Abstract
Ghrelin, glucagon-like peptide-1 (GLP-1), and peptide YY (PYY) are potent hormones mediating food intake according to the nutritional status in fish. However, limited information is available on these genes and their expression in response to nutrition in silver pomfret (Pampus argenteus). A comparison analysis revealed that ghrelin and GLP-1 were relatively conserved in marine fish. PYYa and PYYb shared a low identity and were clustered to different PYY branches. Ghrelin, pyya, and pyyb mRNAs were highly expressed in the brain, while glp-1 was highly expressed in the gills and liver. The ghrelin mRNA expression was relatively high an hour post-feeding and decreased after 3 to 72 h of fasting. The glp-1 mRNA expression was increased after 6 to 24 h of fasting. The pyya mRNA expression was increased after 72 h of fasting, while the pyyb expression remained stable during fasting. The diet with 14% lipid promoted the expressions of glp-1, pyya, and pyyb, but inhibited the ghrelin expression. The ghrelin expression in the intestine was increased after palmitic acid, oleic acid (OA), linoleic acid (LA), α-linolenic acid, eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA) incubation. OA decreased the expression of glp-1 and increased the expression of pyya. The expression of pyyb was downregulated by LA, DHA, and EPA. These results suggest the importance of ghrelin, glp-1, and pyy in coordinating food intake in response to fasting, dietary lipid concentration, and fatty acids in silver pomfret.
Key Contribution:
This study provides insight into how ghrelin, glp-1, and pyy respond to fasting, dietary lipid concentration, and fatty acids in silver pomfret.
1. Introduction
The intestine, as a key player in maintaining systemic energy balance [1], does not only absorb nutrients, but also senses nutritional value and transmits metabolic information to other tissues [2,3]. Appetite and feed intake are affected by signals from the intestine, inducing satiety and hunger. The intestine is the largest hormone producing organ, containing at least 30 hormones, some of which play a crucial role in the regulation of appetite, such as ghrelin, glucagon-like peptide-1 (GLP-1), and peptide YY (PYY) [4,5]. Ghrelin, originally reported to be present in the rat stomach [6], is the only known peripheral appetite stimulating hormone [7]. Previous studies have indicated that ghrelin regulates energy homeostasis and lipid metabolism in teleost [8]. Jönsson and coworkers demonstrated that ghrelin regulates appetite in Atlantic salmon (Salmo salar) [9] and rainbow trout (Oncorhynchus mykiss) [10]. Glucagon-like peptide-1 (GLP-1), another peptide that is found in the intestine [11], also mediates appetite, promoting satiety [12] and inhibiting food intake [13]. In teleost fish, GLP-1 could inhibit food intake in grass carp (Ctenopharyngodon idella) [13] and coho salmon (Oncorhynchus kisutch) [14]. Similarly, peptide YY (PYY), secreted by the intestine, is also an important appetite-regulating peptide [15]. In mammals, PYY acts as an indicator and anorexigenic signal [16]. The teleost PYY is comprised of two subtypes, PYYa, and PYYb [16]. Previous studies in goldfish (Carassius auratus), grass carp, Atlantic halibut (Hippoglossus hippoglossus), and Nile tilapia (Oreochromis niloticus) have demonstrated the appetite-inhibiting effect of PYYa and PYYb in teleost fish [17,18].
Food availability and diet composition are critical factors affecting the secretion of intestinal hormones and food intake. Fasting is a common state of many fish species in both well-managed aquaculture conditions [19] and in the wild [20]. Fish appetite could be increased by fasting [20] and their nutritional status could be regulated by alternation of the endocrine hormones [21]. The effect of fasting on fish appetite has been reported in Leopard Mandarin fish (Siniperca scherzeri) [22], rainbow trout [23], and Siberian sturgeon (Acipenser baeri) [24]. However, the physiological effects of fasting on the fish gut remain largely unknown. Previous studies have reported that short-term starvation increased lipid mobilization and utilization in silver pomfret (Pampus argenteus) [25]. Nevertheless, how starvation affects appetite is not fully understood.
High fat diets (HFD) and vegetable oils (VO) are extensively used in aquaculture [26]. However, excessive fat in feed may suppress appetite, which will affect the metabolism and growth of fish and further limit the use of HFD in fish [27]. Previous studies have shown that HFD reduces food intake in rainbow trout [28], white seabass (Atractoscion nobilis) [29], grass carp [30], and blunt snout bream (Megalobrama amblycephala) [31]. In silver pomfret, excessive lipid intake can result in higher fat accretion and impaired growth performance [32]. However, it is not clear how and whether dietary fat concentration affects the appetite of silver pomfret. Therefore, it is meaningful to investigate the mechanism of the influence of HFD on food intake. Besides dietary lipid concentration, dietary fatty acid (FA) composition changes also influence fish appetite [33]. Oleate acid (OA), linoleic acid (LA), palmitic acid (PA), and α-linolenic acid (ALA) are common FAs of daily VO in the fish diet. Numerous studies in mammals have shown the connection between dietary FAs and appetite regulation [34,35]. However, at present, little information on the impact of intestinal fatty acid on fish appetite is available in the literature. OA and octanoate treatments activate the FA sensing system and inhibit food intake in rainbow trout [36]. OA, LA, and ALA regulate the agouti-related peptide 2, neuropeptide Y, and proopiomelanocortin genes’ expression and thus affect appetite in Chinese perch (Siniperca chuatsi) [37]. With HFD and various VO used for fish feed production, more attention should be paid to its intestinal and satiety effects.
Silver pomfret is a common marine fish with a high commercial and ecological value in China, India, Japan, and Kuwait [38]. In recent years, extensive studies on various aspects, such as the effects of lipid content and fatty acid composition on gonad development [39], growth, and proximate composition [40] and the effects of starvation on lipid mobilization and utilization [25], have been carried out to develop culture technology for silver pomfret. However, little is known about the nutritional requirement related to the appetite of this species, which is very important for developing the culture industry. This study was aimed at characterizing ghrelin, glp-1, pyya, and pyyb and examining their expression patterns in response to fasting, dietary lipid concentration, and FAs in the intestine of silver pomfret.
2. Materials and Methods
2.1. Fish
The procedures in this study were approved by the Ethical Committee of Ningbo University for animal experiments. Approval numbers: NBU20220079, approved on 7 March 2022. All of the silver pomfret juveniles (20.6 ± 0.7 g body weight) were purchased at a commercial farm in Xiangshan Bay, Ningbo, China. All of the fish were cultured in nine 500-L tanks for at least 2 weeks to adapt them to the aquarium environment. Fish were manually fed two times a day at 08:00 a.m. and 17:00 p.m. with commercial feed (larvae love 6#, Japan), until satiety. The experimental condition was controlled according to a previous study [41]. Throughout the experiment, the water temperature ranged between 25.5 °C and 27.5 °C, salinity was 24 ± 0.89%, and dissolved oxygen was 7.3 ± 0.49 mg/L. The body weight of the fish and the feeding conditions in the present study were the same as mentioned above, unless otherwise stated. The fish were anesthetized with 0.05% MS-222 (Sigma-Aldrich, San Luis, USA) and killed by spinal destruction before dissection of the tissues.
2.2. Open Reading Frame (ORF) Cloning of Ghrelin, glp-1, pyya, and pyyb
The ORF cloning process was conducted as in previous studies [42]. The total RNA was extracted from the tissues of three silver pomfret using a Trizol reagent Kit (Invitrogen, Carlsbad, CA, USA). The purified RNA was treated with DNase I (TaKaRa, Shiga, Japan) for removing genomic DNA. Complementary DNA (cDNA) samples were synthesized using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNA wiper) (Vazyme, Nanjing, China). Four pairs of primers were designed based on our previous transcriptome data (unpublished) (Table 1). Each of the 25 μL amplifications contained 12.5 μL 2 × dNTP Mix (TaKaRa, Shiga, Japan), 1.25 μL each of the forward and reverse primers (10 μM), 2.5 μL cDNA, and 7.5 μL ddH2O (TaKaRa, Shiga, Japan). The following PCR parameters were used: initial activation step at 94 °C for 5 min, followed by 94 °C for 30 s, 58 °C for 30 s, 72 °C for 30 s × 30, and 72 °C for 5 min. The PCR products were run on 1% agarose gel electrophoresis, and then the target bands were extracted using the FastPure Gel DNA Extraction Mini Kit (Vazyme, Nanjing, China). The PCR fragments were ligated into pMDTM 18-T vector (TaKaRa, Shiga, Japan). The recombinant plasmids were sequenced by Zhejiang Youkang Biotechnology Co., Ltd. (Hangzhou, China).

Table 1.
Primer oligonucleotide sequences and their applications.
2.3. Sequences Analysis
Multiple sequence alignments of deduced amino acid sequences from different species were performed using DNAMAN software. A phylogenetic tree based on the amino acid sequences was constructed using the neighbor-joining method of the Clustal W (http://www.ddbj.nig.ac.jp/search/clustalw-e.html; accessed on 8 July 2022) [44] and MEGA 7 program (http://www.megasoftware.net/index.html; accessed on 8 July 2022) [45]. One thousand bootstraps were performed to check the credibility of each branch. The accession number of the amino acid sequences used for the alignments and phylogenetic analysis are listed in the legends of the Figure 1 and Figure 2.

Figure 1.
Multiple amino acid sequence alignment of (A) ghrelin, (B) GLP-1, (C) PYYa and PYYb. Amino acids that are blacked indicate that amino acids in that column are identical across all species. Residues that are conserved in more than half of the listed peptides are shadowed. GenBank accession numbers: (A) SI (Silver pomfret) ghrelin; YA (Ya-fish) ghrelin (AEZ36050.1); CR (Crucian carp) ghrelin (ADP55075.1); CA (Grass carp) ghrelin (AFI98391.1); ZE (Zebrafish) ghrelin (ACJ76435.1); PA (Pacific bluefin tuna) ghrelin (BAM34457); LA (Largemouth bass) ghrelin (ACJ13547); EU (European seabass) ghrelin (ABG49130); MO (Mouse) ghrelin (AAI32231.1); HU (Human) ghrelin (ADM33790.1); (B) SI (Silver pomfret) GLP-1; MU (Mullet) GLP-1 (XP_047432601.1); BU (Blue tilapia) GLP-1 (XP_031584898.2); MA (Mandarin fish) GLP-1 (XP_044044264.1); HU (Humphead wrasse) GLP-1 (XP_041638161.1); SW (Swordfish) GLP-1 (XP_039997917.1); CL (Clownfish) GLP-1 (XP_023121936.1); BL (Bluefin tuna) GLP-1 (XP_042261152.1); ZE (Zebrafish) GLP-1 (NP_001258699.1); HM (Human) GLP-1 (NP_002045); (C) SI (Silver pomfret) PYYa; SI (Silver pomfret) PYYb; PE (European Perch) PYYa (XP_039682240.1); ZE (Zebrafish) PYYa (Q9I8P2.1); GO (Goldfish) PYY (ACJ03593.1); GR (Grass carp) PYY (AFE89419.1); MO (Mouse) PYY (AAH10821.1); HU (Human) PYY (P10082.3); FR (African clawed frog) PYY (XP_018092481.2); LA (Large yellow croaker) PYY (XP_019126422.1); SE (Senegalese sole) PYYa (AML27066.1), and CH (Chicken) PYY (P29203.1).
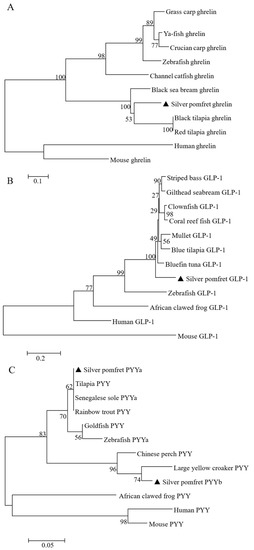
Figure 2.
Phylogenetic analysis of (A) ghrelin, (B) GLP-1, (C) PYYa and PYYb amino acid sequences. Scale bar indicates the substitution rate per residue. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. GenBank accession numbers: (A) Grass carp ghrelin (AFI98391.1); Ya-fish ghrelin (AEZ36050.1); Black sea bream ghrelin (AAV65508); Black tilapia ghrelin (BAC55160.1); Red Tilapia ghrelin (BAC65151.1); Channel catfish ghrelin (NP_001187244.1); Crucian carp ghrelin (ADP55075.1); Zebrafish ghrelin (ACJ76435.1); Mouse ghrelin (AAI32231.1); Human ghrelin (ADM33790.1); (B) Clownfish GLP-1 (XP_023121936.1); Coral reef fish GLP-1 (XP_022071913.1); Striped bass (XP_035533384.1); Bluefin tuna GLP-1 (XP_042261152.1); Mullet GLP-1 (XP_047432601.1); Blue tilapia GLP-1 (XP_031584898.2); Gilthead seabream GLP-1 (XP_030266058.1); Zebrafish GLP-1 (NP_001258699.1); Mouse GLP-1 (NP_001096638); Human GLP-1 (NP_002045); African clawed frog GLP-1 (NP_001079142.1); (C) Tilapia PYY (XP_003446444.1); Senegalese sole PYYa (AML27066.1); Zebrafish PYYa (AAF79942.1); Goldfish PYY (ACJ03593.1); African clawed frog PYY (XP_018092481.2); Human PYY (CAG46926.1); Mouse PYY (AAH10821.1); Chinese perch PYY (XP_044063373.1); Large yellow croaker PYY (XP_019126422.1), and Rainbow trout (PYY P69093.1).
2.4. Feeding Experiment
After acclimation, 36 healthy fish of a similar size were distributed into three tanks (12 fish per tank). The fish were fed manually until apparent satiation. The foreguts were collected 1 h (1 h), 3 h, 6 h, 12 h, 24 h, and 72 h after feeding. For each sampling point, the foreguts (two fish per tank) were excised, snap-frozen in liquid nitrogen, and stored at −80 °C for the next analysis.
2.5. Lipid Concentrition Experiment
The CON diet was a commercial feed and contained 5.0% carbohydrate, 52.1% protein, and 7.9% lipid. The diet with 14% lipid was made by mixing 91.3% (w/w) CON diet with 8.7% (w/w) soybean oil [46]. It finally contained 4.6% carbohydrate, 49.4% protein, and 14.4% lipid. The crude protein, crude lipid, and ash content in the diets were determined according to the methods of AOAC [47]. Briefly, the crude protein content was determined via the Dumas combustion methods with a protein analyzer (FP-528, Leco, Chicago, IL, USA). The crude lipid contents were determined by the ether extraction method using a Soxtec System HT (Soxtec System HT6, Tecator, Hoganas, Sweden). The ash contents were determined using a muffle furnace run at 550 °C for 8 h.
Thirty-six fish were randomly maintained in six tanks. Two diets were divided into triplicate tanks. In this experiment, the fish were fed until apparent satiation three times at 08:00 a.m., 14:00 p.m., and 20:00 p.m. Two hours after the last feeding, the foreguts from three fish per tank were excised, snap-frozen in liquid nitrogen, and stored at −80 °C for the next analysis.
2.6. FAs Incubation
Palmitic acid (PA, C16:0), oleic acid (OA, C18:1n–9), linoleic acid (LA, 18:2n–6), α-linolenic acid (ALA, 18:3n–3), eicosapentaenoic acid (EPA, 20:5n–3), and docosahexaenoic acid (DHA, 22:6n–3) were purchased from Aladdin, China. Six FAs were separately dissolved into bovine serum albumin (BSA, Sigma-Aldrich, San Luis, MO, USA) and configured into six different FA-BSA mother liquors with a concentration of 10 mM. The intestinal tissue was cultured as described previously in goldfish [48], with slight modifications. Briefly, 14 silver pomfret were fasted for 24 h prior to the experiment. The foreguts of the silver pomfret were dissected out and cut into small pieces of approximately 1–2 mm width. The intestinal pieces were washed and placed in Leibovitz’s L-15 medium (L-15, Hyclone, Logan, Utah, USA) containing 200 U/mL penicillin−streptomycin (Solarbio, Beijing, China). Then, the tissues were distributed into six-well culture plates (foreguts of two fishes/three wells/group) (Jet Biofil, Guangzhou, China). The tissues were preincubated for an hour in L-15 medium containing 44 mM sodium bicarbonate, 1% penicillin−streptomycin, and 0.05% gentamicin at 23 °C under an air environment containing 5% CO2 and 95% O2. Then, six FA-BSAs and serum (as control) were added to the L-15 culture mediums, respectively, and the final concentration of six FA-BSAs was 200 μM and the serum was 0.4% [43]. After 2 h of incubation, intestine samples were collected and maintained at −80 °C for the next analysis.
2.7. Quantitative Real-Time PCR (qPCR)
The extraction and purification of the total RNA from the tissues were done as described above. Then, the cDNA samples were synthesized using a PrimeScriptTM RT Reagent Kit (TaKaRa, Shiga, Japan). The sequences of the specific primers designed for each target gene are shown in Table 1. The qPCR amplification systems and reaction conditions were conducted as in a previous study [43]. The relative gene expression levels were calculated by the 2−ΔΔCT method, and β-actin was selected as the internal control.
2.8. Data Analysis and Statistics
The FA incubation assays and feeding experiments were analyzed by one-way analysis of variance (ANOVA) followed by Duncan’s multiple range tests. Tissue distribution and fasting experiments were examined with Levene’s test, followed by Tukey’s multiple range test. Statistical analysis was performed using SPSS 20.0 statistical software package (SPSS, Inc., Chicago, IL, USA). Data were log-transformed when equal variances were not assumed. The data were presented as means ± SEM, and differences were considered statistically significant at the * p < 0.05 level.
3. Results
3.1. Molecular Characterization of Silver Pomfret Ghrelin, glp-1, pyya, and pyyb
The amino acid sequence of the silver pomfret ghrelin showed a high identity with the ghrelin from Pacific bluefin tuna (Thunnus orientalis) (83%) and largemouth bass (Micropterus salmoides) (83%) (Figure 1A). Silver pomfret GLP-1 shared the highest amino acid sequence similarity with bluefin tuna (Thunnus thynnus) GLP-1 (90%), followed by mandarin fish (Siniperca chuatsi) GLP-1 (89%) and swordfish (Xiphias gladius) GLP-1 (89%) (Figure 1B). Silver pomfret PYYa showed the highest identity with the sequences of European perch (Perca fluviatilis) PYYa (95%). PYYb showed the highest identity with the sequences of large yellow croaker (Pseudosciaena crocea) PYY (84%). However, the sequence identity between PYYa and PYYb was 54% in silver pomfret (Figure 1C). The phylogenetic analysis showed that the silver pomfret ghrelin were clustered most tightly with marine fish ghrelin, and then with freshwater fish proteins into a ghrelin clade, which was clearly distinguished from mammalian ghrelin. Teleost GLP-1 was divided from the non-fish cluster. Silver pomfret PYYa and PYYb were clustered to different PYY branches, respectively, and then with other teleosts PYY into a clade, distinguished from the PYY from amphibian and mammals (Figure 2).
3.2. Tissue Distribution of Silver Pomfret Ghrelin, glp-1, pyya, and pyyb
The present results detected that ghrelin mRNA was ubiquitously distributed in the tested tissues and relatively highly expressed in the brain (Figure 3A). glp-1 was relatively highly expressed in the gills and liver, but was also weakly expressed in the kidney, heart, intestine, brain, spleen, and muscle (Figure 3B). The brain was the organ with the highest expression of pyya mRNA, followed by the intestine, and the heart had the lowest expression (Figure 3C). pyyb mRNA existed extensively among the tissues, with the highest levels in the brain, followed by the liver, gill, kidney, heart, intestine, spleen, and muscle (Figure 3D).
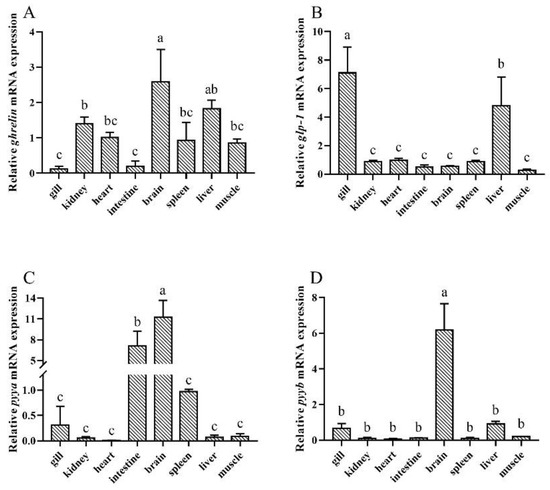
Figure 3.
Tissue distribution of (A) ghrelin, (B) glp-1, (C) pyya, and (D) pyyb of silver pomfret (Pampus argenteus). Bars with different letters represent significant differences between different tissues (p < 0.05).
3.3. Effects of Fasting on Ghrelin, glp-1, pyya, and pyyb Expression
The ghrelin mRNA expression was relatively high at an hour post-feeding. In addition, the ghrelin mRNA expression decreased after 3 h of fasting and remained until 72 h of fasting (Figure 4A). The glp-1 mRNA expression began to increase after 6 h of fasting and peaked after 24 h of fasting, but decreased significantly after 72 h of fasting (Figure 4B). The mRNA levels of pyya in the intestine peaked after 72 h of fasting (Figure 4C), while there were no significant differences in the expression of pyyb between any of the six sampling points (time) (Figure 4D).
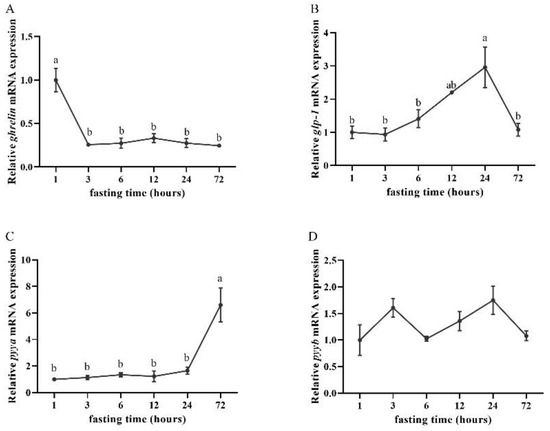
Figure 4.
Relative expression levels of (A) ghrelin, (B) glp-1, (C) pyya, and (D) pyyb mRNA of silver pomfret (Pampus argenteus) at different fasting times. Bars with different letters represent significant differences between different time points (p < 0.05).
3.4. Effects of Dietary Lipid Concentration on Ghrelin, glp-1, pyya, and pyyb Expression
In the intestine, the expression of ghrelin was higher in fish fed with 8% lipid diet compared with the fish fed with 14% lipid diet. On the contrary, when it came to glp-1, pyya, and pyyb mRNA, the 14% lipid diet groups had significantly higher expressions than the 8% lipid diet groups (Figure 5).
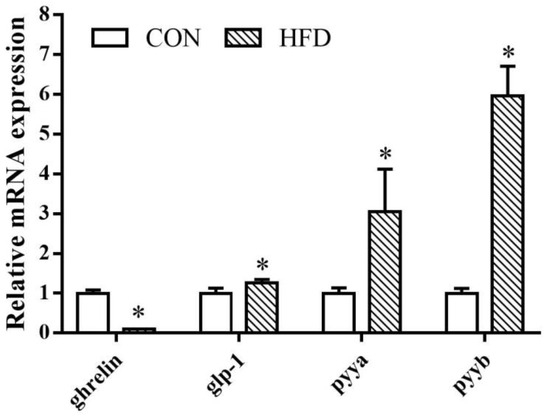
Figure 5.
Relative expression levels of ghrelin, glp-1, pyya, and pyyb in the intestine of silver pomfret (Pampus argenteus) fed diets with different fat contents. Asterisks represent significant differences between the 8% lipid diet groups and 14% lipid diet groups (* p < 0.05).
3.5. Effects of FAs Incubation on Ghrelin, glp-1, pyya, and pyyb Expression
Compared with the control group, ghrelin mRNA was decreased significantly in all FA-treated groups (Figure 6A). The expression of glp-1 was also clearly less detected in the OA-treated intestine (Figure 6B). The expression of pyya was significantly upregulated by OA treatment (Figure 6C). However, pyyb mRNA was highly expressed in the LA, DHA, and EPA incubated intestine (Figure 6D).
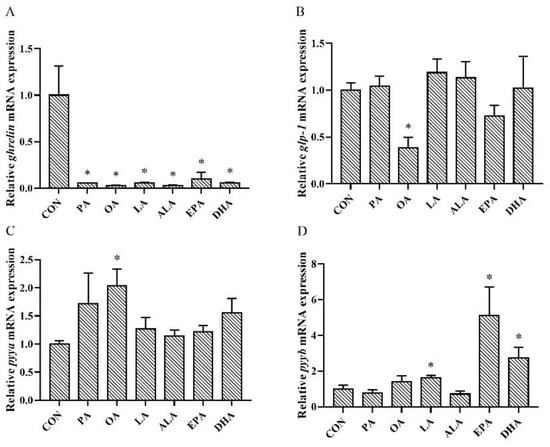
Figure 6.
Relative expression levels of (A) ghrelin, (B) glp-1, (C) pyya, and (D) pyyb in the intestine of silver pomfret (Pampus argenteus) incubated with FAs. Asterisks represent significant differences between the control groups and FAs treatment groups (* p < 0.05).
4. Discussion
In the present study, the amino acid sequence of silver pomfret ghrelin shared a high similarity with other teleost species. The phylogenetic analysis also showed that silver pomfret ghrelin belongs to a branch of teleost. These results indicate a high conservation of ghrelin in teleost [49]. The silver pomfret GLP-1 sequence in this study shared a high similarity with marine fish GLP-1 sequences. The phylogenetic analysis showed that silver pomfret GLP-1 was clustered most closely with the other marine fish GLP-1, but obviously diverged from tetrapod GLP-1. GLP-1 was relatively diverged among amphibians, while it was revealed to be more conserved among mammals [50]. The present results suggest that GLP-1 is relatively conserved in marine fish. Here, silver pomfret PYYa and PYYb had a low sequence identity and shared the highest degree of similarity with different vertebrate PYY. In addition, silver pomfret PYYa and PYYb were clustered to different teleost PYY subtypes. PYYa and PYYb showed a degree of similarity, likely due to their common origin from the same ancestor, as well as divergences because of continual evolution. Previous studies have revealed that PYYa has a high identity between cartilaginous fishes and bony fishes, but fish PYYb is relatively variable and has a rapid rate of evolution [16]. Considering the differences between silver pomfret PYYa and PYYb, it could be speculated that PYYa and PYYb may be responsible for different metabolic process and this might be reflected in their tissue distribution. Expression divergence reflects the functional divergence between duplicate genes [51].
Similar to the tissue expression patterns in other teleost fish, ghrelin mRNA was ubiquitously expressed in silver pomfret [52], which indicates the potential role on both systemic (endocrine) and local (autocrine/paracrine) action of ghrelin [53]. Although there is insufficient information on the expression of the ghrelin in numerous organs, the widespread distribution of ghrelin mRNA in silver pomfret, as in many other fish, suggests that ghrelin may have multiple tissue-specific activities in fish. We observed that glp-1 was mainly expressed in the liver and gills, in which a high glucose metabolism rate has been reported [54]. GLP-1 is involved in glucose metabolism as a potent regulatory factor [55], suggesting a corresponding function in silver pomfret. The expression of the glp-1 gene in the brain and intestine also suggests that it, similar to other fish, may participate in the regulation of appetite through the gut-brain axis [56]. The highest expression of pyya and pyyb mRNA were both observed in the brain, which is consistent with the pattern in grass carp [57] and goldfish [58]. It seems reasonable to think that the brain is one of the major organs of pyy expression and distribution in teleosts. Thus, it also reflects the effect of PYY on the central nervous system, such as the inhibition of feeding and pituitary hormone secretion in fish.
The gut-brain axis is a complex bidirectional communication system between the gastrointestinal tract and the central nervous system [56]. Within this axis, the gastrointestinal tract sends information to the brain, which transmits feedback to the gastrointestinal tract. The gut was chosen for the experiment because it senses changes in nutritional status more directly than the brain. Ghrelin mRNA expression decreased after 3 h of fasting and remained stable after 3 to 72 h of fasting. This was similar to previous studies in Atlantic Salmon, which showed no transcriptional differences of ghrelin between short-term fasted and fed groups [9]. However, a significant reduced expression of ghrelin was observed in goldfish, Atlantic salmon, and grass carp after fasting [59,60,61]. Ghrelin mRNA expression during short-term fasting in different fish may be species-specific. Ghrelin, as an orexigenic signal, is likely to be rapidly induced and relatively short lived. It is also possible that a duration longer than 72 h (tested here) would be needed to see changes in the ghrelin expression levels. Increased glp-1 expression levels were detected 12 h after fasting and reached a maximum 24 h after fasting. In addition, an obvious decrease in glp-1 mRNA expression was observed 72 h after fasting. However, the expression of glp-1 mRNA was significantly upregulated in the first hour of fasting and gradually decreased in the next 4 h of fasting in rats [62]. The changes in glp-1 expression levels may be related to the changes in food storage and energy of silver pomfret. Fish may still have food residues in their stomach within 24 h of eating, which promotes the response of glp-1 expression levels. In the present study, the expression of pyya remained stable during 24 h of fasting, but increased significantly after 72 h of fasting. This is similar to previous studies by Yuan et al. [63], who observed the pyy mRNA expression in ya-fish (Schizothorax prenanti) starved for 3 days was significantly higher than in the fish starved for 1 day. However, in the present data, the pyyb mRNA expression remained stable during 72 h of fasting. Similarly, in previous studies, a stable expression was observed in the pyy mRNA in Spotted scat (Scatophagus argus) [15], goldfish [58], and David’s schizothoracin (Schizothorax davidi) [64] during short periods of fasting. Individual homologues, pyya and pyyb, in this study behaved differently and not all of them were affected by fasting, similar to what was previously reported in Atlantic halibut [18]. The present results indicate that PYYa and PYYb might play different roles in fasting. Considering the unchanged pyyb expression, it was probably PYYa but not PYYb that acted as the main endogenous PYY in short-term fasting.
As lipids support numerous physiological processes in fish [65], it is possible that the dietary lipid concentration is related to food intake in fish. In the current study, the ghrelin mRNA expression in fish fed the 8% lipid diet was significantly lower than fish fed the 14% lipid diet. However, Huang et al. reported that the ghrelin gene expression of the intestine in grass carp was not significantly influenced by high lipid (8% lipid) diets [30]. In mammals, previous studies have also reported that ghrelin expression was downregulated in the rat stomach with HFD feeding [66]. In the present study, glp-1, pyya, and pyyb mRNA expressions increased with the 14% lipid diet. Unlike the stimulation of fasting, the high-fat diet significantly up-regulated the expression of pyyb, and it was even more obvious than the responses of the other anorexia genes, pyya and glp-1. This means that the expression of different genes is related to the stimulus types, and pyyb has an obvious response to lipids. Contrary to the present results, Bonacic et al. observed decreased mRNA levels of glp-1, pyya, and pyyb in Senegalese sole (Solea senegalensis) fed an 18% lipid diet [67]. Previous studies on GLP-1 showed that rats fed a 20% lipid diet had numerically higher serum total GLP-1 concentrations than rats fed the 6% lipid diet [68]. Mul et al. reported that 81% lipid diet did not change the GLP-1 protein levels compared with the 17% lipid diet in rats [40]. In humans, Essah et al. found a higher serum PYY concentration after eating the test meal with a high lipid level, compared with the test meal with a low lipid level [69]. The increase in pyy mRNA expression may reflect a surge in circulating levels of PYY with HFD. The decreased ghrelin mRNA expression concomitant with the increased glp-1, pyya, and pyyb mRNA expressions probably induced the appetite suppression in the 14% lipid diet in silver pomfret.
Long chain FAs affect the growth and metabolism of fish [70]. However, limited information is available about the effects of FAs on intestinal hormones. Plasma ghrelin concentration was upregulated in ewes as the feed EPA and DHA concentrations increased [71]. Ghrelin mRNA expression increased in the pigeon (Columba Livia) intestine when treated with OA, LA, and ALA, while PA treatment had no effect [72]. The expression of preproghrelin in goldfish was mainly upregulated by OA, but downregulated by EPA and DHA [73]. In the current study, the expression of ghrelin was decreased in the intestine treated with PA, OA, LA, ALA, EPA, and DHA, indicating an inhibitory effect of these FAs on ghrelin secretion. glp-1 mRNA expression was decreased only in the OA-treated intestine. However, previous studies have shown that OA exposure causes murine enteroendocrine cells to release GLP-1 [73]. In the present study, an upregulation in pyya mRNA expression was observed in the intestine treated with OA. Meanwhile, the expression of pyyb mRNA appeared to be quite different, which was increased by LA, EPA, and DHA treatment rather than OA. A previous study observed an increase in PYY release in cells treated with OA [74]. Hand et al. observed in the intestinal STC-1 cells that LA incubation significantly stimulated PYY secretion, but DHA incubation markedly decreased PYY release [75]. Hand et al. also observed that over the 72-h period, OA, LA, ALA, EPA, and DHA increased PYY accumulation in the culture media [75]. GLP-1, PYYa, and PYYb are widely believed to play a role as anorexic factors, but they had a completely different expression in the same FAs treatment in this in vitro experiment. The reason for this phenomenon may be that GLP-1, PYYa, and PYYb have different sensitivities for FAs and that their expression in the present study might have been affected by other, undetermined, physiological processes. Considering that these genes have an obvious response to lipid concentration, fatty acid concentration may also be an important factor affecting gene expression. In addition, previous research regarding ALA reported a time-dependent effect of this fatty acid on feed intake [33]. Thus, the effect of fatty acid concentration and the length of incubation time on the appetite regulation of fish remains to be elucidated further.
5. Conclusions
In conclusion, ghrelin, glp-1, and pyya are likely involved in the regulation of appetite by fasting. Diet with 14% lipid probably suppresses the appetite of silver pomfret via promoting anorexigenic factor expression and inhibiting orexigenic factor expression. In addition, the expression of intestinal endocrine factors varied in response to different fatty acids, which may affect their utilization in silver pomfret. However, the regulation of external stimuli on fish feeding depends on the gut−brain axis. It is important and necessary to consider further research on how ghrelin, glp-1, and pyy respond to different nutritional situations in the brain and its underlying mechanisms.
Author Contributions
L.Z. (Lei Zhang) and P.S. conceived the project and designed the experiments. L.Z. (Lei Zhang) wrote the manuscript. P.S. performed the experiments. P.S. and L.Z. (Lei Zhang) analyzed the data. L.Z. (Lin Zhang), Y.W., X.Y. and J.X. provided the resources. The manuscript was revised and improved by K.L. and P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (grant numbers: 42076118 and 41706159); the Major Project of Science, Technology, and Innovation 2025 in Ningbo City (grant number: 2021Z003); and partly sponsored by K.C. Wong Magna Fund in Ningbo University.
Institutional Review Board Statement
The present studies in animals were reviewed and approved by the Committee for the Welfare and Ethics of Laboratory Animals of Ningbo University (approval numbers: NBU20220079, approved on 7 March 2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Yuxiang Zhu, Guochao Ye, Mengqi Zhang, and Lina Bai for their invaluable assistance with this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rehfeld, J.F. The New Biology of Gastrointestinal Hormones. Physiol. Rev. 1998, 78, 1087–1108. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Function and Mechanisms of Enteroendocrine Cells and Gut Hormones in Metabolism. Nat. Rev. Endocrinol. 2019, 15, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M.; Stakenborg, N.; Viola, M.F.; Boeckxstaens, G. Macrophages in the Gut: Masters in Multitasking. Immunity 2022, 55, 1530–1548. [Google Scholar] [CrossRef] [PubMed]
- McCarty, T.R.; Jirapinyo, P.; Thompson, C.C. Effect of Sleeve Gastrectomy on Ghrelin, GLP-1, PYY, and GIP Gut Hormones: A Systematic Review and Meta-Analysis. Ann. Surg. 2020, 272, 72. [Google Scholar] [CrossRef] [PubMed]
- Freire, R.H.; Alvarez-Leite, J.I. Appetite Control: Hormones or Diet Strategies? Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin Is a Growth-Hormone-Releasing Acylated Peptide from Stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Nunez-Salces, M.; Li, H.; Feinle-Bisset, C.; Young, R.L.; Page, A.J. The Regulation of Gastric Ghrelin Secretion. Acta Physiol. 2021, 231, e13588. [Google Scholar] [CrossRef]
- Zhong, H.; Hu, Y.; Yu, F. A Review on Ghrelin and Fish Reproduction. Reprod. Breed. 2021, 1, 128–135. [Google Scholar] [CrossRef]
- Del Vecchio, G.; Lai, F.; Gomes, A.S.; Verri, T.; Kalananthan, T.; Barca, A.; Handeland, S.; Rønnestad, I. Effects of Short-Term Fasting on MRNA Expression of Ghrelin and the Peptide Transporters PepT1 and 2 in Atlantic Salmon (Salmo Salar). Front. Physiol. 2021, 12, 666670. [Google Scholar] [CrossRef]
- Calo, J.; Soengas, J.L.; Pastor, J.J.; Blanco, A.M.; Morais, S. Evidence of Gastrointestinal Sensing and Gut-Brain Communication in Rainbow Trout (Oncorhynchus mykiss) in Response to the Aqueous Extract of Fishmeal and Its Free Amino Acid Fraction. Aquaculture 2023, 567, 739264. [Google Scholar] [CrossRef]
- Panaro, B.L.; Yusta, B.; Matthews, D.; Koehler, J.A.; Song, Y.; Sandoval, D.A.; Drucker, D.J. Intestine-Selective Reduction of Gcg Expression Reveals the Importance of the Distal Gut for GLP-1 Secretion. Mol. Metab. 2020, 37, 100990. [Google Scholar] [CrossRef] [PubMed]
- Hira, T.; Pinyo, J.; Hara, H. What Is GLP-1 Really Doing in Obesity? Trends Endocrinol. Metab. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Xie, Y.; Cheng, D.; Wang, L.; Chen, F.; Chen, H.; Ma, H.; Yang, Y.; Yuan, X. GLP-1 Responds to Postprandial Hyperglycemia by Reducing Transcription Level in Grass Carp (Ctenopharyngodon idella). Aquac. Rep. 2022, 23, 101045. [Google Scholar] [CrossRef]
- White, S.L.; Volkoff, H.; Devlin, R.H. Regulation of Feeding Behavior and Food Intake by Appetite-Regulating Peptides in Wild-Type and Growth Hormone-Transgenic Coho Salmon. Horm. Behav. 2016, 84, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Assan, D.; Mustapha, U.F.; Ndandala, C.B.; Li, Z.; Li, G.; Huang, C.; Zhao, G.; Chen, H.; Jiang, M. Molecular Cloning, Identification, and Expression Analysis of Pyy and Its Effect on Mechanisms Regulating Appetite in Spotted Scat, Scatophagus argus. Aquac. Rep. 2023, 28, 101465. [Google Scholar] [CrossRef]
- Assan, D.; Mustapha, U.F.; Chen, H.; Li, Z.; Peng, Y.; Li, G. The Roles of Neuropeptide Y (Npy) and Peptide YY (Pyy) in Teleost Food Intake: A Mini Review. Life 2021, 11, 547. [Google Scholar] [CrossRef]
- Nadermann, N.; Seward, R.K.; Volkoff, H. Effects of Potential Climate Change-Induced Environmental Modifications on Food Intake and the Expression of Appetite Regulators in Goldfish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 235, 138–147. [Google Scholar] [CrossRef]
- Gomes, A.S.; Lygre, E.; Harboe, T.; Zimmermann, F.; Jordal, A.-E.O.; Hamre, K.; Rønnestad, I. The Role of Cholecystokinin and Peptide YY in Feed Intake in Atlantic Halibut (Hippoglossus hippoglossus) Larvae. Neuropeptides 2022, 91, 102202. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, G.; Qu, L.; Zhong, X.; Gao, Y.; Ding, Z.; Xu, J.; Chen, X.; Cheng, H. Effect of Starvation on Intestinal Morphology, Digestive Enzyme Activity and Expression of Lipid Metabolism-Related Genes in Javelin Goby (Synechogobius hasta). Aquac. Res. 2022, 53, 87–97. [Google Scholar] [CrossRef]
- Kalananthan, T.; Folkedal, O.; Gomes, A.S.; Lai, F.; Handeland, S.O.; Tolås, I.; Gelebart, V.; Rønnestad, I. Impact of Long-Term Fasting on the Stomach-Hypothalamus Appetite Regulating Genes in Atlantic Salmon Postsmolts. Aquaculture 2023, 563, 738917. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Tang, N.; Wang, S.; Liu, Y.; Li, Y.; Liu, Y.; Zhang, S.; Li, Y.; Chen, D.; et al. Neuromedin U Regulates Food Intake of Siberian Sturgeon through the Modulation of Central and Peripheral Appetite Factors. Br. J. Nutr. 2023, 129, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-O.; Oh, S.-Y.; Kim, T. Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri. Animals 2022, 12, 889. [Google Scholar] [CrossRef]
- Mankiewicz, J.L.; Cleveland, B.M. Characterization of a Leptin Receptor Paralog and Its Response to Fasting in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2021, 22, 7732. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, M.; Li, Y.; Tang, N.; Zhang, X.; Chen, H.; Zhang, S.; Liu, Y.; Wang, J.; Chen, D.; et al. Cloning and Expression of Kiss Genes and Regulation of Feeding in Siberian Sturgeon (Acipenser baerii). Fish Physiol. Biochem. 2022, 48, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Meng, R.; Ran, Z.; Cheng, G.; Wang, Y.; Xu, J.; Xu, S.; Yan, X. Short-Term Starvation in Silver Pomfret (Pampus argenteus): Molecular Effects on Lipid Mobilization and Utilization. Aquac. Res. 2017, 48, 4874–4885. [Google Scholar] [CrossRef]
- Li, Y.; Liang, S.; Shao, Y.; Li, Y.; Chen, C.; You, C.; Monroig, Ó.; Rahimnejad, S.; Tocher, D.R.; Wang, S. Impacts of Dietary Konjac Glucomannan Supplementation on Growth, Antioxidant Capacity, Hepatic Lipid Metabolism and Inflammatory Response in Golden Pompano (Trachinotus ovatus) Fed a High Fat Diet. Aquaculture 2021, 545, 737113. [Google Scholar] [CrossRef]
- Tang, T.; Hu, Y.; Peng, M.; Chu, W.; Hu, Y.; Zhong, L. Effects of High-Fat Diet on Growth Performance, Lipid Accumulation and Lipid Metabolism-Related MicroRNA/Gene Expression in the Liver of Grass Carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 234, 34–40. [Google Scholar] [CrossRef]
- Gélineau, A.; Corraze, G.; Boujard, T.; Larroquet, L.; Kaushik, S. Relation between Dietary Lipid Level and Voluntary Feed Intake, Growth, Nutrient Gain, Lipid Deposition and Hepatic Lipogenesis in Rainbow Trout. Reprod. Nutr. Dev. 2001, 41, 487–503. [Google Scholar] [CrossRef]
- López, L.M.; Torres, A.L.; Durazo, E.; Drawbridge, M.; Bureau, D.P. Effects of Lipid on Growth and Feed Utilization of White Seabass (Atractoscion nobilis) Fingerlings. Aquaculture 2006, 253, 557–563. [Google Scholar] [CrossRef]
- Huang, C.; Sun, J.; Ji, H.; Oku, H.; Chang, Z.; Tian, J.; Yu, E.; Xie, J. Influence of Dietary Alpha-Lipoic Acid and Lipid Level on the Growth Performance, Food Intake and Gene Expression of Peripheral Appetite Regulating Factors in Juvenile Grass Carp (Ctenopharyngodon idellus). Aquaculture 2019, 505, 412–422. [Google Scholar] [CrossRef]
- Dai, Y.-J.; Jiang, G.-Z.; Yuan, X.-Y.; Liu, W.-B. High-Fat-Diet-Induced Inflammation Depresses the Appetite of Blunt Snout Bream (Megalobrama amblycephala) through the Transcriptional Regulation of Leptin/Mammalian Target of Rapamycin. Br. J. Nutr. 2018, 120, 1422–1431. [Google Scholar] [CrossRef]
- Hossain, M.A.; Almatar, S.M.; James, C.M. Effect of Varying Dietary Lipid Levels and Protein to Energy (P:E) Ratios on Growth Performance, Feed Utilization and Body Composition of Sub-Adult Silver Pomfrets, Pampus Argenteus (Euphrasen, 1788); Asian Network for Scientific Information: Punjab, Pakistan, 2011. [Google Scholar]
- Calo, J.; Conde-Sieira, M.; Comesaña, S.; Soengas, J.L.; Blanco, A.M. Fatty Acids of Different Nature Differentially Modulate Feed Intake in Rainbow Trout. Aquaculture 2023, 563, 738961. [Google Scholar] [CrossRef]
- Graybeal, A.J.; Shah, M.; Willis, J.L. Manipulation of Fatty Acid Composition in a High-Fat Meal Does Not Result in Differential Alterations in Appetite or Food Intake in Normal Weight Females: A Single-Blind Randomized Crossover Study. Appetite 2021, 160, 105085. [Google Scholar] [CrossRef]
- Behrouz, V.; Yari, Z. A Review on Differential Effects of Dietary Fatty Acids on Weight, Appetite and Energy Expenditure. Crit. Rev. Food Sci. Nutr. 2022, 62, 2235–2249. [Google Scholar] [CrossRef]
- Librán-Pérez, M.; Otero-Rodiño, C.; López-Patiño, M.A.; Míguez, J.M.; Soengas, J.L. Central Administration of Oleate or Octanoate Activates Hypothalamic Fatty Acid Sensing and Inhibits Food Intake in Rainbow Trout. Physiol. Behav. 2014, 129, 272–279. [Google Scholar] [CrossRef]
- Luo, H.; Liang, X.-F.; Li, J.; Zhang, Y.; Xiao, Q.; Peng, B.; Zhang, Z. Effect of Long-Chain Saturated and Unsaturated Fatty Acids on Hypothalamic Fatty Acid Sensing in Chinese Perch (Siniperca chuatsi). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 241, 110395. [Google Scholar] [CrossRef]
- Lan, W.; Che, X.; Xu, Q.; Wang, T.; Du, R.; Xie, J.; Hou, M.; Lei, H. Sensory and Chemical Assessment of Silver Pomfret (Pampus argenteus) Treated with Ginkgo Biloba Leaf Extract Treatment during Storage in Ice. Aquac. Fish. 2018, 3, 30–37. [Google Scholar] [CrossRef]
- Huang, X.; Yin, Y.; Shi, Z.; Li, W.; Zhou, H.; Lv, W. Lipid Content and Fatty Acid Composition in Wild-Caught Silver Pomfret (Pampus argenteus) Broodstocks: Effects on Gonad Development. Aquaculture 2010, 310, 192–199. [Google Scholar] [CrossRef]
- Hossain, M.A.; Almatar, S.M.; James, C.M. Effects of Varying Dietary Docosahexaenoic Acid Levels on Growth, Proximate Composition and Tissue Fatty Acid Profile of Juvenile Silver Pomfrets, Pampus Argenteus (Euphrasen, 1788). Aquac. Res. 2012, 43, 1599–1610. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, J.; Li, Y.; Zhang, M.; Jacques, K.J.; Gu, W.; Sun, Y.; Sun, J.; Yang, Y.; Xu, S.; et al. Immune Response of Silver Pomfret (Pampus argenteus) to Amyloodinium Ocellatum Infection. J. Fish Dis. 2021, 44, 2111–2123. [Google Scholar] [CrossRef]
- Xiong, X.; Yang, H.; Ding, C.; Qin, B.; Deng, Y.; Xiong, L.; Liu, X.; Li, Y.; Xiao, T.; Lv, Z. Functional and Expression Analysis Reveals the Involvement of Integrin Avβ3 in Antiviral Immunity of Grass Carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2022, 129, 52–63. [Google Scholar] [CrossRef]
- Shi, P.; Liao, K.; Xu, J.; Wang, Y.; Xu, S.; Yan, X. Eicosapentaenoic Acid Mitigates Palmitic Acid-Induced Heat Shock Response, Inflammation and Repair Processes in Fish Intestine. Fish Shellfish Immunol. 2022, 124, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D.; Begg, D.P.; Barrera, J.G.; Li, B.; Matter, E.K.; D’Alessio, D.A.; Woods, S.C.; Seeley, R.J.; Sandoval, D.A. High-Fat Diet Changes the Temporal Profile of GLP-1 Receptor-Mediated Hypophagia in Rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2013, 305, R68–R77. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated Software for Molecular Evolutionary Genetics Analysis and Sequence Alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef]
- Arias-Jayo, N.; Abecia, L.; Lavín, J.L.; Tueros, I.; Arranz, S.; Ramírez-García, A.; Pardo, M.A. Host-Microbiome Interactions in Response to a High-Saturated Fat Diet and Fish-Oil Supplementation in Zebrafish Adult. J. Funct. Foods 2019, 60, 103416. [Google Scholar] [CrossRef]
- Arlington, V. Official Methods of Analysis of Official Analytical Chemists International; Association of Official Analytical Chemists (AOAC) International: Gaithersburg, MD, USA, 1995. [Google Scholar]
- Blanco, A.M.; Bertucci, J.I.; Valenciano, A.I.; Delgado, M.J.; Unniappan, S. Ghrelin Suppresses Cholecystokinin (CCK), Peptide YY (PYY) and Glucagon-like Peptide-1 (GLP-1) in the Intestine, and Attenuates the Anorectic Effects of CCK, PYY and GLP-1 in Goldfish (Carassius auratus). Horm. Behav. 2017, 93, 62–71. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Wu, H.; Wu, Y.; Tang, N.; Chen, S.; Liu, Y.; Wang, J.; Zhang, X.; Li, Z. Ghrelin-Ghrelin Receptor (GSHR) Pathway via Endocannabinoid Signal Affects the Expression of NPY to Promote the Food Intake of Siberian Sturgeon (Acipenser baerii). Horm. Behav. 2022, 143, 105199. [Google Scholar] [CrossRef] [PubMed]
- Skovgaard, M.; Kodra, J.T.; Gram, D.X.; Knudsen, S.M.; Madsen, D.; Liberles, D.A. Using Evolutionary Information and Ancestral Sequences to Understand the Sequence–Function Relationship in GLP-1 Agonists. J. Mol. Biol. 2006, 363, 977–988. [Google Scholar] [CrossRef]
- Cheng, X.; Pan, M.; E, Z.; Zhou, Y.; Niu, B.; Chen, C. Functional Divergence of Two Duplicated Fertilization Independent Endosperm Genes in Rice with Respect to Seed Development. Plant J. 2020, 104, 124–137. [Google Scholar] [CrossRef]
- Dar, S.A.; Srivastava, P.P.; Varghese, T.; Gupta, S.; Gireesh-Babu, P.; Krishna, G. Effects of Starvation and Refeeding on Expression of Ghrelin and Leptin Gene with Variations in Metabolic Parameters in Labeo Rohita Fingerlings. Aquaculture 2018, 484, 219–227. [Google Scholar] [CrossRef]
- Muta, H.; Sugita, Y.; Furuta, T.; Shiimura, Y.; Ohshima, K.; Nakashima, K.; Sato, K.; Morioka, M.; Abe, H.; Nozawa, T.; et al. Expression of the Ghrelin/Growth Hormone Secretagogue Receptor Axis and Its Functional Role in Promoting Tumor Growth in Primary Central Nervous System Lymphomas. Neuropathology 2020, 40, 232–239. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional Regulation of Hepatic Glucose Metabolism in Fish. Fish Physiol. Biochem. 2009, 35, 519–539. [Google Scholar] [CrossRef]
- Hindsø, M.; Svane, M.S.; Hedbäck, N.; Holst, J.J.; Madsbad, S.; Bojsen-Møller, K.N. The Role of GLP-1 in Postprandial Glucose Metabolism after Bariatric Surgery: A Narrative Review of Human GLP-1 Receptor Antagonist Studies. Surg. Obes. Relat. Dis. 2021, 17, 1383–1391. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, J.; Luo, Z.; Cui, J.; Luo, Y.; Mak, S.O.K.; Wang, S.; Zhang, F.; Yang, Y.; So, K.-F.; et al. A Gut-Brain Axis Mediates Sodium Appetite via Gastrointestinal Peptide Regulation on a Medulla-Hypothalamic Circuit. Sci. Adv. 2023, 9, eadd5330. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, Y.; Pandit, N.P.; Fu, J.; Li, D.; Li, J. Molecular Cloning, Expression Analysis, and Potential Food Intake Attenuation Effect of Peptide YY in Grass Carp (Ctenopharyngodon idellus). Gen. Comp. Endocrinol. 2013, 187, 66–73. [Google Scholar] [CrossRef]
- Gonzalez, R.; Unniappan, S. Molecular Characterization, Appetite Regulatory Effects and Feeding Related Changes of Peptide YY in Goldfish. Gen. Comp. Endocrinol. 2010, 166, 273–279. [Google Scholar] [CrossRef]
- Unniappan, S.; Canosa, L.F.; Peter, R.E. Orexigenic Actions of Ghrelin in Goldfish: Feeding-Induced Changes in Brain and Gut MRNA Expression and Serum Levels, and Responses to Central and Peripheral Injections. Neuroendocrinology 2004, 79, 100–108. [Google Scholar] [CrossRef]
- Hevrøy, E.M.; Azpeleta, C.; Shimizu, M.; Lanzén, A.; Kaiya, H.; Espe, M.; Olsvik, P.A. Effects of Short-Term Starvation on Ghrelin, GH-IGF System, and IGF-Binding Proteins in Atlantic Salmon. Fish Physiol. Biochem. 2011, 37, 217–232. [Google Scholar] [CrossRef]
- Cai, W.; Yuan, X.; Yuan, Y.; Xie, S.; Gong, Y.; Su, H.; Qiao, Y. Sequence, Genomic Organization and Expression of Ghrelin Receptor in Grass Carp, Ctenopharyngodon idellus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 179, 54–61. [Google Scholar] [CrossRef]
- D’Alessio, D.; Lu, W.; Sun, W.; Zheng, S.; Yang, Q.; Seeley, R.; Woods, S.C.; Tso, P. Fasting and Postprandial Concentrations of GLP-1 in Intestinal Lymph and Portal Plasma: Evidence for Selective Release of GLP-1 in the Lymph System. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 293, R2163–R2169. [Google Scholar] [CrossRef]
- Yuan, D.; Zhou, C.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Wei, R.; Xin, Z.; Liu, J.; Gao, Y.; et al. Molecular Characterization and Tissue Expression of Peptide YY in Schizothorax Prenanti: Effects of Periprandial Changes and Fasting on Expression in the Hypothalamus. Regul. Pept. 2014, 190–191, 32–38. [Google Scholar] [CrossRef]
- Zhou, C.; Lei, L.; Yuan, D.; Deng, X.; Ye, H.; Luo, H.; Fang, J.; Yang, M.; Li, Y.; Zhang, C.; et al. Structural and Functional Characterization of Peptide YY on Feeding in Schizothorax Davidi. J. Exp. Zool. Part Ecol. Integr. Physiol. 2018, 329, 55–61. [Google Scholar] [CrossRef]
- Campos, I.; Matos, E.; Maia, M.R.G.; Marques, A.; Valente, L.M.P. Partial and Total Replacement of Fish Oil by Poultry Fat in Diets for European Seabass (Dicentrarchus labrax) Juveniles: Effects on Nutrient Utilization, Growth Performance, Tissue Composition and Lipid Metabolism. Aquaculture 2019, 502, 107–120. [Google Scholar] [CrossRef]
- Lee, H.-M.; Wang, G.; Englander, E.W.; Kojima, M.; Greeley, G.H., Jr. Ghrelin, A New Gastrointestinal Endocrine Peptide That Stimulates Insulin Secretion: Enteric Distribution, Ontogeny, Influence of Endocrine, and Dietary Manipulations. Endocrinology 2002, 143, 185–190. [Google Scholar] [CrossRef]
- Bonacic, K.; Martínez, A.; Gisbert, E.; Estévez, A.; Morais, S. Effect of Alternative Oil Sources at Different Dietary Inclusion Levels on Food Intake and Appetite Regulation via Enteroendocrine and Central Factors in Juvenile Solea Senegalensis (Kaup, 1858). Aquaculture 2017, 470, 169–181. [Google Scholar] [CrossRef]
- Charrier, J.A.; Martin, R.J.; McCutcheon, K.L.; Raggio, A.M.; Goldsmith, F.; Goita, M.; Senevirathne, R.N.; Brown, I.L.; Pelkman, C.; Zhou, J.; et al. High Fat Diet Partially Attenuates Fermentation Responses in Rats Fed Resistant Starch from High-Amylose Maize. Obesity 2013, 21, 2350–2355. [Google Scholar] [CrossRef]
- Essah, P.A.; Levy, J.R.; Sistrun, S.N.; Kelly, S.M.; Nestler, J.E. Effect of Macronutrient Composition on Postprandial Peptide YY Levels. J. Clin. Endocrinol. Metab. 2007, 92, 4052–4055. [Google Scholar] [CrossRef]
- Yu, J.; Wen, X.; You, C.; Wang, S.; Chen, C.; Tocher, D.R.; Li, Y. Comparison of the Growth Performance and Long-Chain Polyunsaturated Fatty Acids (LC-PUFA) Biosynthetic Ability of Red Tilapia (Oreochromis Mossambicus♀ × O. Niloticus♂) Fed Fish Oil or Vegetable Oil Diet at Different Salinities. Aquaculture 2021, 542, 736899. [Google Scholar] [CrossRef]
- Nickles, K.R.; Hamer, L.; Coleman, D.N.; Relling, A.E. Supplementation with Eicosapentaenoic and Docosahexaenoic Acids in Late Gestation in Ewes Changes Adipose Tissue Gene Expression in the Ewe and Growth and Plasma Concentration of Ghrelin in the Offspring1. J. Anim. Sci. 2019, 97, 2631–2643. [Google Scholar] [CrossRef]
- Xie, P.; Wan, X.P.; Bu, Z.; Zou, X.T. Molecular Cloning, Characterization, and Expression Analysis of Ghrelin and Cholecystokinin in the Pigeon (Columba Livia). Poult. Sci. 2016, 95, 2655–2666. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Canosa, L.F.; Unniappan, S. Glucose, Amino Acids and Fatty Acids Directly Regulate Ghrelin and NUCB2/Nesfatin-1 in the Intestine and Hepatopancreas of Goldfish (Carassius Auratus) in Vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 206, 24–35. [Google Scholar] [CrossRef]
- Bala, V.; Rajagopal, S.; Kumar, D.; Nalli, A.; Mahavadi, S.; Sanyal, A.; Grider, J.; Murthy, K. Release of GLP-1 and PYY in Response to the Activation of G Protein-Coupled Bile Acid Receptor TGR5 Is Mediated by Epac/PLC-ε Pathway and Modulated by Endogenous H2S. Front. Physiol. 2014, 5, 420. [Google Scholar] [CrossRef]
- Hand, K.V.; Bruen, C.M.; O’Halloran, F.; Panwar, H.; Calderwood, D.; Giblin, L.; Green, B.D. Examining Acute and Chronic Effects of Short- and Long-Chain Fatty Acids on Peptide YY (PYY) Gene Expression, Cellular Storage and Secretion in STC-1 Cells. Eur. J. Nutr. 2013, 52, 1303–1313. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).