

Spatial Density, Biomass, and Composition of Crustacean Zooplankton on Lake Michigan Beaches


Abstract
:1. Introduction
2. Materials and Methods
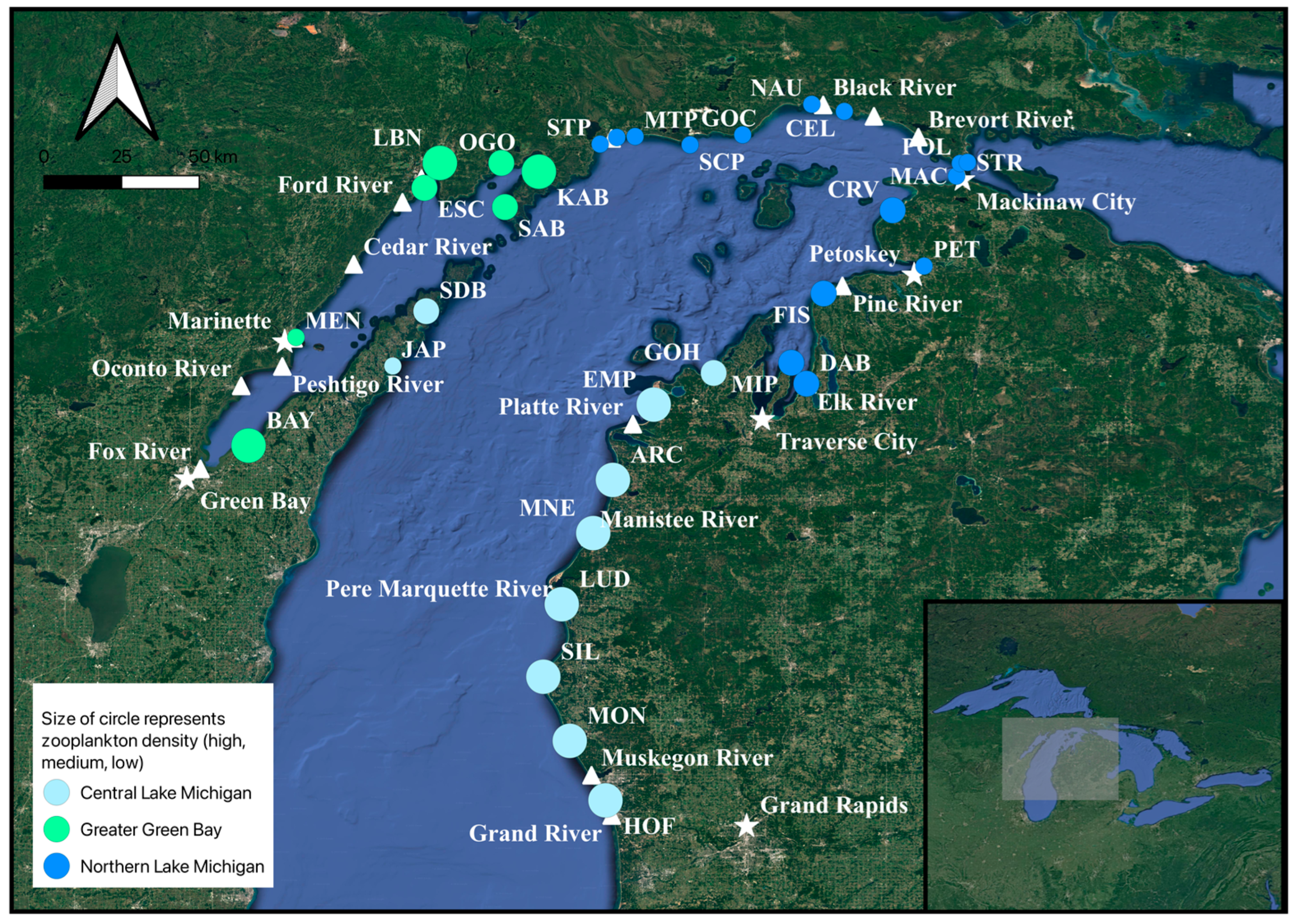
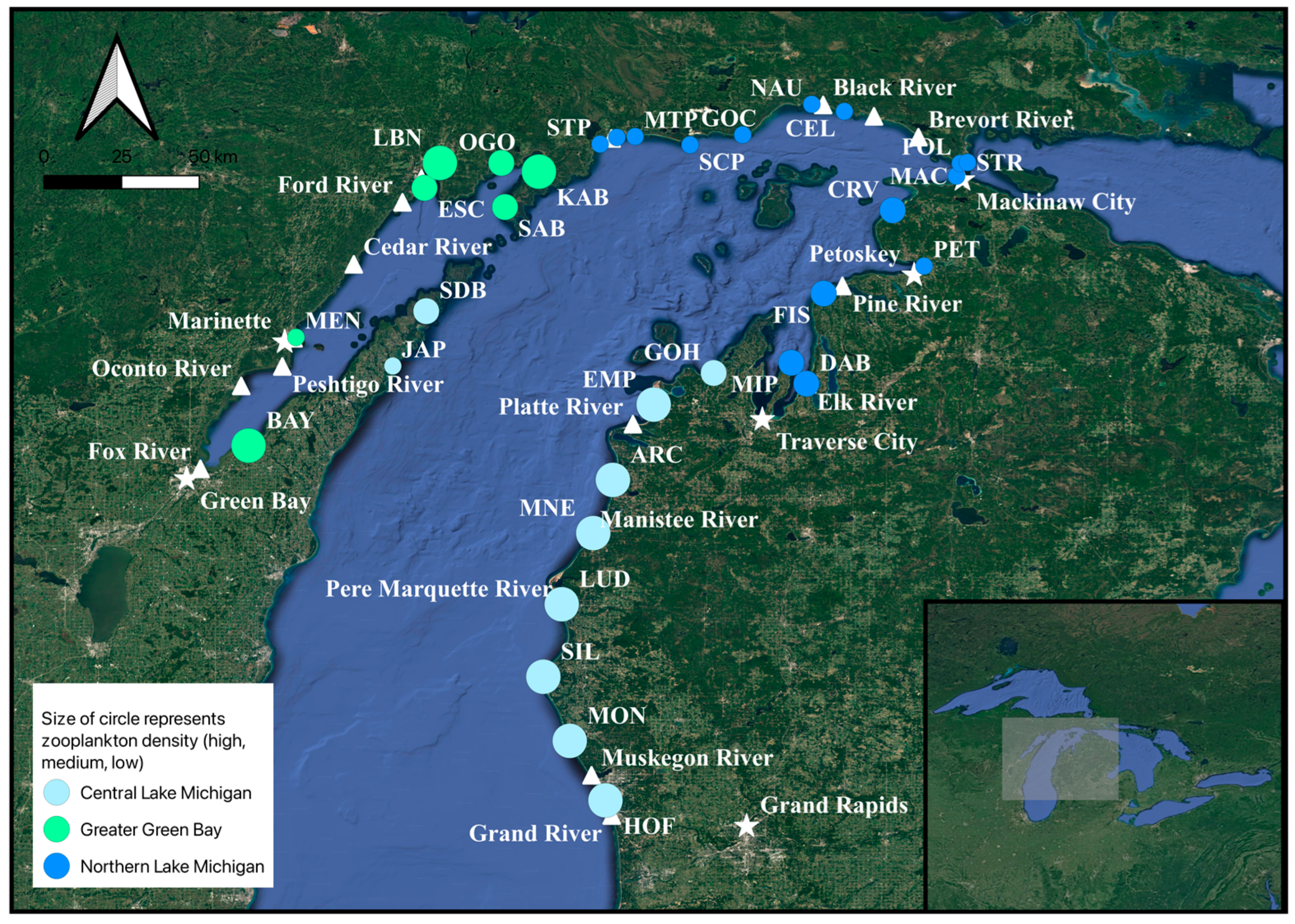
2.1. Sample Sites and Regions
2.2. Zooplankton Laboratory Processing
2.3. Statistical Analyses
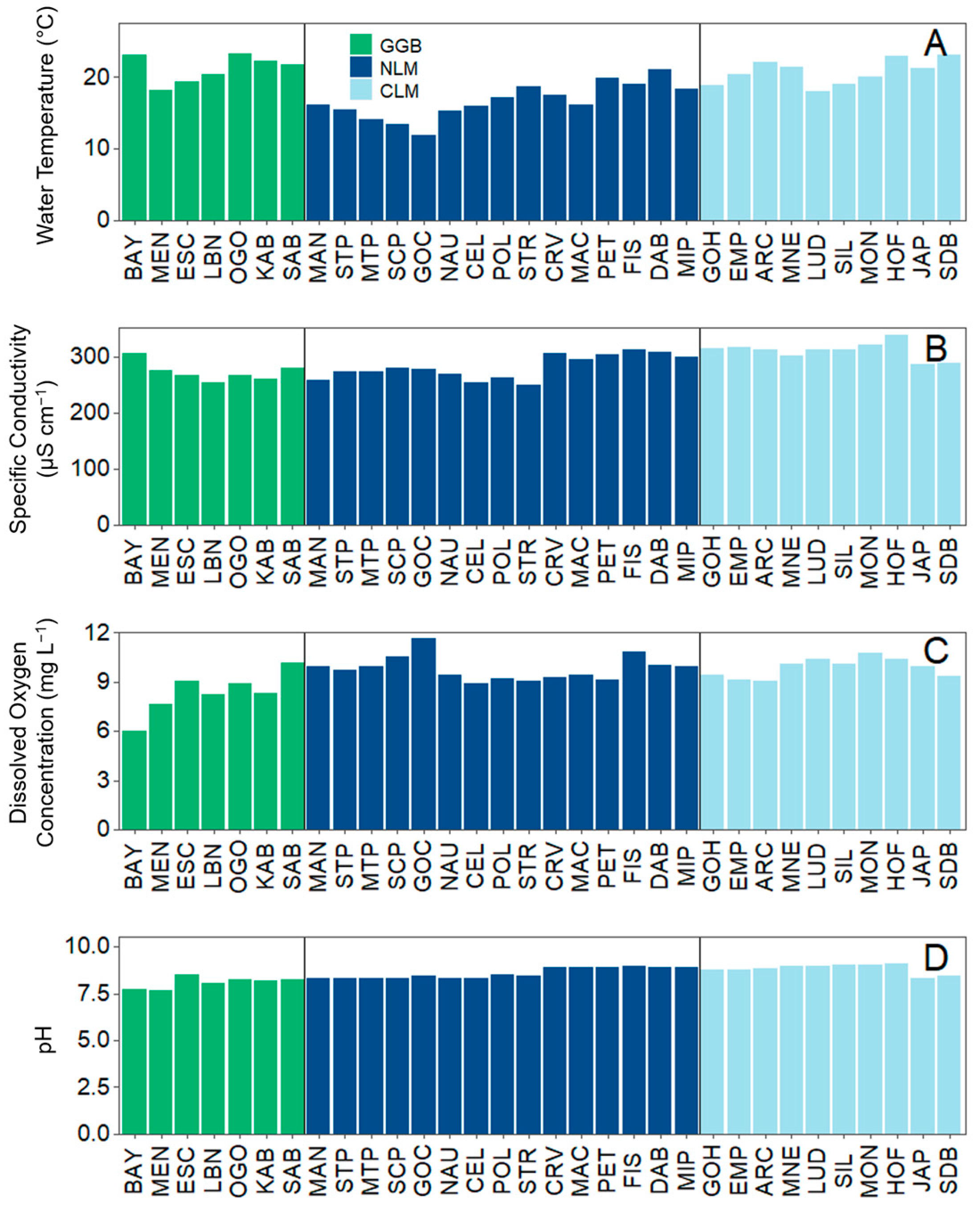
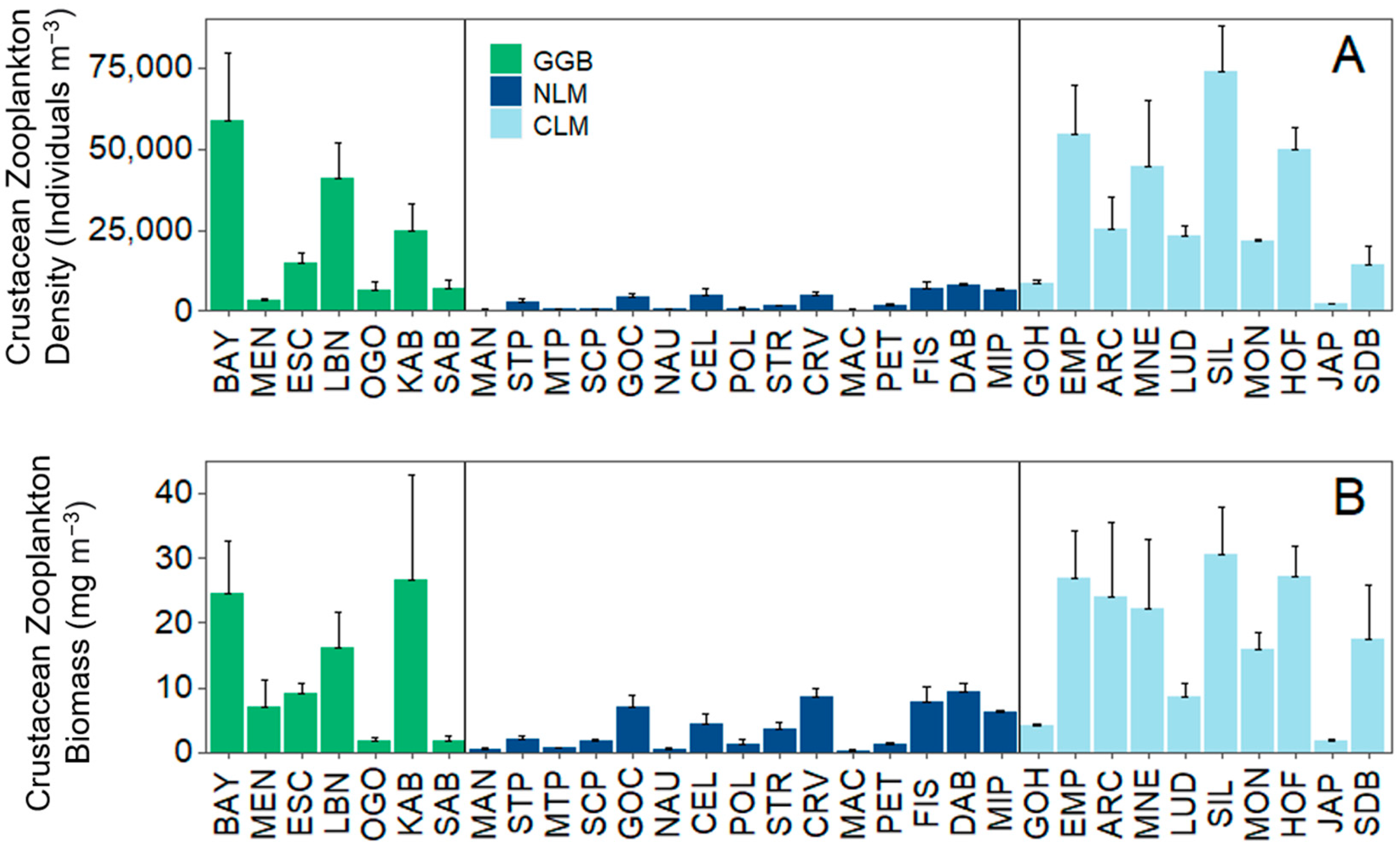
2.3.1. Analysis 1: Environmental Data and Total Crustacean Zooplankton Density and Biomass
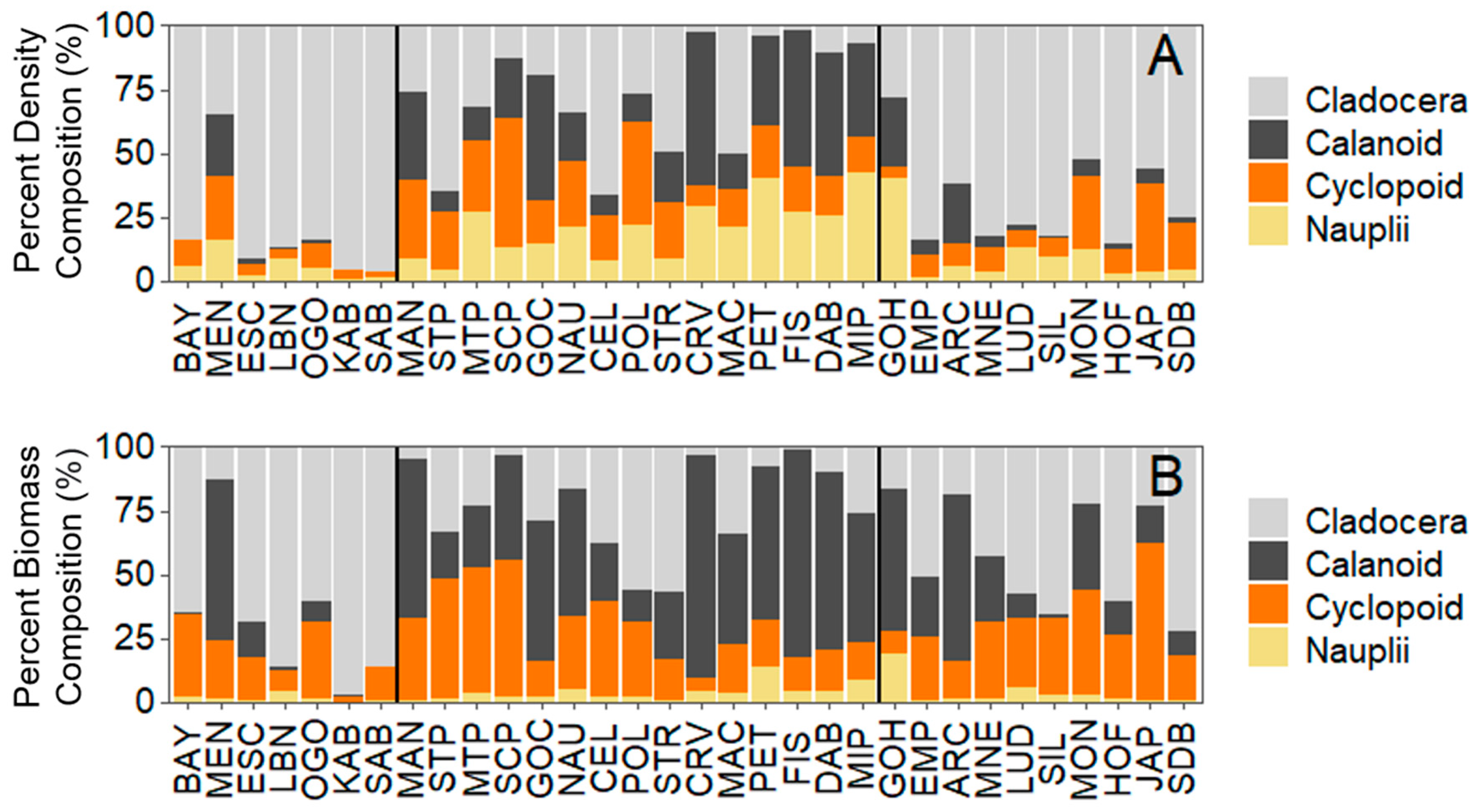
2.3.2. Analysis 2: Zooplankton Community Composition
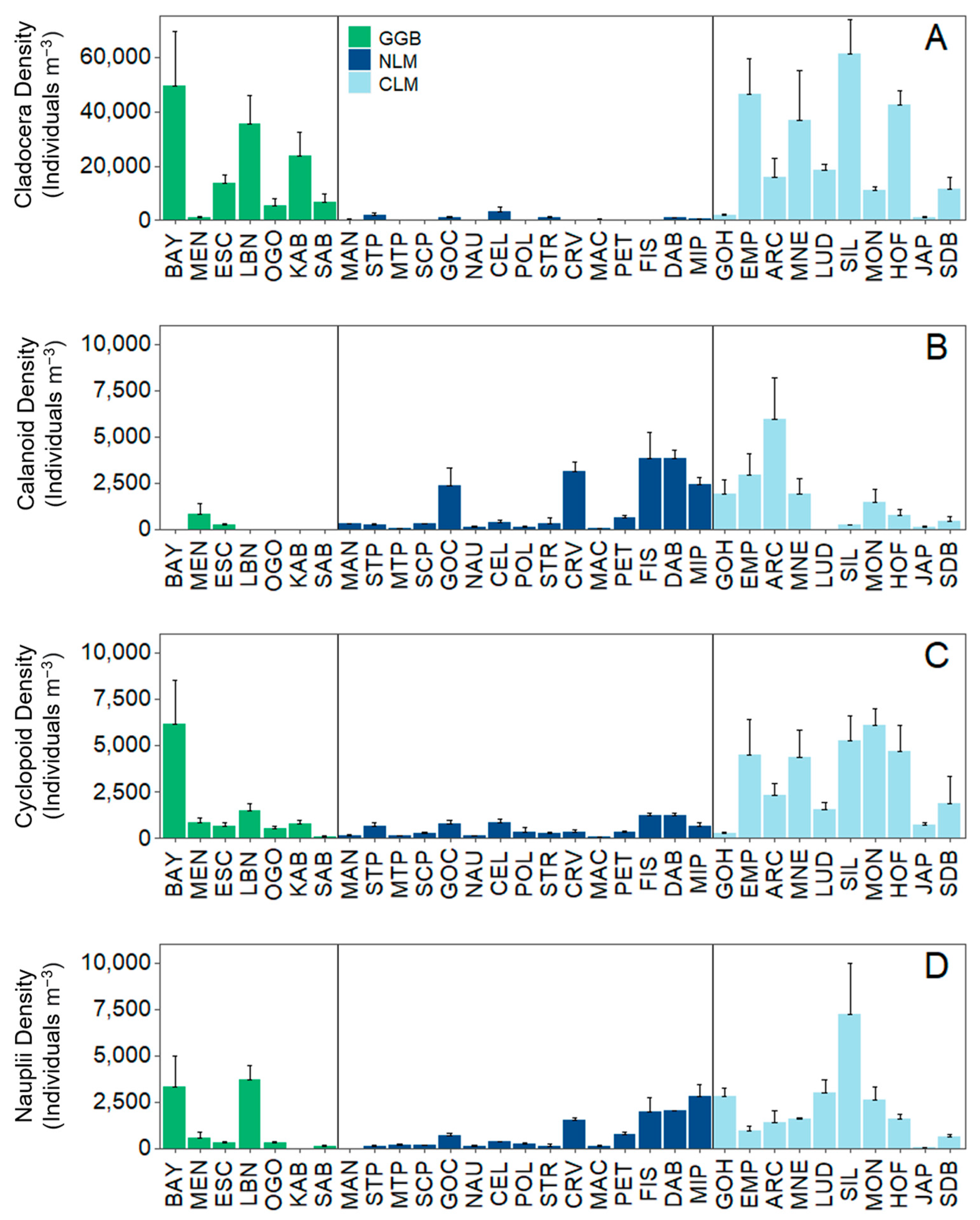
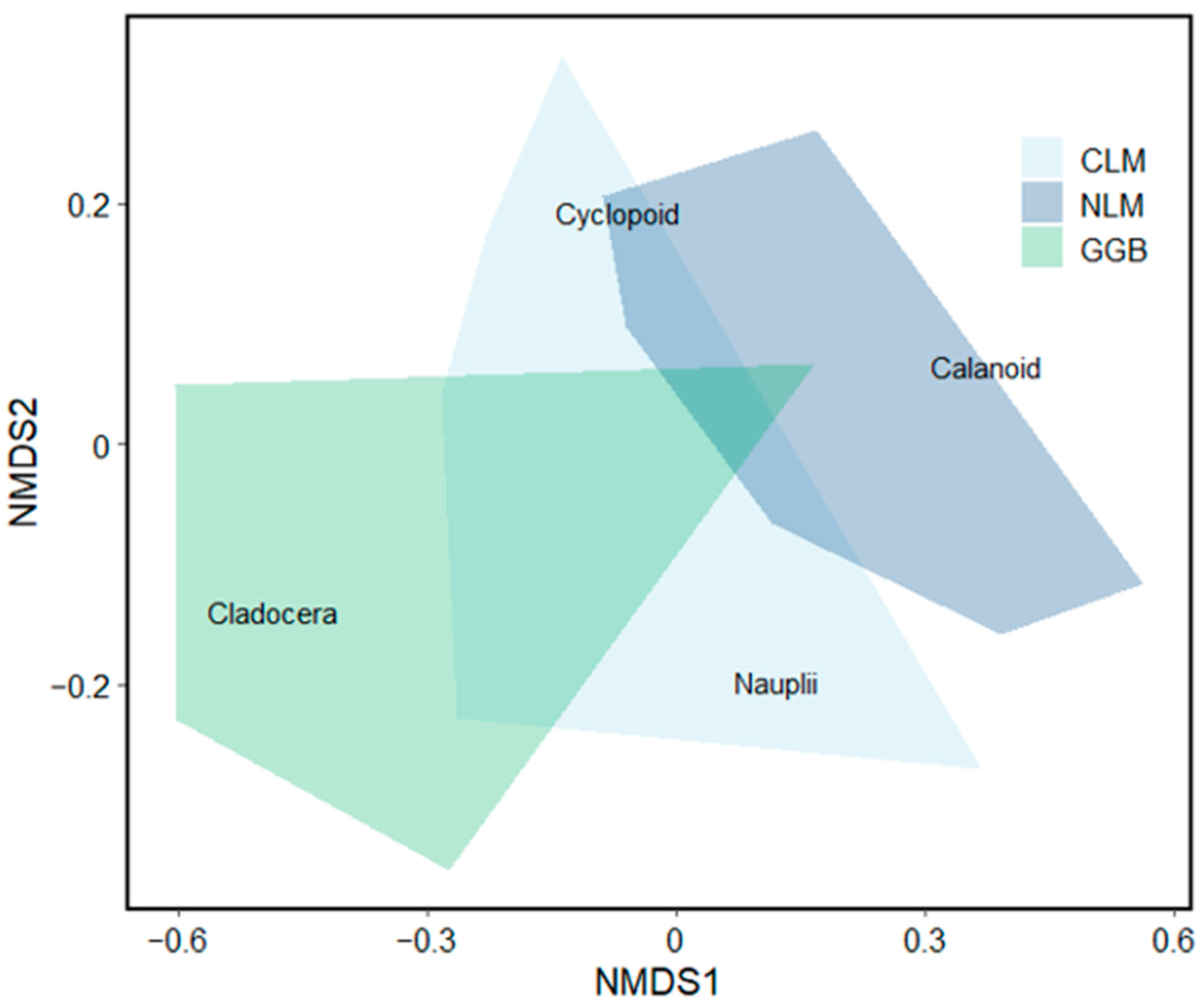
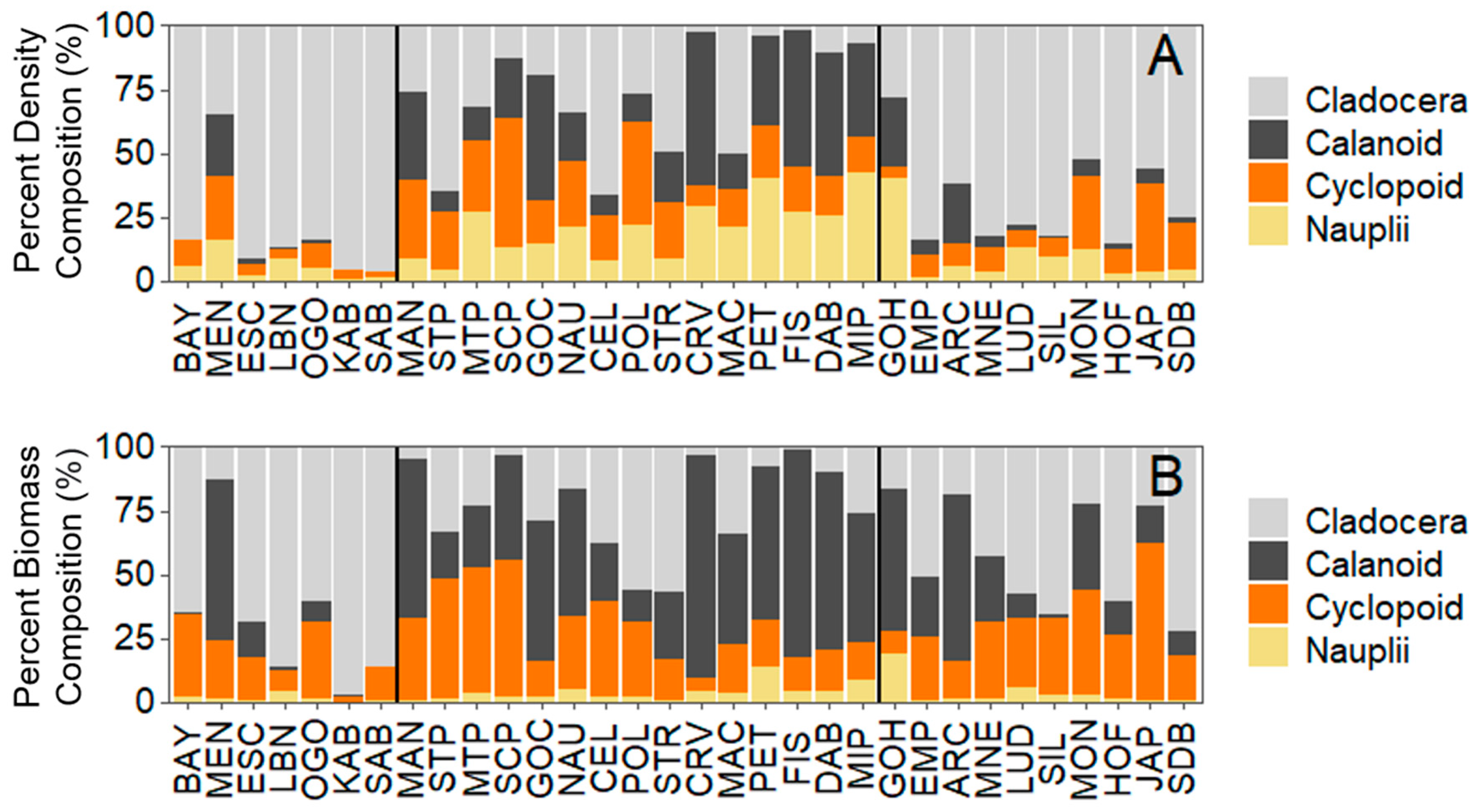
3. Results
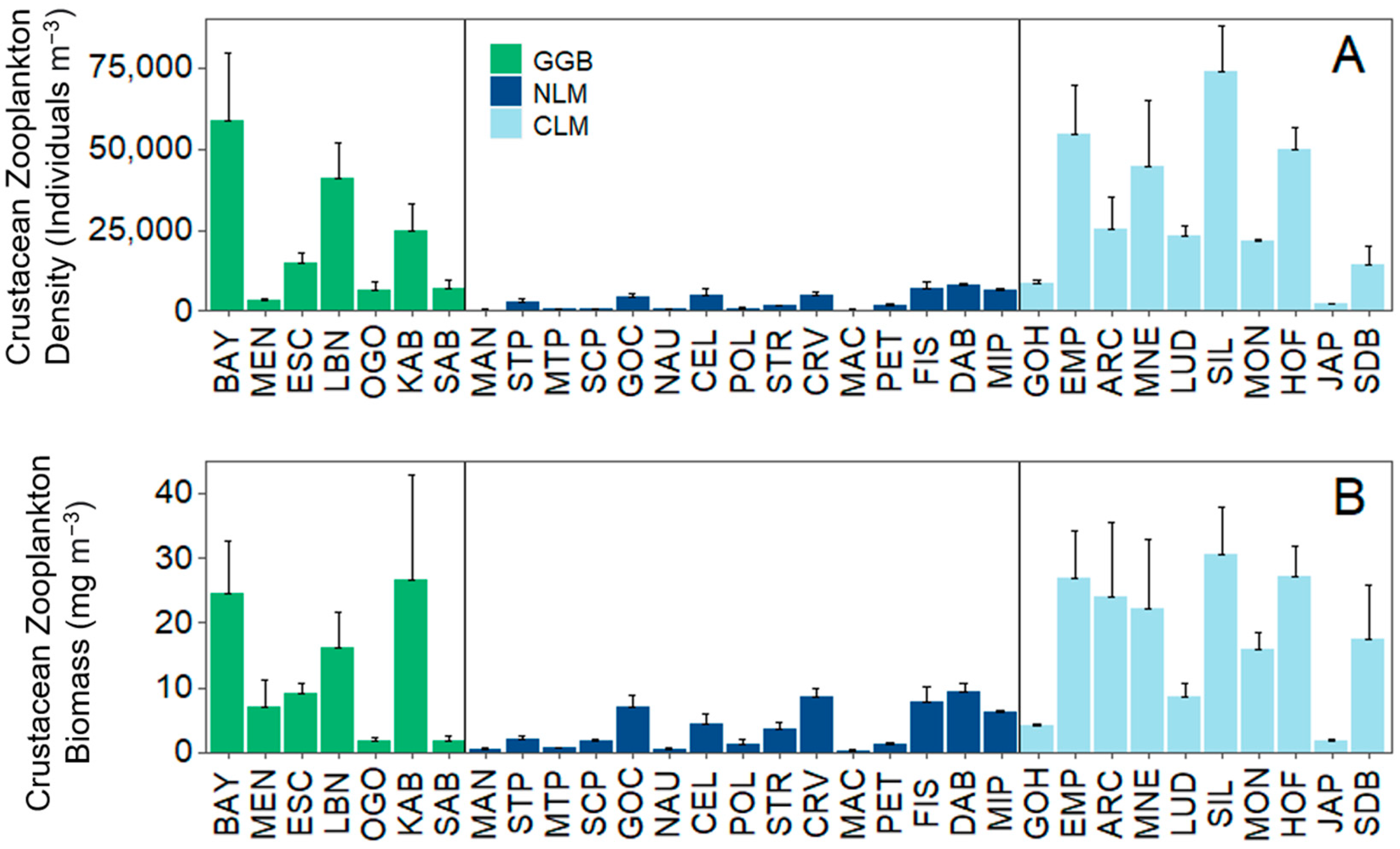
3.1. Analysis 1: Environmental Data and Total Crustacean Zooplankton Density and Biomass
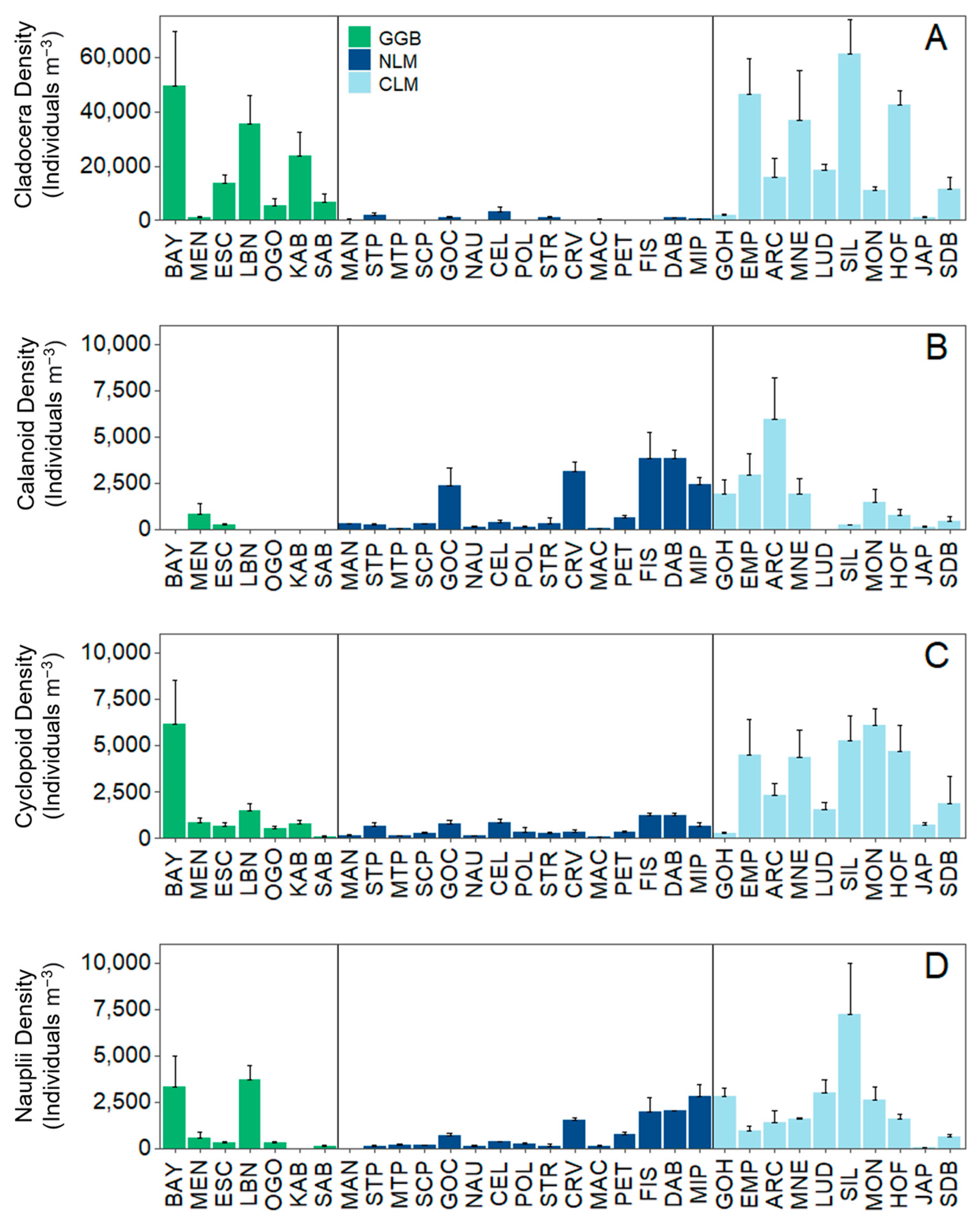
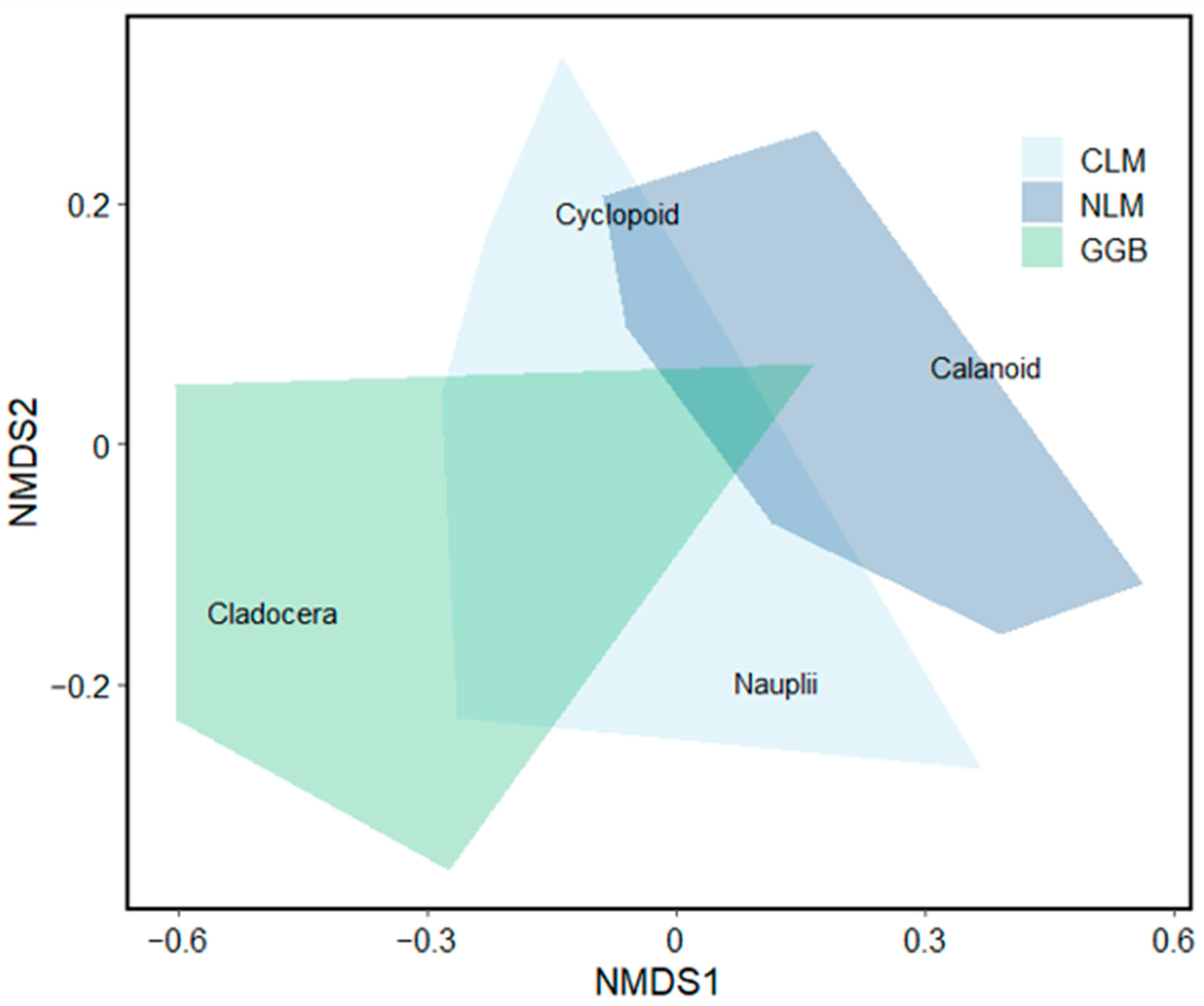
3.2. Analysis 2: Zooplankton Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbiero, R.P.; Lesht, B.M.; Warren, G.J. Convergence of trophic state and the lower food web in Lakes Huron, Michigan, and Superior. J. Great Lakes Res. 2012, 38, 368–380. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Pothoven, S.A.; Fahnenstiel, G.L.; Cavaletto, J.F.; Liebig, J.R.; Stow, C.A.; Nalepa, T.F.; Madenjian, C.P.; Bunnell, D.B. Seasonal zooplankton dynamics in Lake Michigan: Disentangling impacts of resource limitation, ecosystem engineering, and predation during a critical ecosystem transition. J. Great Lakes Res. 2012, 38, 336–352. [Google Scholar] [CrossRef]
- De Stasio, B.T.; Schrimpf, M.B.; Cornwell, B.H. Phytoplankton Communities in Green Bay, Lake Michigan after Invasion by Dreissenid Mussels: Increased Dominance by Cyanobacteria. Diversity 2014, 6, 681–704. [Google Scholar] [CrossRef]
- Knoll, L.B.; Sarnelle, O.; Hamilton, S.K.; Kissman, C.E.H.; Wilson, A.E.; Rose, J.B.; Morgan, M.R. Invasive zebra mussels (Dreissena polymorpha) increase cyanobacterial toxin concentrations in low-nutrient lakes. Can. J. Fish. Aquat. Sci. 2008, 65, 448–455. [Google Scholar] [CrossRef]
- Doubek, J.P.; Lehman, J.T. Historical biomass of Limnocalanus in Lake Michigan. J. Great Lakes Res. 2011, 37, 159–164. [Google Scholar] [CrossRef]
- Barbiero, R.P.; Bunnell, D.B.; Rockwell, D.C.; Tuchman, M.L. Recent increases in the large glacial-relict calanoid Limnocalanus macrurus in Lake Michigan. J. Great Lakes Res. 2009, 35, 285–292. [Google Scholar] [CrossRef]
- Barbiero, R.P.; Balcer, M.; Rockwell, D.C.; Tuchman, M.L. Recent shifts in the crustacean zooplankton community of Lake Huron. Can. J. Fish. Aquat. Sci. 2009, 66, 816–828. [Google Scholar] [CrossRef]
- Pothoven, S.A.; Madenjian, C.P. Changes in consumption by alewives and lake whitefish after dreissenid mussel invasions in Lakes Michigan and Huron. N. Am. J. Fish. Manag. 2008, 28, 308–320. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Nalepa, T.F.; Jude, D.J.; Mills, E.L.; Holeck, K.T.; Liebig, J.R.; Grigorovich, I.A.; Ojaveer, H. Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 1209–1228. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R.; Cochran, P.A.; Elser, J.J.; Elser, M.M.; Lodge, D.M.; Kretchmer, D.; He, X. Regulation of lake primary productivity by food web structure. Ecology 1987, 68, 1863–1876. [Google Scholar] [CrossRef]
- Rinke, K.; Huber, A.M.R.; Kempke, S.; Eder, M.; Wolf, T.; Probst, W.N.; Rothhaupta, K.-O. Lake-wide distributions of temperature, phytoplankton, zooplankton, and fish in the pelagic zone of a large lake. Limnol. Oceanogr. 2009, 54, 1306–1322. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Ruberg, S.A.; Höök, T.O.; Pothoven, S.A.; Brandt, S.B.; Lang, G.A.; Liebig, J.R.; Cavaletto, J.F. Hypoxia affects spatial distributions and overlap of pelagic fish, zooplankton, and phytoplankton in Lake Erie. J. Exp. Mar. Biol. Ecol. 2009, 381, S92–S107. [Google Scholar] [CrossRef]
- Lehman, J.T.; Bowers, J.A.; Gensemer, R.W.; Warren, G.J.; Branstrator, D.K. Mysis relicta in Lake Michigan: Abundances and relationships with their potential prey, Daphnia. Can. J. Fish. Aquat. Sci. 1990, 47, 977–983. [Google Scholar] [CrossRef]
- Pothoven, S.A.; Fahnenstiel, G.L. Spatial and temporal trends in zooplankton assemblages along a nearshore to offshore transect in southeastern Lake Michigan from 2007 to 2012. J. Great Lakes Res. 2015, 41, 95–103. [Google Scholar] [CrossRef]
- Doubek, J.P.; Goldfarb, S.K.; Stockwell, J.D. Should we be sampling zooplankton at night? Limnol. Oceanogr. Lett. 2020, 5, 313–321. [Google Scholar] [CrossRef]
- Pothoven, S.A. The influence of ontogeny and prey abundance on feeding ecology of age-0 lake whitefish (Coregonus clupeaformis) in southeastern Lake Michigan. Ecol. Freshw. Fish. 2020, 29, 103–111. [Google Scholar] [CrossRef]
- Reckahn, J.A. Ecology of young lake whitefish (Coregonus clupeaformis) in South Bay, Manitoulin Island, Lake Huron. In The Biology of Coregonid Fishes; Lindsay, C.C., Woods, C.S., Eds.; University of Manitoba Press: Winnipeg, MN, USA, 1970; pp. 437–459. [Google Scholar]
- Brooks, J.L.; Dodson, S.I. Predation, body size, and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Carter, J.L.; Schindler, D.E.; Francis, T.B. Effects of climate change on zooplankton community interactions in an Alaskan lake. Clim. Chang. Res. 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Carter, J.C.H.; Goudie, K.A. Diel vertical migrations and horizontal distributions of Limnocalanus macrurus and Senecella calanoides (Copepoda, Calanoida) in Lakes of Southern Ontario in relation to planktivorous fish. Can. J. Fish. Aquat. Sci. 1986, 43, 2508–2514. [Google Scholar] [CrossRef]
- Lougheed, V.L.; Chow-Fraser, P. Development and use of a zooplankton index of wetland quality in the Laurentian Great Lakes basin. Ecol. Appl. 2002, 12, 474–486. [Google Scholar] [CrossRef]
- Lougheed, V.L.; Crosbie, B.; Chow-Fraser, P. Predictions on the effect of common carp (Cyprinus carpio) exclusion on water quality, zooplankton, and submergent macrophytes in a Great Lakes wetland. Can. J. Fish. Aquat. Sci. 1998, 55, 1189–1197. [Google Scholar] [CrossRef]
- Fernandez, R.J.; Rennie, M.D.; Sprules, W.G. Changes in nearshore zooplankton associated with species invasions and potential effects on larval lake whitefish (Coregonus clupeaformis). Int. Rev. Hydrobiol. 2009, 94, 226–243. [Google Scholar] [CrossRef]
- Lucke, V.S.; Stewart, T.R.; Vinson, M.R.; Glase, J.D.; Stockwell, J.D. Larval Coregonus spp. diets and zooplankton community patterns in the Apostle Islands, Lake Superior. J. Great Lakes Res. 2020, 46, 1391–1401. [Google Scholar] [CrossRef]
- Rellstab, C.; Bürgi, H.R.; Müller, R. Population regulation in coregonids: The significance of zooplankton concentration for larval mortality. Ann. Zool. Fennici. 2004, 41, 281–290. [Google Scholar]
- Zorn, T.G.; Hayes, D.B.; McCullough, D.E.; Watson, N.M. Crustacean zooplankton available for larval walleyes in a Lake Michigan embayment. J. Great Lakes Res. 2020, 46, 1491–1499. [Google Scholar] [CrossRef]
- Brown, R.W.; Taylor, W.W. Effects of egg composition and prey density on the larval growth and survival of lake whitefish (Coregonus clupeaformis Mitchill). J. Fish Biol. 1992, 40, 381–394. [Google Scholar] [CrossRef]
- Taylor, W.W.; Freeberg, M.H. Effects of food abundance on larval lake whitefish, Coregonus clupeaformis Mitchill, growth and survival. J. Fish. Biol. 1984, 25, 733–741. [Google Scholar] [CrossRef]
- Keeler, K.M.; Bunnell, D.B.; Diana, J.S.; Adams, J.V.; Mychek-Londer, J.G.; Warner, D.M.; Yule, D.L.; Vinson, M.R. Evaluating the importance of abiotic and biotic drivers on Bythotrephes biomass in Lakes Superior and Michigan. J. Great Lakes Res. 2015, 41, 150–160. [Google Scholar] [CrossRef]
- Pawlowski, M.B.; Sierszen, M.E. A lake-wide approach for large lake zooplankton monitoring: Results from the 2006–2016 Lake Superior Cooperative Science and Monitoring Initiative surveys. J. Great Lakes Res. 2020, 46, 1015–1027. [Google Scholar] [CrossRef]
- Pothoven, S.A.; Höök, T.O.; Nalepa, T.F.; Thomas, M.V.; Dyble, J. Changes in zooplankton community structure associated with the disappearance of invasive alewife in Saginaw Bay, Lake Huron. Aquat. Ecol. 2013, 47, 1–12. [Google Scholar] [CrossRef]
- Beaver, J.R.; Tausz, C.E.; Renicker, T.R.; Holdren, G.C.; Hosler, D.M.; Manis, E.E.; Scotese, K.C.; Teacher, C.E.; Vitanye, B.T.; Davidson, R.M. The late summer crustacean zooplankton in western U.S.A. reservoirs reflects ecoregion, temperature and latitude. Freshw. Biol. 2014, 59, 1173–1186. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Dodson, S.I. Latitudinal patterns in the size distribution and seasonal dynamics of new world, freshwater cladocerans. Limnol Oceanogr. 2000, 45, 22–30. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Cavaletto, J.F.; Höök, T.O.; Pothoven, S.A.; Brandt, S.B.; Liebig, J.R.; Lang, G.A. Hypoxic zones as habitat for zooplankton in Lake Erie: Refuges from predation or exclusion zones? J. Exp. Mar. Biol. Ecol. 2009, 381, S108–S120. [Google Scholar] [CrossRef]
- Dodson, S.I.; Newman, A.L.; Will-Wolf, S.; Alexander, M.L.; Woodford, M.P.; Van Egeren, S. The relationship between zooplankton community structure and lake characteristics in temperate lakes (Northern Wisconsin, USA). J. Plankton Res. 2009, 31, 93–100. [Google Scholar] [CrossRef]
- Whitman, R.L.; Nevers, M.B.; Goodrich, M.L.; Murphy, P.C.; Davis, B.M. Characterization of Lake Michigan coastal lakes using zooplankton assemblages. Ecol. Indic. 2004, 4, 277–286. [Google Scholar] [CrossRef]
- Confer, J.L.; Kaaret, T.; Likens, G.E. Zooplankton diversity and biomass in recently acidified lakes. Can. J. Fish. Aquat. Sci. 1983, 40, 36–42. [Google Scholar] [CrossRef]
- Jeziorski, A.; Tanentzap, A.J.; Yan, N.D.; Paterson, A.M.; Palmer, M.E.; Korosi, J.B.; Rusak, J.A.; Arts, M.T.; Keller, W.; Ingram, R.; et al. The jellification of north temperate lakes. Proc. Roy. Soc. B. 2015, 282, 20142449. [Google Scholar] [CrossRef] [PubMed]
- Barbiero, R.P.; Rudstam, L.G.; Watkins, J.M.; Lesht, B.M. A cross-lake comparison of crustacean zooplankton communities in the Laurentian Great Lakes, 1997–2016. J. Great Lakes Res. 2019, 45, 672–690. [Google Scholar] [CrossRef]
- Bonina, S.M.C.; Codling, G.; Corcoran, M.B.; Guo, J.; Giesy, J.P.; Li, A.; Sturchio, N.C.; Rockne, K.J. Temporal and spatial differences in deposition of organic matter and black carbon in Lake Michigan sediments over the period 1850–2010. J. Great Lakes Res. 2018, 44, 705–715. [Google Scholar] [CrossRef]
- Rowe, M.D.; Obenour, D.R.; Nalepa, T.F.; Vanderploeg, H.A.; Yousef, F.; Kerfoot, W.C. Mapping the spatial distribution of the biomass and filter-feeding effect of invasive dreissenid mussels on the winter-spring phytoplankton bloom in Lake Michigan. Freshw. Biol. 2015, 60, 2270–2285. [Google Scholar] [CrossRef]
- Downing, J.A.; Rigler, F.H. Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters; Blackwell Scientific Publications: Oxford, UK, 1984. [Google Scholar]
- Balcer, M.; Korda, N.; Dodson, S.I. Zooplankton of the Great Lakes: A Guide to the Identification and Ecology of the Common Crustacean Species; The University of Wisconsin Press: Madison, WI, USA, 1984. [Google Scholar]
- Chick, J.H.; Levchuk, A.P.; Medley, K.A.; Havel, J.H. Underestimation of rotifer abundance a much greater problem than previously appreciated. Limnol. Oceanogr. 2010, 8, 79–87. [Google Scholar] [CrossRef]
- Doubek, J.P.; Campbell, K.L.; Doubek, K.M.; Hamre, K.D.; Lofton, M.E.; McClure, R.P.; Ward, N.K.; Carey, C.C. The effects of hypolimnetic anoxia on the diel vertical migration of freshwater crustacean zooplankton. Ecosphere 2018, 9, e02332. [Google Scholar] [CrossRef]
- Bottrell, H.H.; Duncan, A.; Gliwicz, M.Z.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. A review of some problems in zooplankton production studies. Norw. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Dumont, H.J.; Van De Velde, I.; Dumont, S. The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.M.; Rudstam, L.G.; Holeck, K.T. Length Weight Regressions for Zooplankton Biomass Calculations—A Review and a Suggestion for Standard Equations. eCommons Cornell. 2011. Available online: http://hdl.handle.net/1813/24566 (accessed on 9 September 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Burks, R.L.; Lodge, D.M.; Jeppesen, E.; Lauridsen, T.L. Diel horizontal migration of zooplankton: Costs and benefits of inhabiting the littoral. Freshw. Biol. 2002, 47, 343–365. [Google Scholar] [CrossRef]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar] [CrossRef]
- Haffner, G.D.; Yallop, M.L.; Herbert, P.D.N.; Griffiths, M. Ecological significance of upwelling events in Lake Ontario. J. Great Lakes Res. 1984, 10, 28–37. [Google Scholar] [CrossRef]
- Cumming, B.F.; Laird, K.R.; Gregory-Eaves, I.; Simpson, K.G.; Sokal, M.A.; Nordin, R.N.; Walker, I.R. Tracking past changes in lake-water phosphorus with a 251-lake calibration dataset in British Columbia: Tool development and application in a multiproxy assessment of eutrophication and recovery in Osoyoos Lake, a transboundary lake in Western North America. Front. Ecol. Evol. 2015, 3, 84. [Google Scholar] [CrossRef]
- Hatt, B.E.; Fletcher, T.D.; Walsh, C.J.; Taylor, S.L. The influence of urban density and drainage infrastructure on the concentrations and loads of pollutants in small streams. Environ. Manag. 2004, 34, 112–124. [Google Scholar] [CrossRef]
- Pace, M.L. An empirical analysis of zooplankton community size structure across lake trophic gradients. Limnol. Oceanogr. 1986, 31, 45–55. [Google Scholar] [CrossRef]
- Yuan, L.L.; Pollard, A.I. Changes in the relationship between zooplankton and phytoplankton biomasses across a eutrophication gradient. Limnol. Oceanogr. 2018, 63, 2493–2507. [Google Scholar] [CrossRef] [PubMed]
- Modeling Subcommittee; Technical Fisheries Committee. Technical Fisheries Committee Administrative Report 2020: Status of Lake Trout and Lake Whitefish Populations in the 1836 Treaty-Ceded Waters of Lakes Superior, Huron and Michigan, with Recommended Yield and Effort Levels for 2020. 2020. Available online: https://www.michigan.gov/greatlakesconsentdecree (accessed on 12 October 2023).
- Goodyear, C.S.; Edsall, T.A.; Dempsey, D.M.O.; Moss, G.D.; Polanski, P.E. Atlas of the Spawning and Nursery Areas of Great Lakes Fishes. Reproductive characteristics of Great Lakes Fishes. U.S. Fish and Wildlife Service, 1982. FWS/OBS-82/52. Available online: https://archive.epa.gov/ecopage/web/pdf/atlas-spawning-nursery-areas-great-lakes-fishes-volume-2-lake-superior-198209-124pp.pdf (accessed on 12 October 2023).
- Gobin, J.; Lester, N.P.; Cottrill, A.; Fox, M.G.; Dunlop, E.S. Trends in growth and recruitment of Lake Huron lake whitefish during a period of ecosystem change, 1985 to 2012. J. Great Lakes Ries. 2015, 41, 405–414. [Google Scholar] [CrossRef]
- Evans, M.S.; Sell, D.W. Mesh size and collection characteristics of a 50-cm diameter conical plankton nets. Hydrobiologia 1977, 122, 97–104. [Google Scholar] [CrossRef]
- Mantzouki, E.; Beklioǧlu, M.; Brookes, J.D.; de Senerpont Domis, L.N.; Dugan, H.A.; Doubek, J.P.; Grossart, H.-P.; Nejstgaard, J.C.; Pollard, A.I.; Ptacnik, R.; et al. Snapshot surveys for lake monitoring, more than a shot in the dark. Front. Ecol. Evol. 2018, 6, 201. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean ± 1 S.D. | Median | Minimum | Maximum |
|---|---|---|---|---|
| Greater Green Bay | ||||
| Water temperature, °C | 21.3 ± 1.9 | 21.7 | 18.2 | 23.4 |
| Dissolved oxygen, mg L−1 | 8.4 ± 1.3 | 9.0 | 6.0 | 10.2 |
| Specific conductivity, µS cm−1 | 270 ± 17 | 280 | 250 | 310 |
| pH | 8.1 ± 0.3 | 8.2 | 7.7 | 8.5 |
| Northern Lake Michigan | ||||
| Water temperature, °C | 16.7 ± 2.5 | 16.2 | 11.9 | 21.1 |
| Dissolved oxygen, mg L−1 | 9.8 ± 0.8 | 9.7 | 8.9 | 11.6 |
| Specific conductivity, µS cm−1 | 280 ± 21 | 270 | 250 | 310 |
| pH | 8.6 ± 0.3 | 8.5 | 8.3 | 9.0 |
| Central Lake Michigan | ||||
| Water temperature, °C | 20.7 ± 1.7 | 20.8 | 18.1 | 23.1 |
| Dissolved oxygen, mg L−1 | 9.9 ± 0.6 | 10.0 | 9.0 | 10.8 |
| Specific conductivity, µS cm−1 | 310 ± 16.0 | 310 | 285 | 340 |
| pH | 8.9 ± 0.3 | 8.9 | 8.3 | 9.1 |
| Group | Mean ± 1 S.D. | Median | Minimum | Maximum |
|---|---|---|---|---|
| Density (ind. m−3) | ||||
| Total crustacean | 16,398 ± 20,226 | 6707 | 341 | 74,010 |
| Cladocera | 12,302 ± 17,669 | 1965 | 78.6 | 61,435 |
| Cyclopoid | 1563 ± 1877 | 773 | 78.6 | 6170 |
| Calanoid | 1102 ± 1473 | 364 | 0 | 5973 |
| Nauplii | 1296 ± 1558 | 655 | 0 | 7205 |
| Biomass (mg DW m−3) | ||||
| Total crustacean | 10.1 ± 9.8 | 7.2 | 0.2 | 30.6 |
| Cladocera | 5.2 ± 7.0 | 1.7 | 0.06 | 26.2 |
| Cyclopoid | 2.3 ± 2.6 | 1.1 | 0.07 | 9.2 |
| Calanoid | 2.7 ± 3.4 | 1.2 | 0 | 15.8 |
| Nauplii | 0.4 ± 0.3 | 0.3 | 0 | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnston, S.J.; Smith, J.B.; Slater, B.D.; Doubek, J.P. Spatial Density, Biomass, and Composition of Crustacean Zooplankton on Lake Michigan Beaches. Fishes 2023, 8, 599. https://doi.org/10.3390/fishes8120599
Johnston SJ, Smith JB, Slater BD, Doubek JP. Spatial Density, Biomass, and Composition of Crustacean Zooplankton on Lake Michigan Beaches. Fishes. 2023; 8(12):599. https://doi.org/10.3390/fishes8120599
Chicago/Turabian StyleJohnston, Samuel J., Jason B. Smith, Brady D. Slater, and Jonathan P. Doubek. 2023. "Spatial Density, Biomass, and Composition of Crustacean Zooplankton on Lake Michigan Beaches" Fishes 8, no. 12: 599. https://doi.org/10.3390/fishes8120599
APA StyleJohnston, S. J., Smith, J. B., Slater, B. D., & Doubek, J. P. (2023). Spatial Density, Biomass, and Composition of Crustacean Zooplankton on Lake Michigan Beaches. Fishes, 8(12), 599. https://doi.org/10.3390/fishes8120599