
Genetic Diversity and Population Structure of Shoshone Sculpin Cottus greenei in the Hagerman Valley of South-Central Idaho

Abstract
1. Introduction
2. Methods
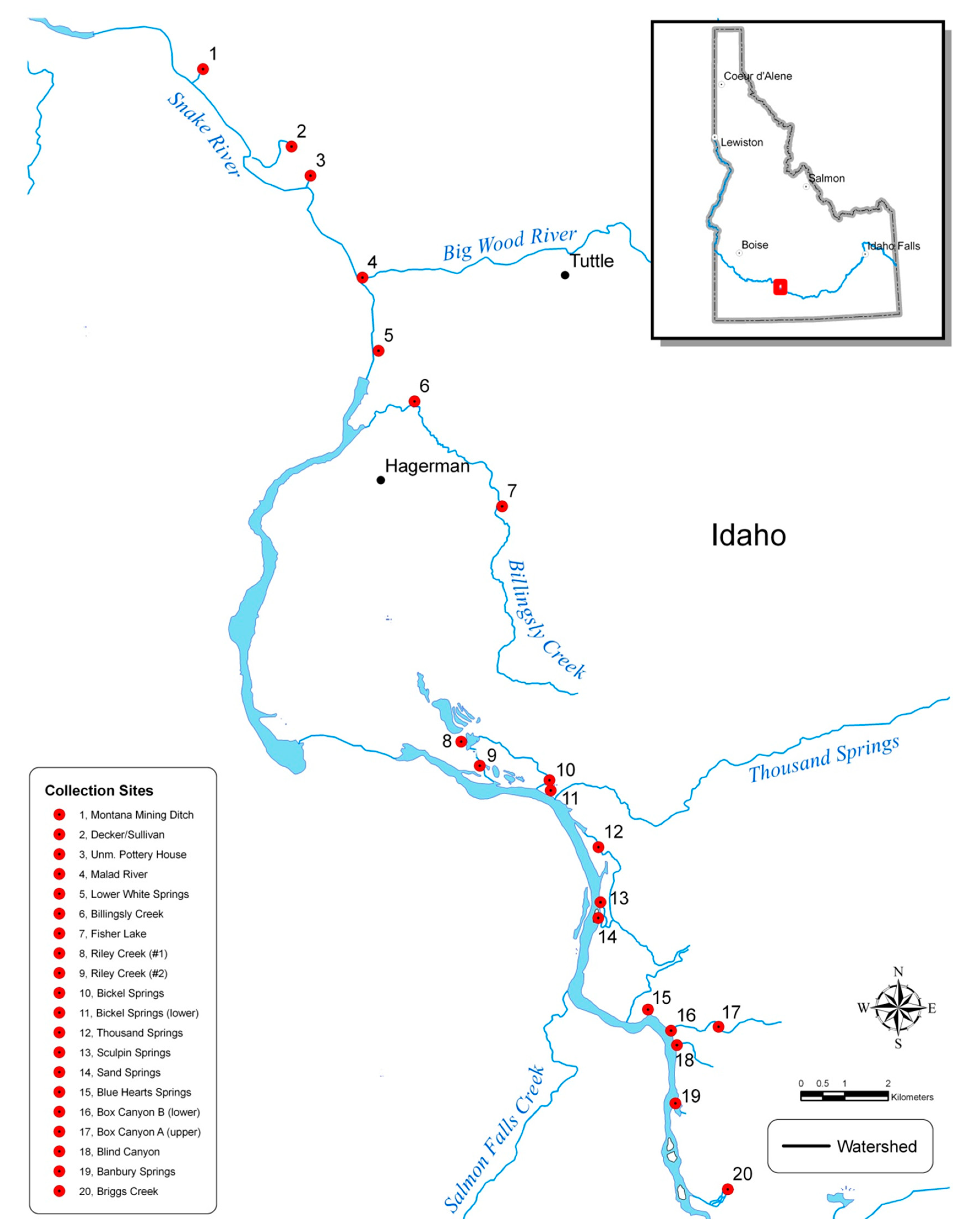
2.1. Population Sampling
2.2. DNA Extraction and Microsatellite DNA Optimization and Screening
2.3. Statistical Analyses
3. Results
3.1. Tests for Hardy-Weinberg Equilibrium and Linkage Disequilibrium
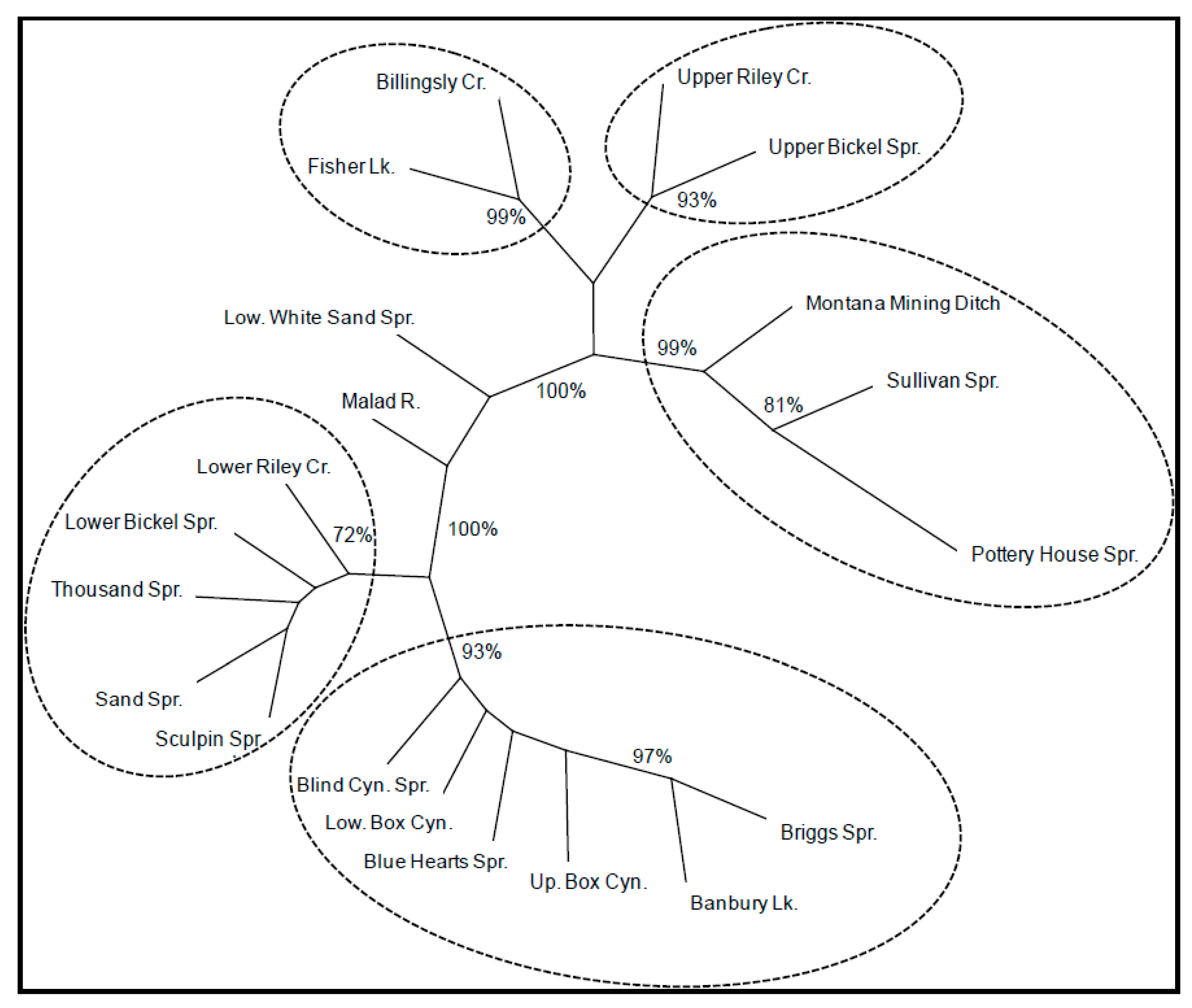
3.2. Genetic Differentiation and Structure
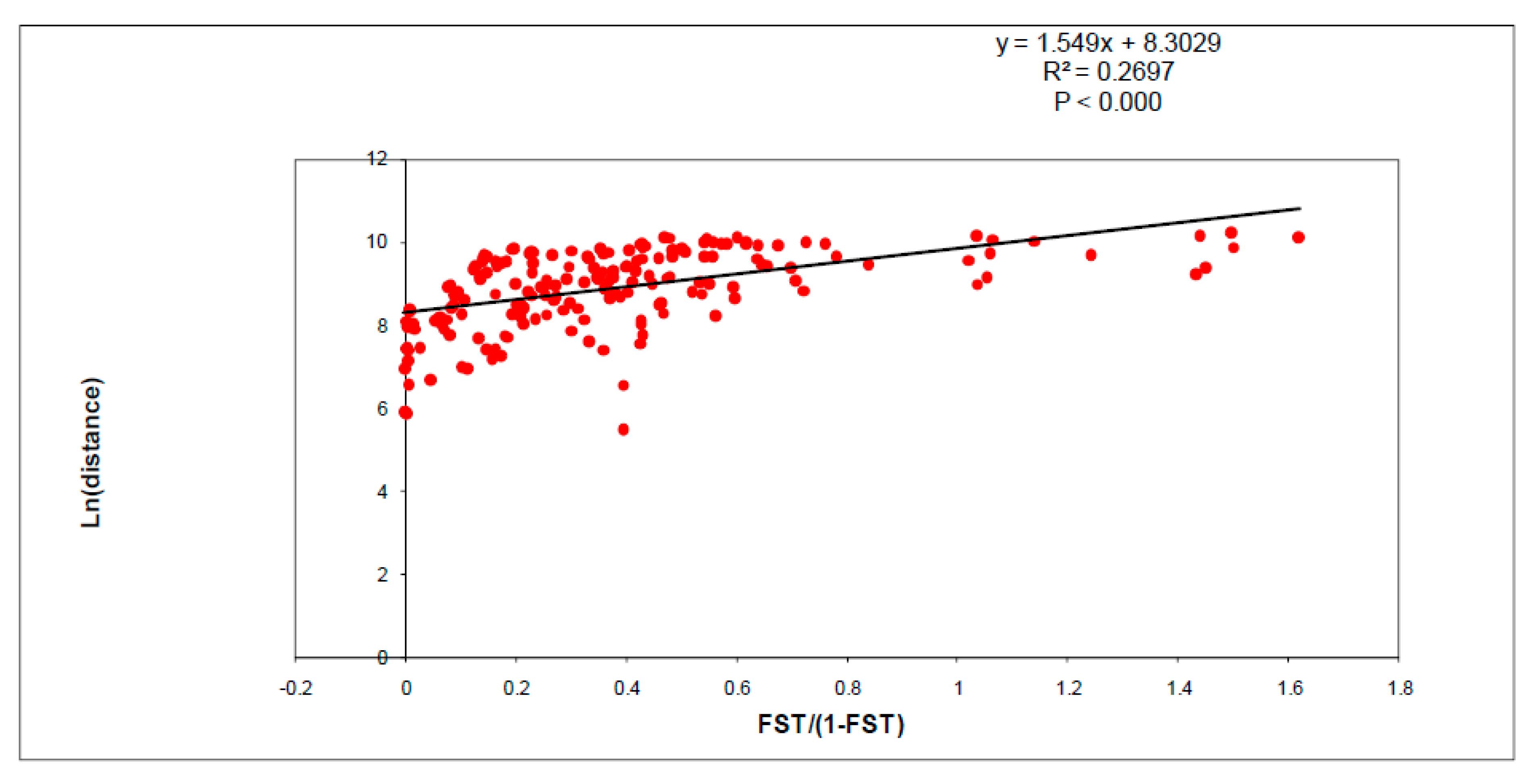
3.3. Isolation by Distance, Effective Population Size, and Bottlenecks
4. Discussion
5. Conclusions
6. Availability of Data and Material
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stearns, H. Origin of the Large Springs and their Alcoves along the Snake River in Southern Idaho. J. Geol. 1936, 44, 429–450. [Google Scholar] [CrossRef]
- IWRB (Idaho Water Resources Board). Comprehensive State Water Plan. Henrys Fork Basin, Idaho Water Resources Board; Rydalch, F.D., Parr, C., Gray, G.M., Bell, B.J., Hungerford, K.E., Kramer, D.R., Platts, W., Satterwhite, M., Eds.; 1992; Comprehensive State Water Plan: Henrys Fork Basin | December 1992. Available online: http://idaho.gov (accessed on 14 January 2023).
- U.S. Fish and Wildlife Service (USFWS). Endangered and threatened wildlife and plants: Determinations of endangered or threatened status for five aquatic snails in South Central Idaho. Fed. Regist. 1992, 57, 59244–59256. [Google Scholar]
- Wallace, R.L.; Griffith, J.S.; Daley, D.M.; Connolly, P.J.; Beckham, G.B. Distribution of the Shoshone sculpin (Cottus greenei: Cottidae) in the Hagerman Valley of South Central Idaho. Great Basin Nat. 1984, 44, 324–326. [Google Scholar]
- Young, M.K.; Smith, R.; Pilgrim, K.L.; Isaak, D.J.; McKelvey, K.S.; Parkes, S.; Egge, J.; Schwartz, M.K. A molecular taxonomy of Cottus in western North America. West. North Am. Nat. 2022, 82, 307–345. [Google Scholar] [CrossRef]
- Connolly, P.J. Life History of Shoshone Sculpin, Cottus greenei, in Southcentral Idaho. Unpublished. Master Thesis, University of Idaho, Moscow, Russia, 1983; 79p. [Google Scholar]
- Griifth, J.; Daley, D.M. Re-Establishment of Shoshone sculpin (Cottus greenei) in the Hagerman Valley, Idaho; Idaho Department of Fish and Game. Final Report; Nongame Program: Boise, ID, USA, 1984; 12p. [Google Scholar]
- NatureServe. NatureServe Explorer. Arlington (VA): NatureServe. Available online: https://explorer.natureserve.org. (accessed on 9 January 2023).
- Idaho Department of Fish and Game. Forthcoming. DRAFT Idaho State Wildlife Action Plan. 2023 rev. Eds. Boise (ID): Idaho Department of Fish and Game. Available online: https://idfg.idaho.gov/swap (accessed on 14 January 2023).
- Junker, J.; Peter, A.; Wagner, C.E.; Mwaiko, S.; Germann, B.; Seehausen, O.; Keller, I. River fragmentation increases localized population genetic structure and enhances asymmetry of dispersal in bullhead (Cottus gobio). Conserv. Genet 2012, 13, 545–556. [Google Scholar] [CrossRef]
- Lamphere, B.A.; Blum, M.J. Genetic estimates of population structure and dispersal in a benthic stream fish. Ecol. Freshw. Fish 2012, 21, 75–86. [Google Scholar] [CrossRef]
- Hudy, M.; Shiflet, J. Movement and recolonization of Potomac sculpin in a Virginia stream. North Am. J. Fish. Manag. 2009, 29, 196–204. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). Snake River Aquatic Species Recovery Plan; Snake River Basin Office, Ecological Services: Boise, ID, USA, 1995; 92p.
- AFS. America Fisheries Society. Use of Fishes in Research Committee (Joint Committee of the American Fisheries Society, the American Institute of Fishery Research Biologists, and the American Society of Ichthyologists and Herpetologists). In Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MD, USA, 2014. [Google Scholar]
- Fiumera, A.C.; Porter, B.A.; Grossman, G.D.; Avise, J.C. Intensive genetic assessment of the mating system and reproductive success in a semiclosed population of the mottled sculpin, Cottus bairdi. Mol. Ecol. 2002, 11, 2367–2377. [Google Scholar] [CrossRef]
- Englbrecht, C.C.; Largiader, C.R.; Hänfling, B.; Tautz, D. Isolation and characterization of polymorphic microsatellite loci in the European bullhead Cottus gobio L. (Osteichthyes) and their applicability to related taxa. Mol. Ecol. 1999, 8, 1966–1969. [Google Scholar] [CrossRef]
- Nolte, A.W.; Stemshorn, K.C.; Tautz, D. Direct Cloning of Microsatellite Loci from Cottus Gobio through a Simplified Enrichment Procedure. Mol. Ecol. Notes 2005, 5, 628–636. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): A population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rice, W.E. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Park, S.D.E. Trypanotolerance in West African Cattle and the Population Genetic Effects of Selection. Ph.D. Thesis, University of Dublin, Dublin, Ireland, 2001. [Google Scholar]
- Weir, B.S.; Cockerham, C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [PubMed]
- Cavalli-Sforza, L.L.; Edwards, A.W.F. Phylogenetic Analysis. Models and Estimation Procedures. Am. J. Hum. Genet 1967, 19, 233–257. [Google Scholar]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package) Version 3.5c. Distributed by the Author; Department of Genetics. University of Washington: Seattle, DC, USA, 1993. Available online: http://www.washington.edu/ (accessed on 14 January 2023).
- Page, R.D.M. TREEVIEW: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Waples, R.S. A bias correction for estimates of effective population size based on linkage disequilibrium at unlinked gene loci. Conserv. Genet. 2006, 7, 167–184. [Google Scholar] [CrossRef]
- Waples, R.S.; Do, C. LDNE: A program for estimating effective population size from data on linkage disequilibrium. Mol. Ecol. Resour. 2008, 8, 753–756. [Google Scholar] [CrossRef]
- England, P.R.; Cornuet, J.M.; Berthier, P.; Tallmon, D.; Luikart, G. Estimating effective population size from linkage disequilibrium: Severe bias in small sizes. Conserv. Genet. 2006, 7, 303–308. [Google Scholar] [CrossRef]
- Waples, R.S. Genetic estimates of contemporary effective population size: To what time periods do the estimates apply? Mol. Ecol. 2005, 14, 3335–3352. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Piry, S.; Luikart, G.; Cornuet, J.M. Bottleneck: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Hendricks, P. Status, Distribution, and Biology of Sculpins (Cottidae) in Montana: A Review; Montana Natural Heritage Program: Helena, MT, USA, 1997; 29p. [Google Scholar]
- Griffith, J.S.; Kuda, D.B. Distribution, Habitat Use, and Reproductive Ecology of the Shoshone sculpin (Cottus greenei); Technical Appendix E.3.1-C for New License Application: Upper Salmon Falls (FERC No. 2777), Lower Salmon Falls (FERC No. 2061), Bliss (FERC No. 1975); Idaho Power Company: Boise, ID, USA, 1994; Volume 1, 130p. [Google Scholar]
- Wright, S. Evolution in Mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef] [PubMed]
- Waples, R.S.; Do, C. Linkage disequilibrium estimates of contemporary Ne using highly variable genetic markers: A largely untapped resource for applied conservation and evolution. Evol. Appl. 2010, 3, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Luikart, G.; Cornuet, J.M.; Allendorf, F.W. Temporal changes in allele frequencies provide estimates of population bottleneck size. Conserv. Biol. 1999, 13, 523–530. [Google Scholar] [CrossRef]
- Moritz, C.; Sherwin, W.B. Genetics and the Conservation of Wild Populations; Sinauer Associates: Sunderland, MA, USA, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Population | Collection Site # | Year | N | HE | HO | NA | NE | NE (95% L) | NE (95% U) |
|---|---|---|---|---|---|---|---|---|---|
| Montana Mining Ditch | 1 | 2009 | 35 | 0.38 | 0.37 | 3.1 | −131.8 | 68.2 | ∞ |
| Decker/Sullivan | 2 | 2009 | 70 | 0.37 | 0.38 | 3.4 | −6256.8 | 95 | ∞ |
| Unm. Pottery House | 3 | 2009 | 53 | 0.35 | 0.34 | 3.1 | 441.1 | 61.3 | ∞ |
| Malad River | 4 | 2008 | 49 | 0.50 | 0.49 | 4.9 | −1972.7 | 128.4 | ∞ |
| Lower White Springs | 5 | 2009 | 80 | 0.48 | 0.49 | 4.9 | 937.5 | 126.5 | ∞ |
| Billingsley Creek | 6 | 2008 | 50 | 0.33 | 0.32 | 2.7 | 114.5 | 34.1 | ∞ |
| Fisher Lake | 7 | 2010 | 50 * | 0.36 | 0.36 | 4.9 | 149.8 | 37 | ∞ |
| Riley Creek (upper) | 8 | 2009 | 57 | 0.39 | 0.38 | 4.1 | 687.9 | 100.1 | ∞ |
| Riley Creek (lower) | 9 | 2009 | 54 | 0.62 | 0.61 | 6.7 | 271.2 | 82.1 | ∞ |
| Bickel Springs (upper) | 10 | 2008 | 50 | 0.36 | 0.35 | 3.3 | 131.9 | 32.6 | ∞ |
| Bickel Springs (lower) | 11 | 2009 | 50 | 0.61 | 0.59 | 6.4 | 19,674.3 | 224.5 | ∞ |
| Thousand Springs | 12 | 2008 | 50 | 0.62 | 0.59 | 7.2 | 132.4 | 69.2 | 605.3 |
| Sculpin Springs | 13 | 2008 | 50 | 0.62 | 0.62 | 6.7 | 175 | 77.2 | ∞ |
| Sand Springs | 14 | 2008 | 50 | 0.62 | 0.59 | 7.0 | −1169.3 | 226.1 | ∞ |
| Blue Hearts Springs | 15 | 2008 | 23 | 0.55 | 0.59 | 4.4 | 270.4 | 43.5 | ∞ |
| Box Canyon (lower) | 16 | 2008 | 50 | 0.57 | 0.56 | 6.1 | 131.1 | 50.8 | ∞ |
| Box Canyon (upper) | 17 | 2008 | 49 | 0.46 | 0.43 | 4.3 | 150.8 | 56.4 | ∞ |
| Blind Canyon | 18 | 2009 | 55 | 0.56 | 0.55 | 5.8 | 1485.7 | 106 | ∞ |
| Banbury Springs | 19 | 2008 | 56 | 0.33 | 0.32 | 4.3 | 186.2 | 48.3 | ∞ |
| Briggs Creek | 20 | 2009 | 65 | 0.22 | 0.22 | 2.4 | −230.3 | 83.2 | ∞ |
| Panel | Locus | Primer Names | Dye | Primer Sequences | Range b.p. | AO | DM | Reference |
|---|---|---|---|---|---|---|---|---|
| A | Cba42 | Cba42F | VIC | AAATGGTCGTCTGCTCCCTG | 110–129 | 9 | N | [15] |
| Cba42R | AGGCAGTGTGGGCATGAAAG | |||||||
| A | Cott100 | Cott100F | NED | CTCATCGTGGTTTGATCGGTG | 178–194 | 7 | Y | [17] |
| Cott100-PIG R | CCGAGCGTGAGTCAGGCGTG | |||||||
| A | Cott105 | Cott105F | NED | TCCTACAGGGTGCGATCGTG | 305–320 | 7 | N | [17] |
| Cott105R | TGCAGGAGTCAGGACTCTGC | |||||||
| A | Cott113 | Cott113F | 6FAM | AGCGCCAGAATGCAGCATCC | 139–166 | 10 | Y | [17] |
| Cott113R | AGTGTGGCGAGCCCAAGATC | |||||||
| A | Cott130 | Cott130F | PET | TCTGGATCCCTCGGACCAGG | 152–177 | 10 | Y | [17] |
| Cott130-PIG R | TGAGCTCCATCGTGGGTTCG | |||||||
| A | Cott207 | Cott207F | PET | AGTCCTTGTCGGGAGCCTCG | 299–347 | 18 | N | [17] |
| Cott207R | ATTGGGCGTTGCTCACCAGC | |||||||
| A | CottES10 | CottES10F | VIC | CAGGCGGCGACACGGTG | 175–197 | 10 | N | [17] |
| CottES10R | TTATGAGGAGTCTGCCAATGCAG | |||||||
| B | Cgo310 | Cgo310F | 6FAM | AGAACCAGTGTTTGACTCTGC | 181–209 | 13 | Y | [16] |
| Cgo310R | CACTGTCATGTAGCGGCTC | |||||||
| B | Cgo1114 | Cgo1114F | 6FAM | GTGACTGAGCCTTGAGATTC | 109–147 | 15 | N | [16] |
| Cgo1114R | GAACCAACGGAAATGAAAC | |||||||
| B | Cgo33 | Cgo33F | PET | CAAAAGACAGACCTGTTGAC | 153–193 | 13 | N | [16] |
| Cgo33-PIG R | TTAACAGTGAAGGATGTGAG | |||||||
| B | Cott118 | Cott118F | PET | ACTGGTCTCCAGGCGGTGTC | 383–395 | 6 | Y | [17] |
| Cott118R | GACGCCGTCATGCTCAGGTC | |||||||
| B | LCE89 | LCE89F | 6FAM | AGAGCACACACCCTTCCGGTC | 260–328 | 12 | N | [17] |
| LCE89R | GAACCTGCACAGGGCTACAGC |
| Population | Montana 1 Mining Ditch | Decker 2/ Sullivan | Unm. 3 Pottery House | Malad 4 River | Lower 5 White Springs | Billingsley 6 Creek | Fisher 7 Lake | Riley 8 Creek (upper) | Riley 9 Creek (lower) | Bickel 10 Springs (upper) | Bickel 11 Springs (lower) | Thousand 12 Springs | Sculpin 13 Springs | Sand 14 Springs | Blue 15 Hearts Springs | Box 16 Canyon (lower) | Box 17 Canyon (upper) | Blind 18 Canyon | Banbury 19 Springs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Decker/Sullivan 2 | 0.01 | ||||||||||||||||||
| Unm. Pottery House 3 | 0.06 | 0.04 | |||||||||||||||||
| Malad River 4 | 0.21 | 0.24 | 0.23 | ||||||||||||||||
| Lower White Springs 5 | 0.20 | 0.23 | 0.22 | 0.02 | |||||||||||||||
| Billingsley Creek 6 | 0.26 | 0.27 | 0.27 | 0.17 | 0.15 | ||||||||||||||
| Fisher Lake 7 | 0.25 | 0.27 | 0.27 | 0.20 | 0.17 | 0.01 | |||||||||||||
| Riley Creek (upper) 8 | 0.12 | 0.14 | 0.19 | 0.26 | 0.22 | 0.21 | 0.21 | ||||||||||||
| Riley Creek (lower) 9 | 0.26 | 0.30 | 0.29 | 0.11 | 0.12 | 0.24 | 0.28 | 0.28 | |||||||||||
| Bickel Springs (upper) 10 | 0.23 | 0.25 | 0.25 | 0.23 | 0.19 | 0.12 | 0.14 | 0.12 | 0.26 | ||||||||||
| Bickel Springs (lower) 11 | 0.29 | 0.32 | 0.31 | 0.11 | 0.13 | 0.26 | 0.29 | 0.30 | <0.00 | 0.28 | |||||||||
| Thousand Springs 12 | 0.30 | 0.34 | 0.33 | 0.12 | 0.14 | 0.27 | 0.31 | 0.32 | <0.00 | 0.30 | <0.00 | ||||||||
| Sculpin Springs 13 | 0.30 | 0.33 | 0.33 | 0.12 | 0.15 | 0.28 | 0.32 | 0.31 | <0.00 | 0.30 | <0.00 | <0.00 | |||||||
| Sand Springs 14 | 0.30 | 0.33 | 0.33 | 0.13 | 0.15 | 0.29 | 0.32 | 0.32 | 0.01 | 0.30 | <0.00 | <0.00 | <0.00 | ||||||
| Blue Hearts Springs 15 | 0.35 | 0.38 | 0.40 | 0.18 | 0.21 | 0.39 | 0.41 | 0.37 | 0.09 | 0.37 | 0.10 | 0.09 | 0.06 | 0.07 | |||||
| Box Canyon (lower) 16 | 0.32 | 0.36 | 0.37 | 0.16 | 0.18 | 0.36 | 0.40 | 0.35 | 0.07 | 0.35 | 0.08 | 0.08 | 0.05 | 0.06 | <0.00 | ||||
| Box Canyon (upper) 17 | 0.37 | 0.42 | 0.43 | 0.26 | 0.27 | 0.44 | 0.46 | 0.41 | 0.17 | 0.42 | 0.18 | 0.17 | 0.16 | 0.17 | 0.13 | 0.09 | |||
| Blind Canyon 18 | 0.32 | 0.35 | 0.36 | 0.16 | 0.19 | 0.35 | 0.39 | 0.35 | 0.07 | 0.34 | 0.09 | 0.08 | 0.06 | 0.07 | <0.00 | <0.00 | 0.10 | ||
| Banbury Springs 19 | 0.51 | 0.52 | 0.53 | 0.30 | 0.33 | 0.51 | 0.51 | 0.51 | 0.20 | 0.51 | 0.20 | 0.18 | 0.17 | 0.18 | 0.15 | 0.14 | 0.25 | 0.13 | |
| BriggsCreek 20 | 0.60 | 0.59 | 0.62 | 0.38 | 0.39 | 0.60 | 0.55 | 0.59 | 0.29 | 0.59 | 0.31 | 0.29 | 0.26 | 0.27 | 0.24 | 0.20 | 0.36 | 0.19 | 0.15 |
| N | NE | NE(95%L) | NE(95%U) |
|---|---|---|---|
| 50 | 149.8 | 37.0 | ∞ |
| 100 | 329.9 | 91.9 | ∞ |
| 150 | 4119.4 | 207.5 | ∞ |
| 200 | −1473.6 | 487.8 | ∞ |
| 250 | 1729.6 | 277.8 | ∞ |
| 300 | 2156.2 | 401.1 | ∞ |
| Population | Collection Site # | Sign Test TPM | Sign Test SMM | Wilcoxon Test (Deficiency) TPM | Wilcoxon Test (Excess) TPM | Wilcoxon Test (Deficiency) SMM | Wilcoxon Test (Excess) SMM |
|---|---|---|---|---|---|---|---|
| Montana Mining Ditch | 1 | 0.57 | 0.31 | 0.52 | 0.52 | 0.31 | 0.72 |
| Decker/Sullivan | 2 | 0.30 | 0.12 | 0.31 | 0.72 | 0.12 | 0.90 |
| Unm. Pottery House | 3 | 0.32 | 0.13 | 0.28 | 0.74 | 0.08 | 0.94 |
| Malad River | 4 | 0.13 | 0.04 D8/12 | 0.21 | 0.82 | 0.01 | 0.99 |
| Lower White Springs | 5 | 0.08 | 0.00 D10/12 | 0.06 | 0.95 | 0.00 | 1.00 |
| Billingsley Creek | 6 | 0.47 | 0.24 | 0.72 | 0.31 | 0.28 | 0.75 |
| Fisher Lake | 7 | 0.02 D9/12 | 0.00 D11/12 | 0.02 | 0.99 | 0.00 | 1.00 |
| Riley Creek (upper) | 8 | 0.21 | 0.07 | 0.22 | 0.81 | 0.01 | 0.99 |
| Riley Creek (lower) | 9 | 0.07 | 0.00 D10/12 | 0.10 | 0.91 | 0.00 | 1.00 |
| Bickel Springs (upper) | 10 | 0.54 | 0.25 | 0.31 | 0.72 | 0.10 | 0.92 |
| Bickel Springs (lower) | 11 | 0.36 | 0.00 D10/12 | 0.34 | 0.69 | 0.00 | 1.00 |
| Thousand Springs | 12 | 0.16 | 0.00 D112/12 | 0.12 | 0.90 | 0.00 | 1.00 |
| Sculpin Springs | 13 | 0.17 | 0.00 D11/12 | 0.31 | 0.72 | 0.00 | 1.00 |
| Sand Springs | 14 | 0.07 | 0.00 D10/12 | 0.09 | 0.92 | 0.00 | 1.00 |
| Blue Hearts Springs | 15 | 0.08 | 0.08 | 0.26 | 0.77 | 0.05 | 0.96 |
| Box Canyon (lower) | 16 | 0.00 D10/12 | 0.00 D11/12 | 0.00 | 1.00 | 0.00 | 1.00 |
| Box Canyon (upper) | 17 | 0.20 | 0.02 | 0.10 | 0.91 | 0.01 | 0.99 |
| Blind Canyon | 18 | 0.02 D9/12 | 0.00 D10/12 | 0.06 | 0.95 | 0.00 | 1.00 |
| Banbury Springs | 19 | 0.02 D9/12 | 0.00 D10/12 | 0.00 | 1.00 | 0.00 | 1.00 |
| Briggs Creek | 20 | 0.15 | 0.14 | 0.14 | 0.88 | 0.07 | 0.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, M.R.; Tretter, E.D.; Trainer, J.C.; Wilkison, R.A. Genetic Diversity and Population Structure of Shoshone Sculpin Cottus greenei in the Hagerman Valley of South-Central Idaho. Fishes 2023, 8, 55. https://doi.org/10.3390/fishes8010055
Campbell MR, Tretter ED, Trainer JC, Wilkison RA. Genetic Diversity and Population Structure of Shoshone Sculpin Cottus greenei in the Hagerman Valley of South-Central Idaho. Fishes. 2023; 8(1):55. https://doi.org/10.3390/fishes8010055
Chicago/Turabian StyleCampbell, Matthew R., Eric D. Tretter, James C. Trainer, and Richard A. Wilkison. 2023. "Genetic Diversity and Population Structure of Shoshone Sculpin Cottus greenei in the Hagerman Valley of South-Central Idaho" Fishes 8, no. 1: 55. https://doi.org/10.3390/fishes8010055
APA StyleCampbell, M. R., Tretter, E. D., Trainer, J. C., & Wilkison, R. A. (2023). Genetic Diversity and Population Structure of Shoshone Sculpin Cottus greenei in the Hagerman Valley of South-Central Idaho. Fishes, 8(1), 55. https://doi.org/10.3390/fishes8010055