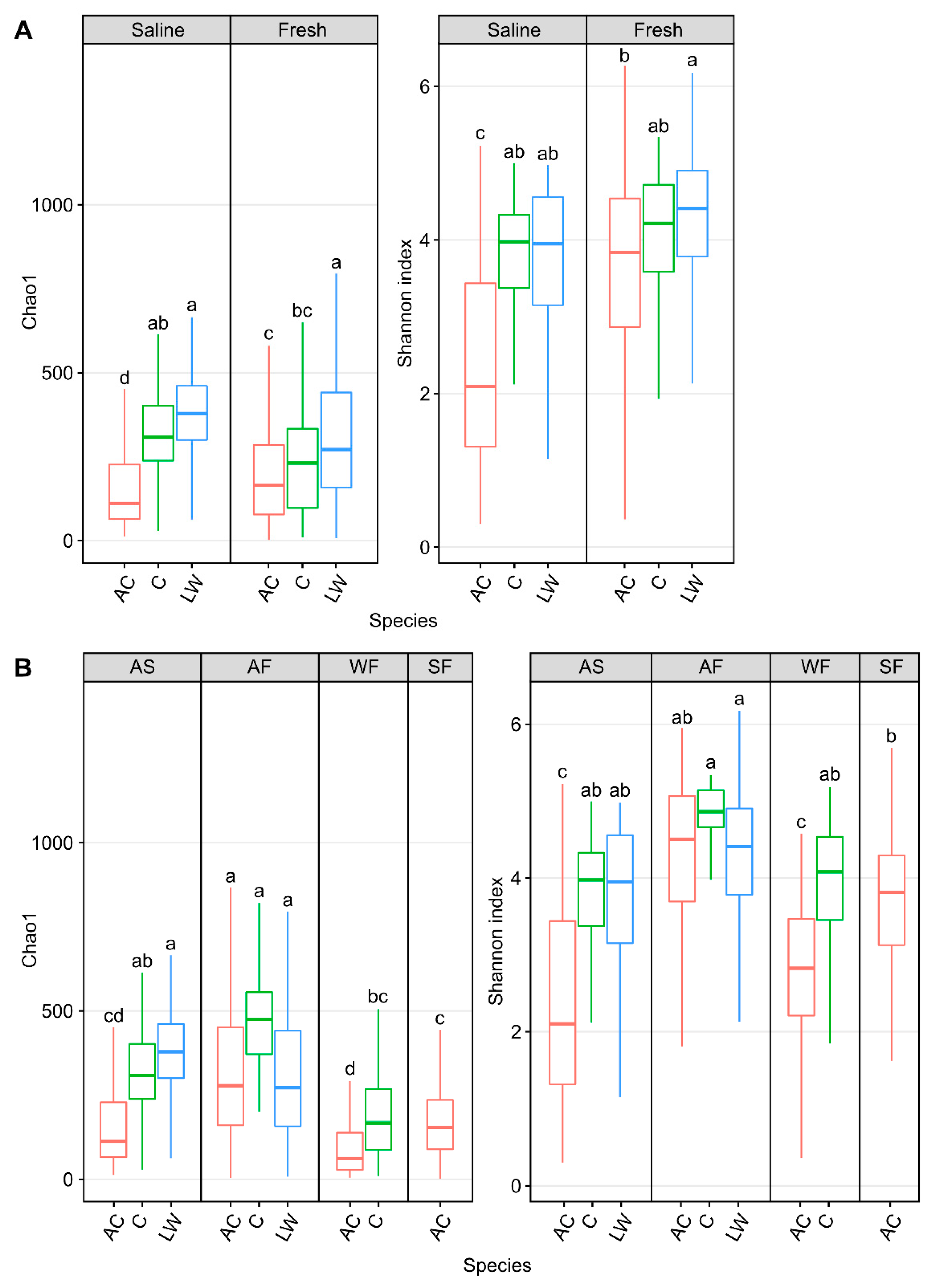
3.2. Arctic Char Skin Microbiome
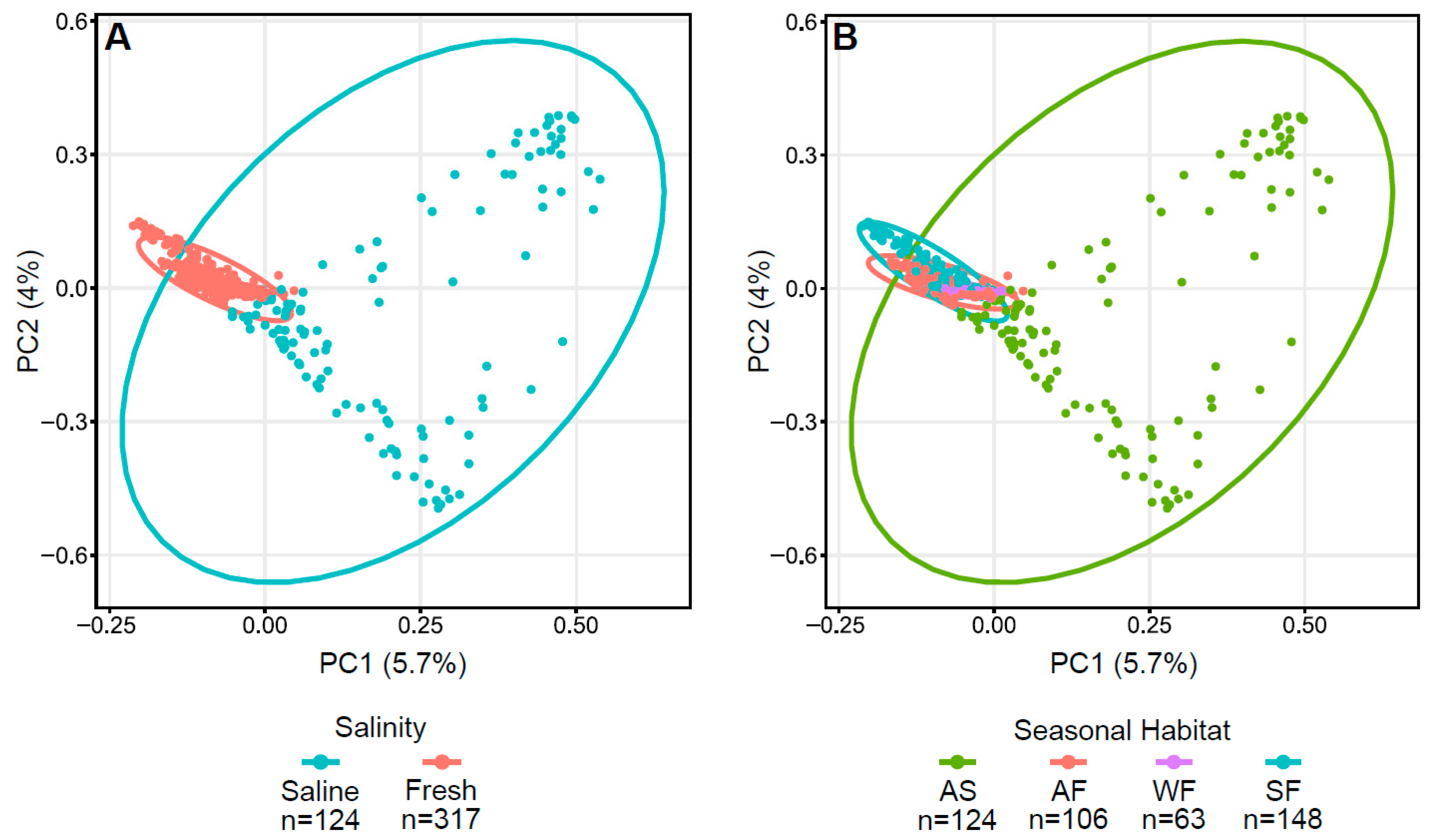
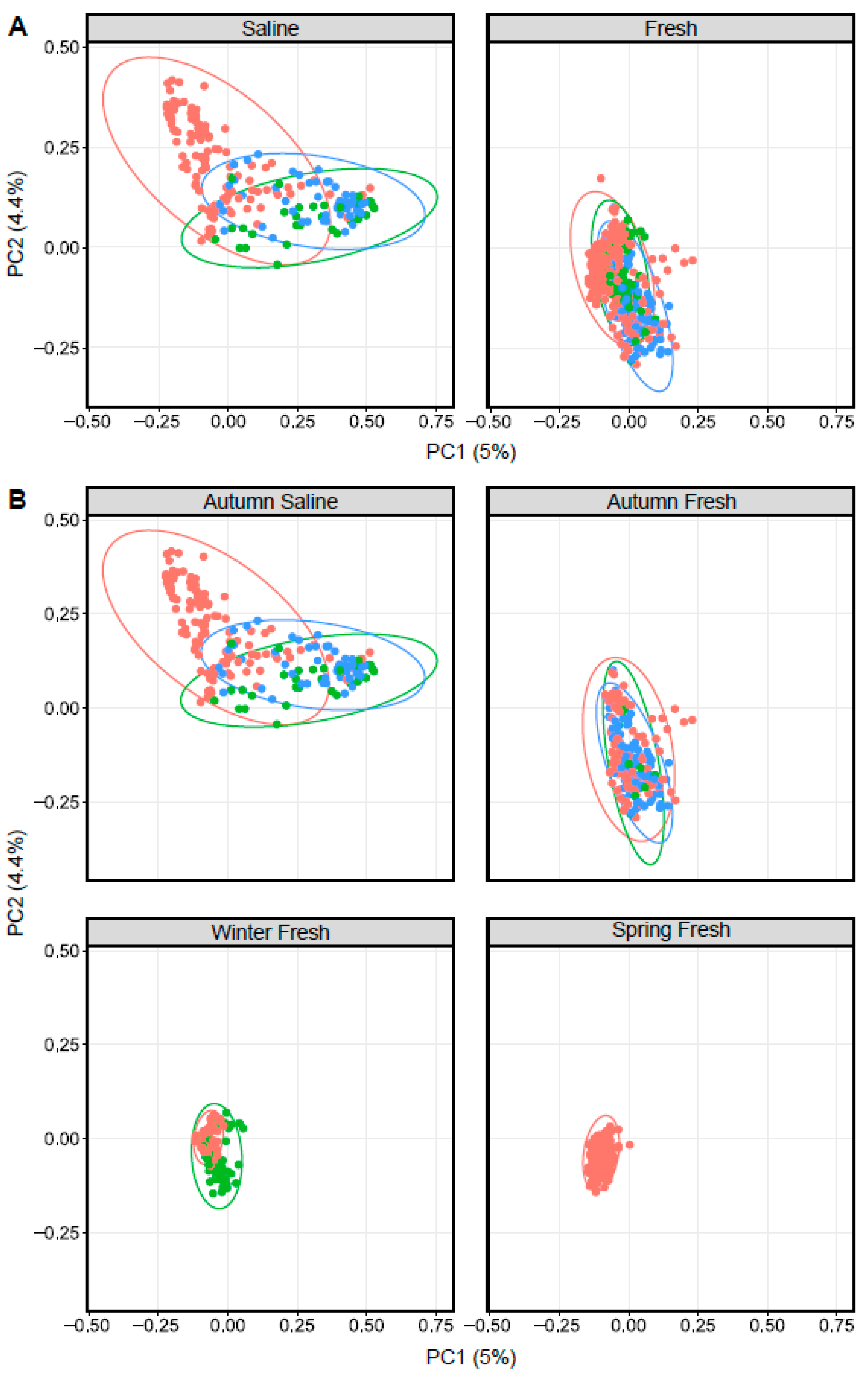
3.3. Influence of Surrounding Water on Arctic Char Skin Communities
3.4. CSC Skin Microbiomes, Fishing Sites, and Water Microbiota
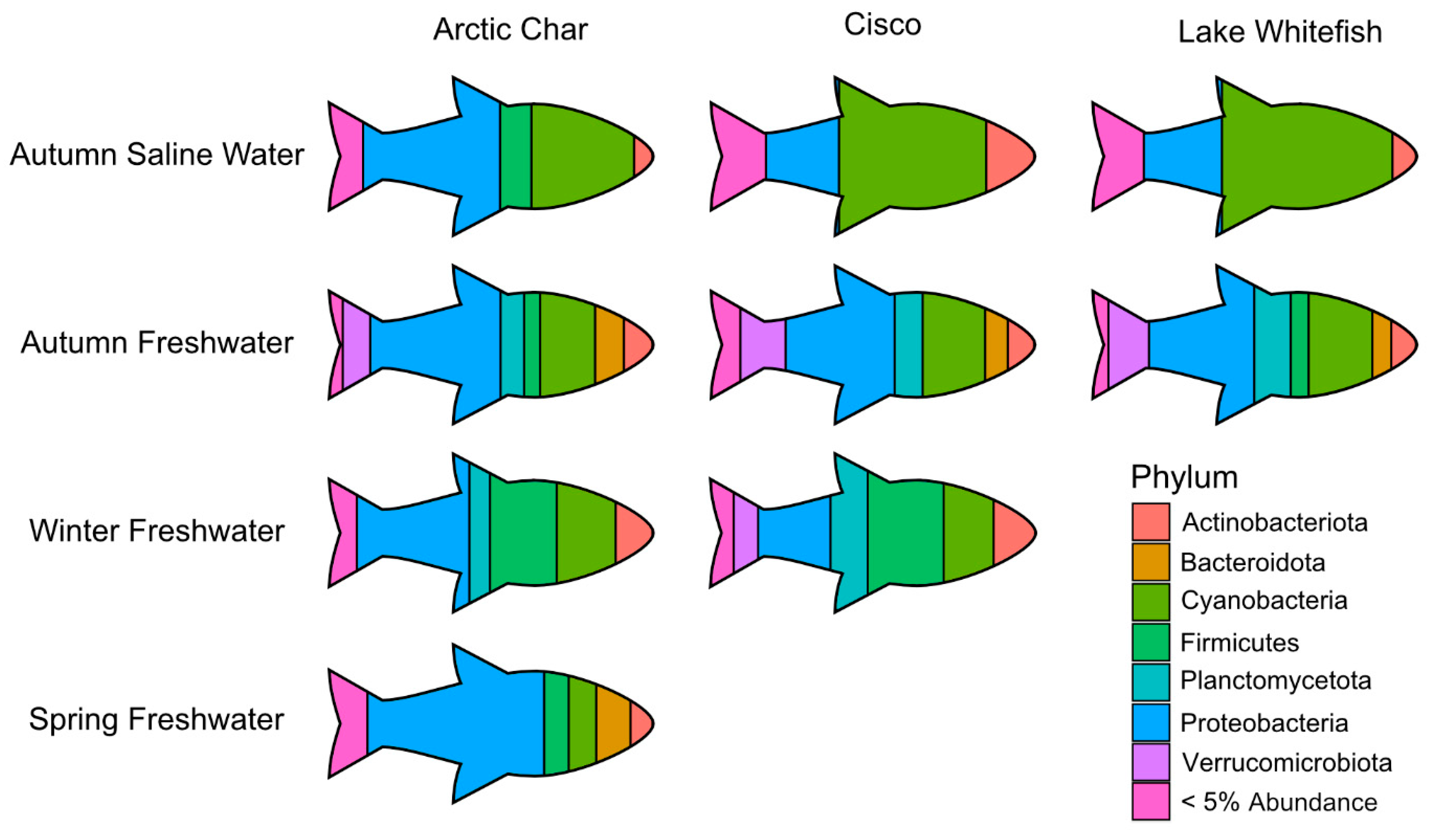
3.5. Impact of Fish Host on Skin Microbiomes
4. Discussion
4.1. Skin Microbiota in Related Migrating Salmonids
4.2. Adaptation to Environmental Conditions, a Changing Climate, and Fisheries Management
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klemetsen, A.; Amundsen, P.A.; Dempson, J.B.; Jonsson, B.; Jonsson, N.; O’Connell, M.F.; Mortensen, E. Atlantic Salmon Salmo salar L., Brown Trout Salmo trutta L. and Arctic Charr Salvelinus alpinus (L.): A Review of Aspects of Their Life Histories. Ecol. Freshw. Fish 2003, 12, 1–59. [Google Scholar] [CrossRef]
- Scott, W.B.; Crossman, E.J. Freshwater Fishes of Canada. Fish. Res. Board Can. Bull. 1973, 184, 966. [Google Scholar]
- Morin, R.; Dodson, J.J.; Power, G. Life History Variations of Anadromous Cisco (Coregonus artedii), Lake Whitefish (C. clupeaformis), and Round Whitefish (Prosopium cylindraceum) Populations of Eastern James–Hudson Bay. Can. J. Fish. Aquat. Sci. 1982, 39, 958–967. [Google Scholar] [CrossRef]
- Brown, R.J.; Bickford, N.; Severin, K. Otolith Trace Element Chemistry as an Indicator of Anadromy in Yukon River Drainage Coregonine Fishes. Trans. Am. Fish. Soc. 2007, 136, 678–690. [Google Scholar] [CrossRef]
- Koch, I.; Das, P.; McPhedran, B.E.; Casselman, J.M.; Moniz, K.L.; van Coeverden de Groot, P.; Chen, C.Y.; Walker, V.K. Correlation of Mercury Occurrence with Age, Elemental Composition, and Life History in Sea-Run Food Fish from the Canadian Arctic Archipelago’s Lower Northwest Passage. Foods 2021, 10, 2621. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, G.; Li, S.; Li, X.; Liu, Y. Intestinal Microbiota of Healthy and Unhealthy Atlantic Salmon Salmo salar L. in a Recirculating Aquaculture System. J. Ocean Limnol. 2018, 36, 414–426. [Google Scholar] [CrossRef]
- Webster, T.M.U.; Consuegra, S.; Hitchings, M.; de Leaniz, C.G. Interpopulation Variation in the Atlantic Salmon Microbiome Reflects Environmental and Genetic Diversity. Appl. Environ. Microbiol. 2018, 84, e00691-18. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, F.É.; Holland, A.; Bouslama, S.; Audet-Gilbert, É.; Lavoie, C.; Val, A.L.; Derome, N. Fish Skin and Gut Microbiomes Show Contrasting Signatures of Host Species and Habitat. Appl. Environ. Microbiol. 2020, 86, e00789-20. [Google Scholar] [CrossRef]
- Lokesh, J.; Kiron, V. Transition from Freshwater to Seawater Reshapes the Skin-Associated Microbiota of Atlantic Salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A. Seawater Transfer Alters the Intestinal Microbiota Profiles of Atlantic Salmon (Salmo salar L.). Sci. Rep. 2017, 7, 13877. [Google Scholar] [CrossRef]
- Hamilton, E.F.; Element, G.; van Coeverden de Groot, P.; Engel, K.; Neufeld, J.D.; Shah, V.; Walker, V.K. Anadromous Arctic Char Microbiomes: Bioprospecting in the High Arctic. Front. Bioeng. Biotechnol. 2019, 7, 32. [Google Scholar] [CrossRef]
- Wilson, B.; Danilowicz, B.S.; Meijer, W.G. The Diversity of Bacterial Communities Associated with Atlantic Cod Gadus morhua. Microb. Ecol. 2008, 55, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Kueneman, J.G.; Parfrey, L.W.; Woodhams, D.C.; Archer, H.M.; Knight, R.; McKenzie, V.J. The Amphibian Skin-Associated Microbiome across Species, Space and Life History Stages. Mol. Ecol. 2014, 23, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A. Marine Animal Microbiomes: Toward Understanding Host–Microbiome Interactions in a Changing Ocean. Front. Mar. Sci. 2017, 4, 222. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Wong, M.K.-S.; Hyodo, S.; Goto, S.; Hamasaki, K. Temperature Modulation Alters the Gut and Skin Microbial Profiles of Chum Salmon (Oncorhynchus keta). Front. Mar. Sci. 2022, 9, 1027621. [Google Scholar] [CrossRef]
- Xu, Z.; Parra, D.; Gomez, D.; Salinas, I.; Zhang, Y.A.; Von Gersdorff Jorgensen, L.; LaPatra, S.E.; Sunyer, J.O. Teleost Skin, an Ancient Mucosal Surface That Elicits Gut-Like Immune Responses. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef]
- Esteban, M.Á.; Cerezuela, R. Fish Mucosal Immunity: Skin. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 67–92. [Google Scholar] [CrossRef]
- Scharschmidt, T.C.; Fischbach, M.A. What Lives on Our Skin: Ecology, Genomics and Therapeutic Opportunities of the Skin Microbiome. Drug Discov. Today Dis. Mech. 2013, 10, e83–e89. [Google Scholar] [CrossRef] [PubMed]
- Klenerman, P.; Ogg, G. Killer T Cells Show Their Kinder Side. Nature 2018, 555, 594–595. [Google Scholar] [CrossRef]
- Linehan, J.L.; Harrison, O.J.; Han, S.J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.; Tamoutounour, S.; et al. Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair. Cell 2018, 172, 784–796. [Google Scholar] [CrossRef]
- Sanford, J.A.; Zhang, L.J.; Williams, M.R.; Gangoiti, J.A.; Huang, C.M.; Gallo, R.L. Inhibition of HDAC8 and HDAC9 by Microbial Short-Chain Fatty Acids Breaks Immune Tolerance of the Epidermis to TLR Ligands. Sci. Immunol. 2016, 1, eaah4609. [Google Scholar] [CrossRef]
- Chiarello, M.; Paz-Vinas, I.; Veyssière, C.; Santoul, F.; Loot, G.; Ferriol, J.; Boulêtreau, S. Environmental Conditions and Neutral Processes Shape the Skin Microbiome of European Catfish (Silurus glanis) Populations of Southwestern France. Environ. Microbiol. Rep. 2019, 11, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.M.; Tao, Z.; Bullard, S.A.; Arias, C.R. Diversity of the Skin Microbiota of Fishes: Evidence for Host Species Specificity. FEMS Microbiol. Ecol. 2013, 85, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Boutin, S.; Sauvage, C.; Bernatchez, L.; Audet, C.; Derome, N. Interindividual Variations of the Fish Skin Microbiota: Host Genetics Basis of Mutualism? PLoS ONE 2014, 9, e102649. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.A.; Müller, K.M.; Weese, J.S.; Neufeld, J.D. Comprehensive Skin Microbiome Analysis Reveals the Uniqueness of Human Skin and Evidence for Phylosymbiosis within the Class Mammalia. Proc. Natl. Acad. Sci. USA 2018, 115, E5786–E5795. [Google Scholar] [CrossRef]
- Brooks, A.W.; Kohl, K.D.; Brucker, R.M.; van Opstal, E.J.; Bordenstein, S.R. Phylosymbiosis: Relationships and Functional Effects of Microbial Communities across Host Evolutionary History. PLoS Biol. 2016, 14, e2000225. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.; Salinas, I. Under Pressure: Interactions between Commensal Microbiota and the Teleost Immune System. Front. Immunol. 2017, 8, 559. [Google Scholar] [CrossRef] [PubMed]
- Crête-Lafrenière, A.; Weir, L.K.; Bernatchez, L. Framing the Salmonidae family phylogenetic portrait: A more complete picture from increased taxon sampling. PLoS ONE 2012, 7, e46662. [Google Scholar] [CrossRef]
- Wu, Y.; Lougheed, D.R.; Lougheed, S.C.; Moniz, K.; Walker, V.K.; Colautti, R.I. baRcodeR: An Open-Source R Package for Sample Labelling. Methods Ecol. Evol. 2020, 11, 980–985. [Google Scholar] [CrossRef]
- Campana, S.E.; Casselman, J.M.; Jones, C.M. Bomb Radiocarbon Chronologies in the Arctic, with Implications for the Age Validation of Lake Trout (Salvelinus namaycush) and Other Arctic Species. Can. J. Fish. Aquat. Sci. 2008, 65, 733–743. [Google Scholar] [CrossRef]
- Casselman, J.M.; Jones, C.M.; Campana, S.E. Bomb Radiocarbon Age Validation for the Long-Lived, Unexploited Arctic Fish Species Coregonus clupeaformis. Mar. Freshw. Res. 2019, 70, 1781–1788. [Google Scholar] [CrossRef]
- Barnham, C.; Baxter, A. Condition Factor, K, for Salmonid Fish. Fisheries Notes. Nat. Resour. Environ. 1998, 5, 1–3. [Google Scholar]
- Isermann, D.A.; Knight, C.T. A Computer Program for Age-Length Keys Incorporating Age Assignment to Individual Fish. N. Am. J. Fish. Manag. 2005, 25, 1153–1160. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit rRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Quince, C.; Lanzen, A.; Davenport, R.J.; Turnbaugh, P.J. Removing Noise from Pyrosequenced Amplicons. BMC Bioinform. 2011, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Greer, C.W. Chemical Dispersants Enhance the Activity of Oil-and Gas Condensate-Degrading Marine Bacteria. ISME J. 2017, 11, 2793–2808. [Google Scholar] [CrossRef] [PubMed]
- Cobanli, S.E.; Wohlgeschaffen, G.; Ryther, C.; MacDonald, J.; Gladwell, A.; Watts, T.; Greer, C.W.; Elias, M.; Wasserscheid, J.; Robinson, B.; et al. Microbial Community Response to Simulated Diluted Bitumen Spills in Coastal Seawater and Implications for Oil Spill Response. FEMS Microbiol. Ecol. 2022, 98, fiac033. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Min, D.; Doxey, A.C.; Neufeld, J.D. AXIOME3: Automation, Extension, and Integration of Microbial Ecology. GigaScience 2021, 10, giab006. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.F.; Juurakko, C.L.; Engel, K.; de Groot, P.J.C.; Casselman, J.M.; Greer, C.W.; Neufeld, J.D.; Walker, V.K. Characterization of Skin- and Intestine Microbial Communities in Migrating High Arctic Lake Whitefish and Cisco. bioRxiv. 2023. Available online: https://www.biorxiv.org/content/10.1101/2023.03.08.531621v1 (accessed on 12 April 2023). [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersion. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Arbizu, P.M. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis, R Package Version 0.4. 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 12 April 2023).
- Wickham, H. Elegant Graphics for Data Analysis (ggplot2); Springer: New York, NY, USA, 2009. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Lahti, L.; Shetty, S. Microbiome R Package. Bioconductor 2017. Available online: https://bioconductor.org/packages/release/bioc/html/microbiome.html (accessed on 12 April 2023). [CrossRef]
- Liu, S.; Wawrik, B.; Liu, Z. Different Bacterial Communities Involved in Peptide Decomposition between Normoxic and Hypoxic Coastal Waters. Front. Microbiol. 2017, 8, 353. [Google Scholar] [CrossRef]
- Jiang, X.; Zhu, Z.; Wu, J.; Lian, E.; Liu, D.; Yang, S.; Zhang, R. Bacterial and Protistan Community Variation across the Changjiang Estuary to the Ocean with Multiple Environmental Gradients. Microorganisms 2022, 10, 991. [Google Scholar] [CrossRef]
- Padra, J.T.; Murugan, A.V.; Sundell, K.; Sundh, H.; Benktander, J.; Lindén, S.K. Fish Pathogen Binding to Mucins from Atlantic Salmon and Arctic Char Differs in Avidity and Specificity and Is Modulated by Fluid Velocity. PLoS ONE 2019, 14, e0215583. [Google Scholar] [CrossRef]
- Wu, Z.; Kan, F.W.K.; She, Y.M.; Walker, V.K. Biofilm, Ice Recrystallization Inhibition and Freeze-Thaw Protection in an Epiphyte Community. Appl. Biochem. Microbiol. 2012, 48, 363–370. [Google Scholar] [CrossRef]
- Cai, W.; De La Fuente, L.; Arias, C.R. Biofilm Formation by the Fish Pathogen Flavobacterium columnare: Development and Parameters Affecting Surface Attachment. Appl. Environ. Microbiol. 2013, 79, 5633–5642. [Google Scholar] [CrossRef]
- Christmas, N.A.; Barker, G.; Anesio, A.M.; Sánchez-Baracaldo, P. Genomic Mechanisms for Cold Tolerance and Production of Exopolysaccharides in the Arctic Cyanobacterium Phormidesmis Priestleyi BC1401. BMC Genom. 2016, 17, 533. [Google Scholar] [CrossRef]
- Webster, T.M.U.; Rodriguez-Barreto, D.; Castaldo, G.; Gough, P.; Consuegra, S.; de Leaniz, C.G. Environmental Plasticity and Colonisation History in the Atlantic Salmon Microbiome: A Translocation Experiment. Mol. Ecol. 2020, 29, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, P.; De Benedetto, G.E.; Francavilla, M.; Barca, A.; Caroppo, C. Bioactive Potential of Two Marine Picocyanobacteria Belonging to Cyanobium and Synechococcus Genera. Microorganisms 2021, 9, 2048. [Google Scholar] [CrossRef]
- Boutin, S.; Bernatchez, L.; Audet, C.; Derôme, N. Network Analysis Highlights Complex Interactions between Pathogen, Host and Commensal Microbiota. PLoS ONE 2013, 8, e84772. [Google Scholar] [CrossRef] [PubMed]
- Chiarello, M.; Villeger, S.; Bouvier, C.; Bettarel, Y.; Bouvier, T. High Diversity of Skin-Associated Bacterial Communities of Marine Fishes Is Promoted by Their High Variability among Body Parts, Individuals and Species. FEMS Microbiol. Ecol. 2015, 91, fiv061. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef]
- Øygarden, E.T. Influence of Genetic Background and Environmental Factors on the Skin and Gut Microbiota of Atlantic Salmon (Salmo salar) Fry. Master’s Thesis, Norwegian University of Science and Technology, Trondheim, Norway, 2017. Available online: https://ntnuopen.ntnu.no/ntnu-xmlui/handle/11250/2454377 (accessed on 1 May 2021).
- Guardabassi, L.; Dalsgaard, A.; Olsen, J.E. Phenotypic Characterization and Antibiotic Resistance of Acinetobacter spp. Isolated from Aquatic Sources. J. Appl. Microbiol. 1999, 87, 659–667. [Google Scholar] [CrossRef]
- Junge, K.; Christner, B.; Staley, J. Diversity of Psychrophilic Bacteria from Sea Ice and Glacial Ice Communities. In Extremophiles Handbook; Horikoshi, K., Ed.; Springer: Tokyo, Japan, 2011; pp. 794–815. [Google Scholar] [CrossRef]
- Zhan, M.; Huang, Z.; Cheng, G.; Yu, Y.; Su, J.; Xu, Z. Alterations of the Mucosal Immune Response and Microbial Community of the Skin upon Viral Infection in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2022, 23, 14037. [Google Scholar] [CrossRef]
- Ernst, B.; Hoeger, S.J.; O’Brien, E.; Dietrich, D.R. Oral Toxicity of the Microcystin-Containing Cyanobacterium Planktothrix rubescens in European Whitefish (Coregonus lavaretus). Aquat. Toxicol. 2006, 79, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Faria, S.I.; Teixeira-Santos, R.; Romeu, M.J.; Morais, J.; Jong, E.D.; Sjollema, J.; Vasconcelos, V.; Mergulhão, F.J. Unveiling the Antifouling Performance of Different Marine Surfaces and Their Effect on the Development and Structure of Cyanobacterial Biofilms. Microorganisms 2021, 9, 1102. [Google Scholar] [CrossRef] [PubMed]
- Pancrace, C.; Barny, M.A.; Ueoka, R.; Calteau, A.; Scalvenzi, T.; Pédron, J.; Humbert, J.-F.; Gugger, M. Insights into the Planktothrix Genus: Genomic and Metabolic Comparison of Benthic and Planktic Strains. Sci. Rep. 2017, 7, 41181. [Google Scholar] [CrossRef]
- Hahn, M.W.; Jezberová, J.; Koll, U.; Saueressig-Beckm, T.; Schmidt, J. Complete Ecological Isolation and Cryptic Diversity in Polynucleobacter Bacteria Not Resolved by 16S rRNA Gene Sequences. ISME J. 2016, 10, 1642–1655. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Li, Y.; Niu, L.; Zhang, W.; Wang, C.; Wang, P.; Meng, F. New Insights into the Spatial Variability of Biofilm Communities and Potentially Negative Bacterial Groups in Hydraulic Concrete Structures. Water Res. 2017, 123, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.; Acuña, V.; Sabater, S. Multiple Stressors Determine Community Structure and Estimated Function of River Biofilm Bacteria. Appl. Environ. Microbiol. 2020, 86, E00291-20. [Google Scholar] [CrossRef]
- Li, P.; Wang, C.; Liu, G.; Luo, X.; Rauan, A.; Zhang, C.; Li, T.; Yu, H.; Dong, S.; Gao, Q. A Hydroponic Plants and Biofilm Combined Treatment System Efficiently Purified Wastewater from Cold Flowing Water Aquaculture. Sci. Total Environ. 2022, 821, 153534. [Google Scholar] [CrossRef]
- Fournier, I.B.; Lovejoy, C.; Vincent, W.F. Changes in the Community Structure of Under-Ice and Open-Water Microbiomes in Urban Lakes Exposed to Road Salts. Front. Microbiol. 2021, 12, 660719. [Google Scholar] [CrossRef]
- Webster, T.M.U.; Consuegra, S.; de Leaniz, C.G. Early Life Stress Causes Persistent Impacts on the Microbiome of Atlantic Salmon. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100888. [Google Scholar] [CrossRef]
- Eberle, J.J.; Greenwood, D.R. Life at the Top of the Greenhouse Eocene World—A Review of the Eocene Flora and Vertebrate Fauna from Canada’s High Arctic. Bulletin 2012, 124, 3–23. [Google Scholar] [CrossRef]
- Swanson, H.K.; Kidd, K.A.; Reist, J.D. Effects of Partially Anadromous Arctic Charr (Salvelinus alpinus) Populations on Ecology of Coastal Arctic Lakes. Ecosystems 2010, 13, 261–274. [Google Scholar] [CrossRef]
- Corush, J.B. Evolutionary Patterns of Diadromy in Fishes: More than a Transitional State between Marine and Freshwater. BMC Evol. Biol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Laske, S.M.; Amundsen, P.A.; Christoffersen, K.S.; Erkinaro, J.; Guðbergsson, G.; Hayden, B.; Kahilainen, K.K.; Klemetsen, A.; Knudsen, R.; L’Abée-Lund, J.H.; et al. Circumpolar Patterns of Arctic Freshwater Fish Biodiversity: A Baseline for Monitoring. Freshw. Biol. 2022, 67, 176–193. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Ma, J.; Ding, Y.; Zhang, S. Phosvitin Plays a Critical Role in the Immunity of Zebrafish Embryos via Acting as a Pattern Recognition Receptor and an Antimicrobial Effector. J. Biol. Chem. 2011, 286, 22653–22664. [Google Scholar] [CrossRef]
- Pietrzak, E.; Mazurkiewicz, J.; Slawinska, A. Innate Immune Responses of Skin Mucosa in Common Carp (Cyprinus carpio) Fed a Diet Supplemented with Galactooligosaccharides. Animals 2020, 10, 438. [Google Scholar] [CrossRef]
- Mansson, M.; Nielsen, A.; Kjærulff, L.; Gotfredsen, C.H.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T.O. Inhibition of Virulence Gene Expression in Staphylococcus aureus by Novel Depsipeptides from a Marine Photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef]
- Element, G.; Engel, K.; Neufeld, J.D.; Casselman, J.M.; van Coeverden de Groot, P.; Greer, C.W.; Walker, V.K. Seasonal Habitat Drives Intestinal Microbiome Composition in Anadromous Arctic Char (Salvelinus alpinus). Environ. Microbiol. Rep. 2020, 22, 3112–3125. [Google Scholar] [CrossRef]
- Lai, K.P.; Lin, X.; Tam, N.; Ho, J.C.H.; Wong, M.K.-S.; Gu, J.; Chan, T.F.; Tse, W.K.F. Osmotic Stress Induces Gut Microbiota Community Shift in Fish. Environ. Microbiol. 2020, 22, 3784–3802. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, E.F.; Juurakko, C.L.; Engel, K.; Neufeld, J.D.; Casselman, J.M.; Greer, C.W.; Walker, V.K. Environmental Impacts on Skin Microbiomes of Sympatric High Arctic Salmonids. Fishes 2023, 8, 214. https://doi.org/10.3390/fishes8040214
Hamilton EF, Juurakko CL, Engel K, Neufeld JD, Casselman JM, Greer CW, Walker VK. Environmental Impacts on Skin Microbiomes of Sympatric High Arctic Salmonids. Fishes. 2023; 8(4):214. https://doi.org/10.3390/fishes8040214
Chicago/Turabian StyleHamilton, Erin F., Collin L. Juurakko, Katja Engel, Josh D. Neufeld, John M. Casselman, Charles W. Greer, and Virginia K. Walker. 2023. "Environmental Impacts on Skin Microbiomes of Sympatric High Arctic Salmonids" Fishes 8, no. 4: 214. https://doi.org/10.3390/fishes8040214
APA StyleHamilton, E. F., Juurakko, C. L., Engel, K., Neufeld, J. D., Casselman, J. M., Greer, C. W., & Walker, V. K. (2023). Environmental Impacts on Skin Microbiomes of Sympatric High Arctic Salmonids. Fishes, 8(4), 214. https://doi.org/10.3390/fishes8040214