

Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Age Determination
2.3. Sexual Maturity
2.4. Stomach and Intestine Content Analysis
2.4.1. Morphological Identification of Stomach and Intestine Contents
2.4.2. Molecular Identification of Stomach and Intestine Contents
3. Results
3.1. Size and Age
3.2. Feeding Habits
3.2.1. Morphological Identification of Stomach and Intestine Contents
3.2.2. Molecular Analysis of Stomach and Intestine Contents
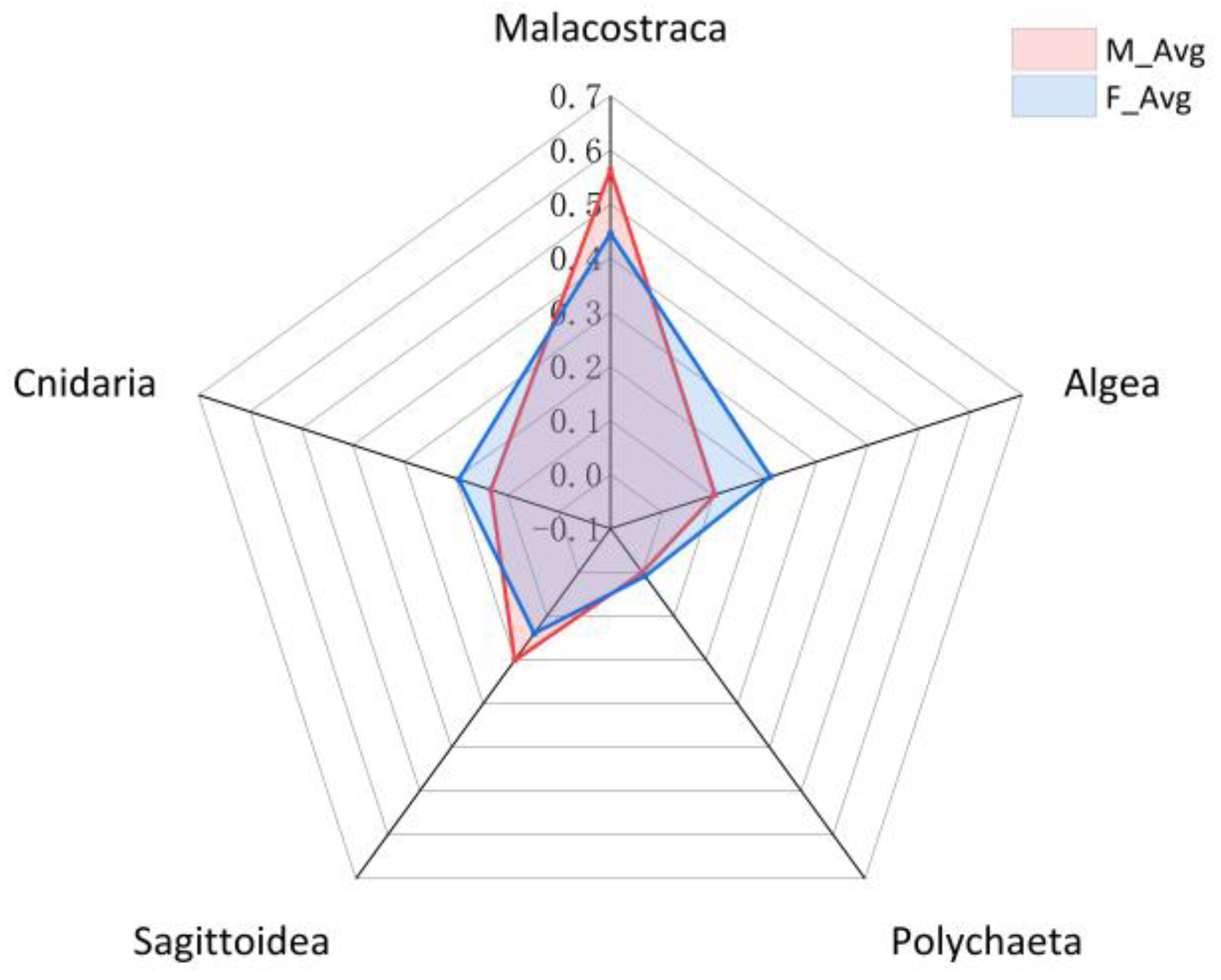
3.3. Feeding Habits Change with Gender
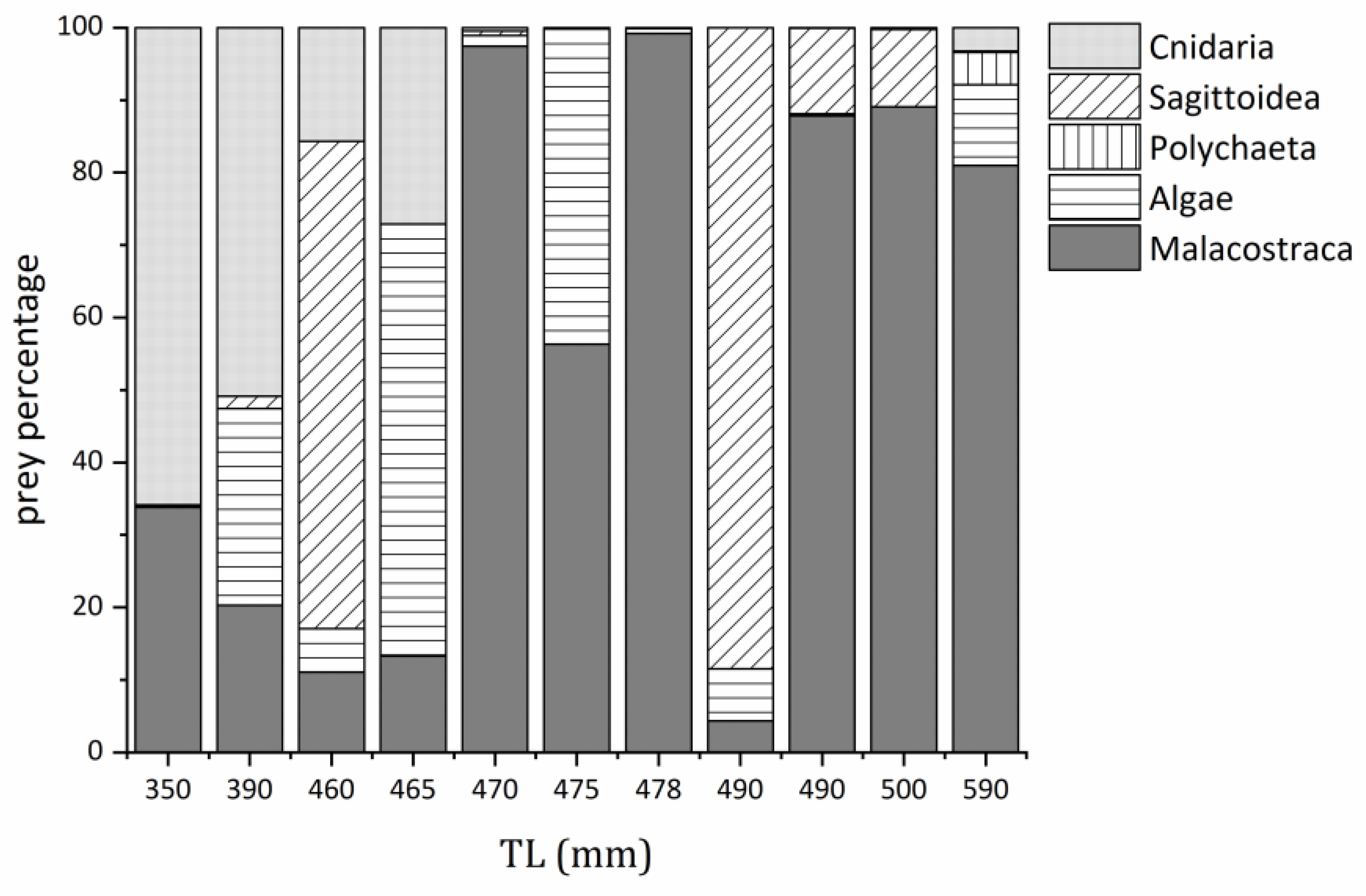
3.4. Prey Composition Changes with a Total Length
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, P.J.; Steinke, D.; McMillan, P.J.; Stewart, A.L.; McVeagh, S.M.; De Astarloa, J.M.D.; Welsford, D.; Ward, R.D. DNA barcoding highlights a cryptic species of grenadier Macrourus in the Southern Ocean. J. Fish Biol. 2010, 78, 355–365. [Google Scholar] [CrossRef] [PubMed]
- McMillan, P.; Iwamoto, T.; Stewart, A.; Smith, P.J. A new species of grenadier, genus Macrourus (Teleostei, Gadiformes, Macrouridae) from the southern hemisphere and a revision of the genus. Zootaxa 2012, 3165, 19. [Google Scholar] [CrossRef]
- McMillan, P.J.; Marriott, P.; Hanchet, S.M.; Fenaughty, J.M.; Mackay, E.; Sui, H.; Wei, F. Fishes of the Ross Sea region. A Field Guide to Common Species Caught in the Longline Fishery; New Zealand Aquatic Environment and Biodiversity Report, No. 134; Ministry for Primary Industries (MPI): Wellington, New Zealand, 2014; 54p.
- Daniel, M.C.; Tadashi, I.; Tomio, I.; Nadia, S. FAO Species Catalogue Vol.10 Gadiform Fishes of the World; FAO: Rome, Italy, 1990; Volume 10, pp. 238–240. [Google Scholar]
- Fenaughty, J.M.; Stevens, D.W.; Hanchet, S.M. Diet of the Antarctic toothfish (Dissostichus mawsoni) from the Ross Sea, Antarctica (CCAMLR Statistical Subarea 88.1). CCAMLR Sci. 2003, 10, 113–123. [Google Scholar]
- Stevens, D.W.; Dunn, M.R.; Pinkerton, M.H.; Forman, J.S. Diet of Antarctic Toothfish (Dissostichus mawsoni) from the Western Ross Sea, Antarctica; Document WG-FSA-12/52; CCAMLR: Hobart, Australia, 2012. [Google Scholar]
- Pinkerton, M.; McMillan, P.; Forman, J.; Marriott, P.; Horn, P.; Bury, S.; Brown, J. Distribution, morphology and ecology of Macrourus whitsoni and M. caml (Gadiformes, Macrouridae) in the Ross Sea region. CCAMLR Sci. 2013, 20, 37–61. [Google Scholar]
- Marriott, P.M.; Manning, M.J.; Horn, P.L. Age estimation and maturity of the ridge-scaled macrourid (Macrourus whitsoni) from the Ross Sea. CCAMLR Sci. 2006, 13, 291–303. [Google Scholar]
- Moore, B.R.; Parker, S.J.; Pinkerton, M.H. Otolith shape as a tool for species identification of the grenadiers Macrourus caml and M. whitsoni. Fish. Res. 2022, 253, 106370. [Google Scholar] [CrossRef]
- Van Beveren, E.; Bonhommeau, S.; Fromentin, J.-M.; Bigot, J.-L.; Bourdeix, J.-H.; Brosset, P.; Roos, D.; Saraux, C. Rapid changes in growth, condition, size and age of small pelagic fish in the Mediterranean. Mar. Biol. 2014, 161, 1809–1822. [Google Scholar] [CrossRef]
- Ju, P.; Cheung, W.W.L.; Lu, Z.; Yang, S.; Guo, Z.; Chen, M.; Xiao, J. Age, growth, and abundance fluctuation of Jordan’s damsel, Teixeirichthys jordani (Actinopterygii: Perciformes: Pomacentridae), in the southern Taiwan Strait. Acta Ichthyol. Et Piscat. 2019, 49, 243–250. [Google Scholar] [CrossRef]
- Liao, C.S.; Li, W.; Yuan, J.; Ye, S.W.; Zhang, T.L.; Liu, J.S. Age, growth, and motality characteristics of Pelteobagrus Nitidus (sauvage et dabry) in the three gorges reservoir. Acta Hydrobiol. Sin. 2022, 46, 29–36. (In Chinese) [Google Scholar] [CrossRef]
- Marriott, P.; Horn, P.L.; McMillan, P. Species identification and age estimation for the ridge-scaled Macrourid (Macrourus whitsoni) from the Ross Sea. CCAMLR Sci. 2003, 10, 37–51. [Google Scholar]
- Xie, X.; Bao, Z.Y.; Wang, Q.Z. Advances on research and application of age determination by hard tissues in fish: A review. J. Dalian Ocean Univ. 2021, 36, 1071–1080. (In Chinese) [Google Scholar]
- Jalbani, S.; Nareio, N.T.; Jalbani, Y.M.; Lashari, I.A. Age determination of minor carp, cirrhinus reba (hamilton) from manchar lake, sindh, pakistan. Stand. Sci. Res. Essays 2014, 2, 577–580. [Google Scholar]
- Barnett, B.K.; Chanton, J.P.; Ahrens, R.; Thornton, L.; Iii, W.F.P. Life history of northern Gulf of Mexico Warsaw grouper Hyporthodus nigritus inferred from otolith radiocarbon analysis. PLoS ONE 2020, 15, e0228254. [Google Scholar] [CrossRef]
- Sanchez, P.J.; Pinsky, J.P.; Rooker, J.R. Bomb Radiocarbon Age Validation of Warsaw Grouper and Snowy Grouper. Fisheries 2019, 44, 524–533. [Google Scholar] [CrossRef]
- Cassia, M.C. Comparison of age readings from scales and otoliths of the Patagonian toothfish (Dissostichus Eleginoides) from South Georgia. CCAMLR Sci. 1998, 5, 191–203. [Google Scholar]
- Brickle, P.; Arkhipkin, A.I.; Shcherbich, Z.N. Age and growth in a temperate euryhaline notothenioid, eleginops maclovinus from the falkland islands. J. Mar. Biol. Assoc. U. K. 2005, 85, 1217–1221. [Google Scholar] [CrossRef]
- Zhu, G.P.; Wei, L. Age and growth of Antarctic fish species: A review. J. Fish. China 2017, 41, 1638–1647. (In Chinese) [Google Scholar]
- Xue, Y.; Jin, X.S.; Zhang, B.; Liang, Z.L. Diet composition and seasonal variation in feeding habits of small yellow croaker Pseudosciaena polyactis Bleeker in the central Yellow Sea. J. Fish. Sci. China 2004, 11, 237–243. (In Chinese) [Google Scholar]
- Wang, J.; Jiang, R.-J.; Hu, C.-L.; Li, Z.; Xiao, Y.; Xu, Y.-J.; He, Z.-T.; Xu, H.-X. Feeding ecology of Engraulis japonicus based on stomach contents and stable isotope. Yingyong Shengtai Xuebao 2021, 32, 2035–2044. (In Chinese) [Google Scholar] [CrossRef]
- Hynes, H.B.N. The Food of Fresh-Water Sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius), with a review of methods used in studies of the food of fishes. J. Anim. Ecol. 1950, 19, 36–58. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Scacco, U.; Tiralongo, F.; Mancini, E. Feeding in Deep Waters: Temporal and Size-Related Plasticity in the Diet of the Slope Predator Fish Coelorinchus caelorhincus (Risso, 1810) in the Central Tyrrhenian Sea. J. Mar. Sci. Eng. 2022, 10, 1235. [Google Scholar] [CrossRef]
- Scacco, U.; Mancini, E.; Marcucci, F.; Tiralongo, F. Microplastics in the Deep: Comparing Dietary and Plastic Ingestion Data between Two Mediterranean Bathyal Opportunistic Feeder Species, Galeus melastomus, Rafinesque, 1810 and Coelorinchus caelorhincus (Risso, 1810), through Stomach Content Analysis. J. Mar. Sci. Eng. 2022, 10, 624. [Google Scholar] [CrossRef]
- Saraswati, E.; Perdhana, G.O. Analysis of stomach content of Nemipterus japonicus from the Blimbingsari waters, Banyuwangi, East Java. IOP Conf. Ser. Earth Environ. Sci. 2020, 404, 012019. [Google Scholar] [CrossRef]
- Hashimoto, R. Investigation of feeding habits and variation of inhabiting depths with cod (Gadus macrocephalus) distributed on the northeastern fishing ground in Japan. Bull. Tohoku Reg. Fish. Res. Lab. 1974, 33, 51–67. [Google Scholar]
- Mata-Sotres, J.A.; Moyano, F.J.; Martínez-Rodríguez, G.; Yúfera, M. Daily rhythms of digestive enzyme activity and gene expression in gilthead seabream (Sparus aurata) during ontogeny. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 197, 43–51. [Google Scholar] [CrossRef]
- Xi, X.Q.; Bao, B.L.; Zhang, S.Y. Application of DNA barcoding in species analysis of fish stomach content. J. Shanghai Ocean Univ. 2015, 24, 203–210. (In Chinese) [Google Scholar]
- Carreon-Martinez, L.; Johnson, T.B.; Ludsin, S.A.; Heath, D.D. Utilization of stomach content DNA to determine diet diversity in piscivorous fishes. J. Fish Biol. 2011, 78, 1170–1182. [Google Scholar] [CrossRef]
- Paquin, M.M.; Buckley, T.W.; Hibpshman, R.E.; Canino, M.F. DNA-based identification methods of prey fish from stomach contents of 12 species of eastern North Pacific groundfish. Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2013, 85, 110–117. [Google Scholar] [CrossRef]
- Symondson, W.O.C. Molecular identification of prey in predator diets. Mol. Ecol. 2002, 11, 627–641. [Google Scholar] [CrossRef]
- Leray, M.; Meyer, C.; Mills, S. Metabarcoding dietary analysis of coral dwelling predatory fish demonstrates the minor contribution of coral mutualists to their highly partitioned, generalist diet. Peer J. 2015, 3, e1047. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.P.; Ye, Z.J.; Xue, Y.; Zhang, C.; Xu, B.D.; Zhang, C.L.; Wang, J.; Mu, X.X. Fishery Resource Biology; China Agriculture Press: Beijing, China, 2020. [Google Scholar]
- Gordon, J.D.M.; Swan, S.C. Validation of age readings from otoliths of juvenile roundnose grenadier, Coryphaenoides rupestris, a deep-water macrourid fish. J. Fish Biol. 1996, 49, 289–297. [Google Scholar] [CrossRef]
- Pinkerton, M.H.; Forman, J.; Stevens, D.W.; Bury, S.J.; Brown, J. Diet and trophic niche of Macrourus spp. (Gadiformes, Macrouridae) in the Ross Sea region of the Southern Ocean. J. Ichthyol. 2012, 52, 787–799. [Google Scholar] [CrossRef]
- Pakhomov, E. Feeding and exploitation of the food supply by demersal fishes in the Antarctic part of the Indian Ocean. J. Ichthyol. 1997, 37, 360–380. [Google Scholar]
- Guo, B.; Zhang, B.; Jin, X.S. Diet composition and ontogenetic variation in feeding habits of juvenile small yellow croaker Pseudosciaena polyactis Bleeker in the Yellow Sea. J. Fish. Sci. China 2010, 17, 289–297. [Google Scholar]
- Harms-Tuohy, C.; Schizas, N.; Appeldoorn, R. Use of DNA metabarcoding for stomach content analysis in the invasive lionfish Pterois volitans in Puerto Rico. Mar. Ecol. Prog. Ser. 2016, 558, 181–191. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequences (5′–3′) |
|---|---|
| 18SV8V9F | ATAACAGGTCTGTGATGCCCT |
| 18SV8V9R | CCTTCYGCAGGTTCACCTAC |
| Reagent Name | Volume/μL |
|---|---|
| Q5 Hot Start High-Fidelity 2X Master Mix | 25 |
| Forward primer | 1 |
| Reverse primer | 1 |
| DNA | 1 |
| Nuclease free water | 22 |
| Samples | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gender | M | F | M | F | M | F | F | M | M | F | M |
| Age | 13 | 13 | 10 | - | 9 | 15 | 12 | 12 | 10 | 19 | 10 |
| Maturity | 3 | 2 | 2 | 3 | 2 | 4 | 4 | 3 | 3 | 5 | 2 |
| Weight/g | 450 | 480 | 270 | 630 | 220 | 620 | 670 | 500 | 520 | 900 | 470 |
| Standard Length/mm | 455 | 485 | 385 | 490 | 348 | 485 | 470 | 460 | 473 | 585 | 460 |
| Total length/mm | 460 | 490 | 390 | 500 | 350 | 490 | 475 | 465 | 478 | 590 | 470 |
| Anal length/mm | 150 | 160 | 140 | 180 | 125 | 175 | 175 | 165 | 170 | 190 | 160 |
| Prey Types | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Malacostraca | + | + | + | + | + | + | + | + | + | + | + |
| Algae | + | + | + | ||||||||
| Cephalopoda | + | + | |||||||||
| Fish | + | + | |||||||||
| Polychaeta | + | + | + | + | + | + | + | ||||
| Other benthos | + |
| Prey Types | Percentage of Contigs | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | Total | ||
| Malacostraca | All Malacostraca | 11.064 | 4.346 | 20.280 | 89.085 | 33.835 | 87.840 | 56.332 | 13.277 | 99.186 | 80.977 | 97.455 | 57.820 |
| Peracarida | 0.008 | 0 | 4.001 | 0 | 0.022 | 0 | 0.074 | 0 | 1.002 | 0.017 | 1.620 | 0.544 | |
| Eucarida | 11.056 | 4.346 | 16.278 | 89.085 | 33.813 | 87.840 | 56.259 | 13.277 | 98.184 | 80.961 | 95.660 | 57.255 | |
| Malacostraca (unidentified) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.175 | 0.021 | |
| Algae | All Algae | 5.977 | 7.201 | 27.158 | 0 | 0.044 | 0.293 | 43.471 | 59.588 | 0.713 | 11.155 | 1.443 | 5.666 |
| Thalassiosira spp. | 0 | 0.008 | 12.428 | 0 | 0 | 0.101 | 0.000 | 21.254 | 0 | 3.794 | 0.101 | 0.643 | |
| Chaetoceros spp. | 0 | 0 | 0 | 0 | 0 | 0.077 | 0.000 | 0 | 0 | 0.142 | 0 | 0.012 | |
| Actinocyclus curvatulus | 0 | 0 | 0 | 0 | 0 | 0 | 7.587 | 0 | 0 | 0.000 | 0 | 0.238 | |
| Navicula spp. | 5.976 | 0 | 0 | 0 | 0.024 | 0 | 0.301 | 0 | 0 | 2.980 | 0 | 1.158 | |
| Phaeocystis spp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.176 | 0 | 0 | 0 | 0.002 | |
| Algae (unidentified) | 0.001 | 7.194 | 14.730 | 0.000 | 0.019 | 0.115 | 35.584 | 38.158 | 0.713 | 4.239 | 1.342 | 3.613 | |
| Polychaeta | Rhynchospio spp. | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 4.469 | 0.000 | 0.255 |
| Sagittoidea | All Phragmophora | 67.263 | 88.435 | 1.672 | 10.681 | 0.254 | 11.790 | 0.000 | 0.000 | 0.023 | 0.182 | 0.638 | 27.165 |
| Eukrohnia fowleri | 0.025 | 33.613 | 0.534 | 0.364 | 0.000 | 11.508 | 0.000 | 0.000 | 0.000 | 0.182 | 0.000 | 6.481 | |
| Phragmophora (unidentified) | 67.238 | 54.822 | 1.137 | 10.317 | 0.254 | 0.282 | 0.000 | 0.000 | 0.023 | 0.000 | 0.638 | 20.684 | |
| Cnidaria | All Cnidaria | 15.697 | 0.018 | 50.891 | 0.234 | 65.867 | 0.077 | 0.196 | 27.135 | 0.078 | 3.217 | 0.464 | 9.094 |
| Coronatae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.007 | 0 | 0 | |
| Siphonophorae | 12.381 | 0.010 | 50.891 | 0.234 | 0 | 0 | 0.092 | 27.029 | 0.003 | 0.000 | 0.464 | 3.096 | |
| Cnidaria (unidentified) | 3.316 | 0.008 | 0 | 0 | 65.867 | 0.077 | 0.104 | 0.106 | 0.075 | 3.210 | 0.000 | 5.997 | |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Sun, P.; Zhang, C.; Li, J.; Xi, X.; Ma, S.; Zhang, W.; Tian, Y. Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea. Fishes 2023, 8, 56. https://doi.org/10.3390/fishes8010056
Xu S, Sun P, Zhang C, Li J, Xi X, Ma S, Zhang W, Tian Y. Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea. Fishes. 2023; 8(1):56. https://doi.org/10.3390/fishes8010056
Chicago/Turabian StyleXu, Siqing, Peng Sun, Chi Zhang, Jianchao Li, Xiaoyu Xi, Shuyang Ma, Wenchao Zhang, and Yongjun Tian. 2023. "Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea" Fishes 8, no. 1: 56. https://doi.org/10.3390/fishes8010056
APA StyleXu, S., Sun, P., Zhang, C., Li, J., Xi, X., Ma, S., Zhang, W., & Tian, Y. (2023). Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea. Fishes, 8(1), 56. https://doi.org/10.3390/fishes8010056