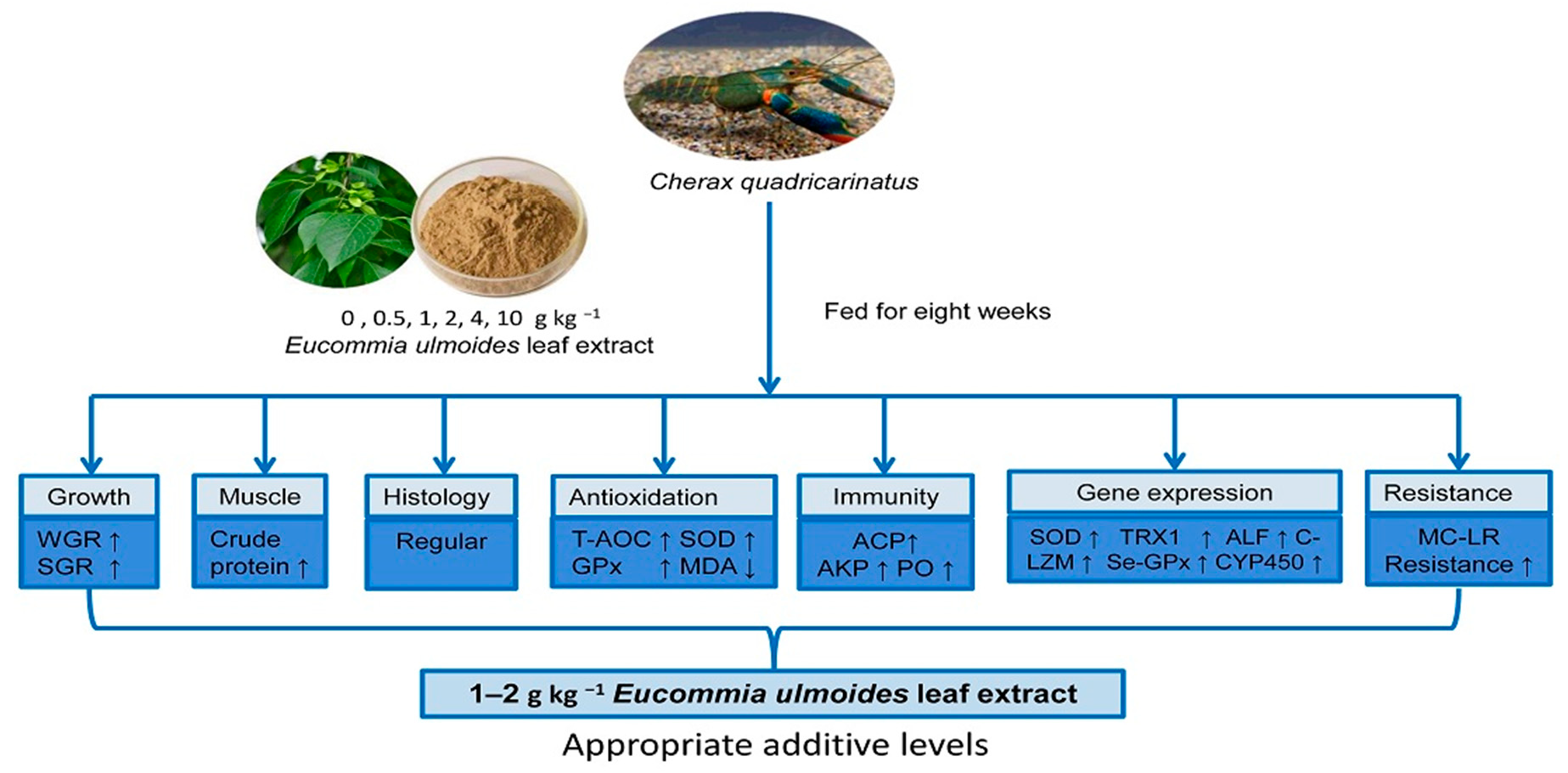
1. Introduction
The red claw crayfish (
Cherax quadricarinatus) is suitable for intensive or semi-intensive culture in many countries, including Australia, America, Mexico, Argentina, Cuba, and China [
1]. It is one of the most popular crayfish species widely farmed in Guangdong, Fujian, Hainan, and Jiangsu Provinces in China because of its high resistance, fast growth, large size, and ease of farming [
2,
3,
4]. With the continuous development of the high-density intensive culture of aquaculture, the water environment has become a major concern for farmers [
5]. Deterioration of the water environment, especially the eutrophication of aquaculture water, can affect aquatic species’ immunity, survival, and growth [
6], leading to the occurrence of diseases. To avoid the outbreak of diseases, farmers frequently use chemicals and antimicrobials to prevent viral, fungal, and bacterial diseases. However, the abusive use of chemicals and antimicrobials can lead to the formation of drug residues, disruption of immune balance in aquatic animals, microbial resistance, immunosuppression, and environmental pollution, which affects crayfish culture [
7]. The application of green plant additives to aquatic species is the key to the sustainable development of aquaculture [
8]. Therefore, Chinese herbal extracts have become suitable alternatives to chemicals and antimicrobials for protecting aquaculture environments.
Cyanobacteria bloom can threaten water quality in many eutrophic ponds [
9]. A cyanobacterial harmful algal bloom is one of the common microbial hazards in freshwater aquaculture and has become a serious global public health and environmental problem [
10,
11,
12]. Toxins produced by cyanobacteria endanger the health of freshwater aquaculture animals and the safety of aquatic products [
13]. The toxins of freshwater cyanobacteria, including cyclic-peptide microcystins and nodularin, are divided into neurotoxins and hepatotoxins. Microcystins (MCs) are a group of cyanobacterial hepatotoxins which have more than 279 kinds of different MC congeners, and microcystin-LR (MC-LR) is the most common and potently toxic MC [
14,
15,
16,
17]. MC-LR can damage the hepatopancreas, head kidney, spleen, etc. [
18,
19,
20,
21] and cause intrahepatic hemorrhage [
22] and extensive centrilobular necrosis [
23]. Besides, microcystins can suppress protein phosphatase expression, leading to increased protein phosphorylation, which is directly related to their cytotoxicity and immunotoxicity [
23,
24,
25]. The immune system can be altered by MCs through several immune-related genes and pathways [
26]. Several studies reported that MC-LR exposure significantly changed the glutathione (GSH) content and reactive oxygen species (ROS) formation [
27]. MC-LR changes the activities of various antioxidant enzymes, including glutathione reductase (GR), glutathione-s-transferase (GST), superoxide dismutase (SOD), glutathione peroxidase (GPx), and catalase (CAT) [
18,
25]. One study has shown a fast increase of SOD and CAT activities at the early stage and subsequent decrease at the later stage of MC-LR exposure, while GPx was activated by longer MC-LR exposure in
Procambarus clarkii [
18]. Exposed to the low dose of MC-LR, zebrafish exhibited a positive defensive response with an increase in the CAT, SOD, and GST activities [
25]. On the other hand, high MC-LR dose in zebrafish resulted in a negative antioxidant response by inhibition in the CAT, GPx, and GST activities. GSH depletion in both dose group suggested the crucial role of GSH in cellular antioxidant protection and MC-LR detoxification [
25].
Plant extracts can be effective alternatives to chemicals and antimicrobials in aquaculture [
28,
29,
30]. Various plant extracts have been developed to improve immunity and protect tissues from a variety of environmental stress stimuli in aquaculture [
31]. Among plants,
E. ulmoides has been widely used in medicine and animal feed research [
32,
33,
34,
35].
E. ulmoides leaf extract (ELE) contains many biologically active compounds, including terpenoids, phenolics, lignans, steroids, iridoids, and flavonoids [
36,
37]. Chlorogenic acid is the principal bioactive component of ELE that scavenges free radicals and improves antioxidant ability and immunity [
38,
39].
E. ulmoides has several beneficial functions, including the antioxidant effect [
40,
41], immunity regulation [
42,
43], and antibacterial [
44] and anti-inflammatory [
32]. However, few published reports about the application of ELE in crayfish feed [
45]. We studied the effect of ELE on growth performance, muscle composition, hepatopancreas histology, immune responses, gene expression, and disease resistance of juvenile red claw crayfish, which could promote the development of pond culture in red claw crayfish. This is the first time to apply ELE to crayfish feed research, laying the foundation for the screening of crayfish feed additives.
2. Materials and methods
2.1. Diet Preparation
The composition of the basal diet is shown in
Table 1.
E. ulmoides leaf extract (ELE) (alcoholic extract) was added to the diet at 0 (Diet 1), 0.5 (Diet 2), 1 (Diet 3), 2 (Diet 4), 4 (Diet 5), and 10 (Diet 6) g dry weight (dw) ELE kg (dw)
−1 diets. ELE was provided by Shandong Longchang Biotechnology Co. Ltd. (Shandong, China). Preparation of ethanol extract of ELEs: Firstly,
Eucommia ulmoides leaves are fully mixed with 70% ethanol at a ratio of 1:12, extracted twice by heating reflux under 60–70 °C, and then the ethanol extract is obtained by chromatographic purification. The main components of ELE are 5% chlorogenic acid, 8% flavonoids, and 20% polysaccharides. All the ingredients of the diet were ground, sifted through 80 mesh, and mixed with oil. Then deionized water (40%) was added, and the 1.5 mm diameter feed was wet-extruded by a pelletizer. Subsequently, all diets were air-dried to below 100 g moisture kg
−1 diet. After drying, the diets were stored at −20 °C until used.
2.2. Experimental Crayfish and Sample Collection
The red claw crayfish purchased from a commercial farm in Hainan Province (Wenchang, China) were transported to the experimental condition in cylindrical tanks and were fed with the basal diet for two weeks to acclimate to the experimental diet and conditions. A total of 720 healthy crayfish with an average initial body weight of 0.24 ± 0.01 g were randomly divided into eighteen tanks (500 L; three tanks per group) with three replicates in each group and 40 crayfish in each replicate. To prevent crayfish cannibalism, 40 PVC pipes (Nominal diameter 150, length 10) were placed in each tank and glued together. The crayfish were fed diets two times each day at 8:00 and 18:00. The crayfish were fed every day until satiation. The feeding trial lasted for 8 weeks. During the experimental period, water temperature, pH, ammonia-N, and nitrite-N were 26–29 °C, 7.2–7.7, 0.05–0.11 mg L−1, and 0.05–0.10 mg L−1, respectively. Dissolved oxygen was maintained at 5.0–7.3 mg L−1.
After the culture experiment, crayfish were fasted for 24 h before sampling. The number, body length, and body weight of crayfish were measured. Hemolymph, hepatopancreas, and back muscle samples were collected as described previously [
46].
2.3. Growth Performance
The weight gain rate (WGR), specific growth rate (SGR), and survival rate (SR) were calculated as follows [
47]:
2.4. Proximate Composition Analysis
Muscle and diet compositions were analyzed according to the established methods of the Association of Official Analytical Chemists [
48]. Crude protein content was determined by the Kjeldahl method using Kjeltec
TM 8400 (Kjeltec, Foss, Denmark) after digesting the samples with sulfuric acid. Crude lipid was determined by the Soxhlet extraction method using a lipid analyzer (SZF-06A, Hongji, Shanghai, China). Feed and muscle samples were placed in a muffle furnace (SX2-2.5-10, Yiheng, Shanghai, China) at 550 °C for 12 h to determine the ash content. Feed and muscle moisture contents were determined by oven drying at 105 °C.
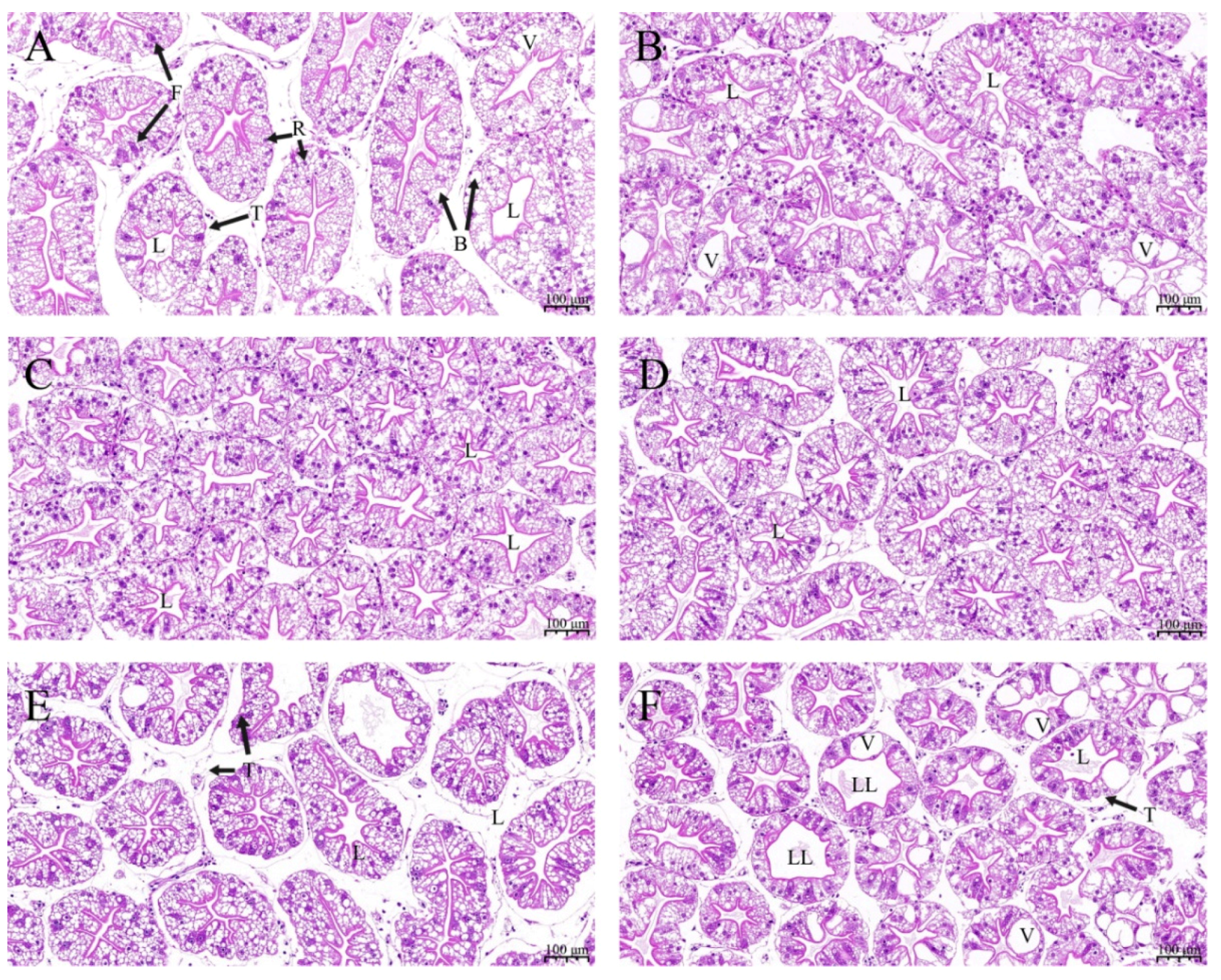
2.5. Histopathological Analysis
Three hepatopancreas samples from each tank were collected for morphological analysis. All tissue samples were fixed and stored in 4% paraformaldehyde (Biosharp, Beijing, China) for 48 h. Then, all samples were dehydrated in graded ethanol concentration series and paraffin-embedded. Consequently, the pieces were stained with hematoxylin and eosin (H&E) for optical examination. Morphological changes in the hepatopancreas were observed using Nikon Eclipse E100 Microscope (Nikon, Tokyo, Japan).
2.6. Antioxidant and Immune Enzyme Assays
GPx, GSH, CAT, GST, SOD, alkaline phosphatase (AKP), and acid phosphatase (ACP) activities, and total antioxidant capacity (T-AOC) and malondialdehyde (MDA) contents were measured using their respective kits following the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu, China). Phenoloxidase (PO) content was measured by the double antibody sandwich enzyme-linked immunosorbent assay (ELISA) method using commercial kits (Dongguan Enzyme-linked Biotechnology Co., Ltd., Dongguan, Guangdong, China).
2.7. RNA Extraction and Gene Expression Analysis
Hepatopancreas samples from three crayfish in each tank were used for total RNA isolation by TRIzol reagent (Invitrogen, Waltham, MA, USA). The quantity of each RNA was measured by NanoDrop One spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA), and RNA integrity was detected using agarose gel (1%) electrophoresis. Subsequently, the complementary DNA (cDNA) was synthesized from total RNA using a PrimeScript RT reagent Kit With gDNA Eraser (Takara, Dalian, China) according to the manufacturer’s instructions. The cDNA was stored at −20 °C for real-time quantitative PCR (qRT-PCR) analysis.
The appropriate primers of each gene (
Table 2) were designed based on our transcriptome unigenes and references [
49] using Primer Premier v5 (PREMIER Biosoft International (Palo Alto, CA, USA). The real-time PCR reactions were carried out using 2× SYBR
® Green qPCR Mix buffer (Takara, Dalian, China) on a Stratagene Mx3005P real-time PCR machine (Agilent, Santa Clara, CA, USA). The cycling parameters were as follows: 94 °C for 3 min, followed by 40 cycles of 94 °C for 15 s, 58 °C for 15 s, and 72 °C for 20 s. The expression results were calculated by the 2
−ΔΔCt method [
50].
2.8. MC-LR LD50 Determination
The MC-LR (Purity ≥ 95%) was obtained from Algal Science (Algal Science Co. Ltd., Taoyuan, Taiwan). MC-LR was suspended in 0.85% NaCl solution. The concentration of the pre-experiment was 0, 30.2, 42.66, 60.26, 85.11, 120.11, 169.82, and 239.88 μg kg
−1. The lethal dose 50 (LD
50) LD
50 was determined using the Statistical Package for the Social Sciences (SPSS) 18.0 software (SPSS, Chicago, IL, USA) with the regression (Probit analysis method). The mortality rates of each dose of the pre-experiment are shown in
Table 3. The LD
50s are shown in
Table 4.
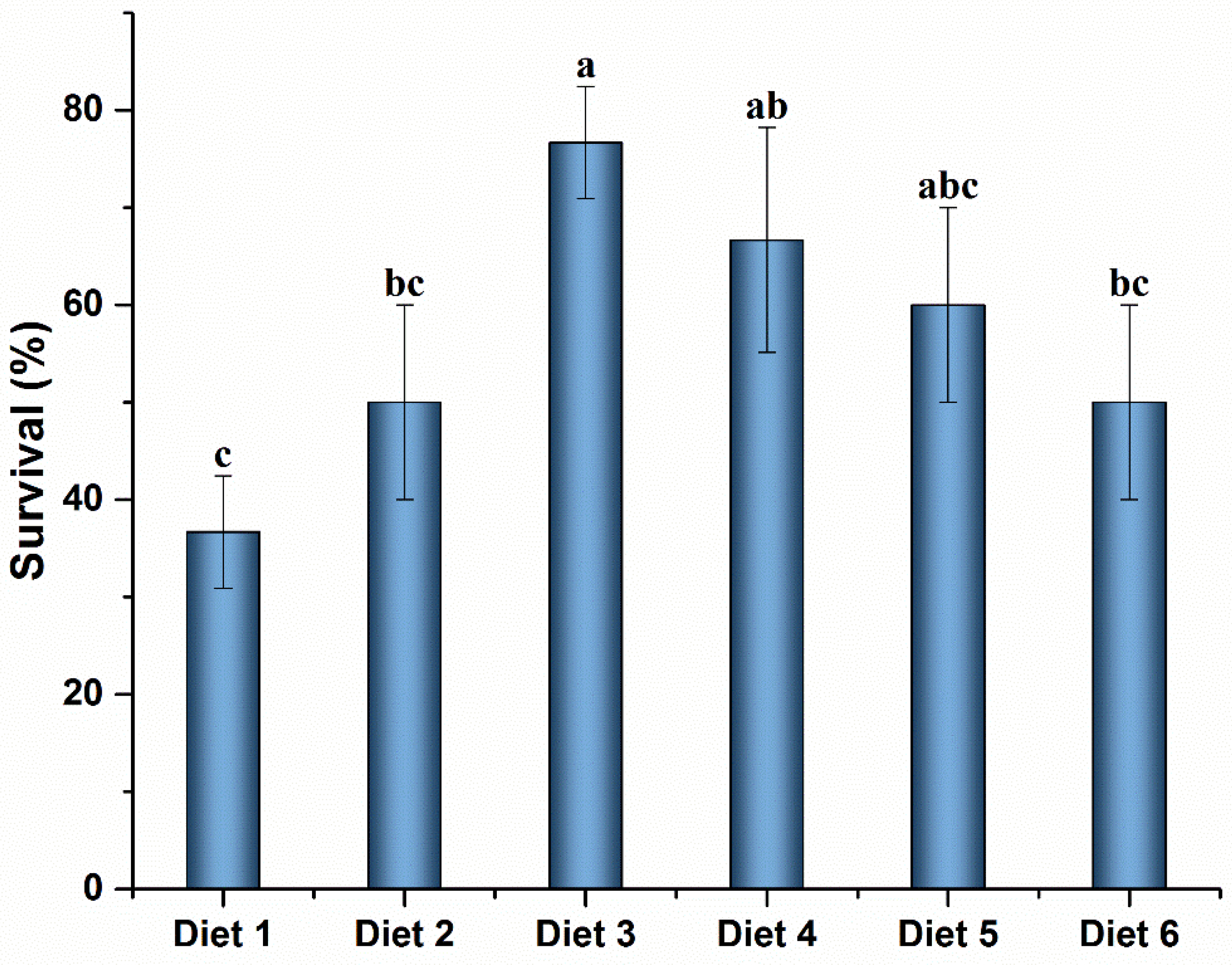
2.9. MC-LR Challenge
At the end of the breeding trial, ten crayfish per tank were randomly selected for MC-LR challenge experiments, and the muscular injection dose was 108 μg MC-LR kg
−1 BW (body weight). The dose of MC-LR used in the study was based on the result from the 48-h lethal doses 50 (LD
50) of MC-LR in the pre-experiment of red claw crayfish (
Table 4). After MC-LR injection, crayfish mortality was recorded at 48 h, and SR was then determined. The surviving crayfish hepatopancreas was dissected and stored at −80 °C.
2.10. Statistical Analysis
The results were expressed as means ± standard deviation (SD). One-way analysis of variance (ANOVA) was used to analyze the data. SPSS 18.0 software (SPSS, Chicago, IL, USA) was used to analyze the data. The significant differences among means were detected using Tukey’s multiple range tests at a significance level of 0.05.
4. Discussion
The potential efficacy of plant extracts for abiotic stress and disease prevention has received widespread attention in aquaculture. Few studies have been reported on the application of ELE in aquatic animal feeds. We reported the effects of dietary ELE on growth, muscle composition, non-specific immunity, and disease resistance of juvenile red claw crayfish. In the present study, a diet supplemented with 1–2 g dw ELE kg (dw)
−1 diet significantly increased the WGR and SGR of crayfish, which was similar to previous studies in weaned piglets [
51] and broilers [
39]. Supplementation with a 4–10 g dw ELE kg (dw)
−1 diet did not significantly improve the growth performance of crayfish, which indicates that the beneficial effects of plant supplementation on crayfish growth are dose-dependent. If optimal doses are exceeded, the benefits might be lost [
52,
53]. The supplementation of excessive ELE in the feed could affect the metabolism of crayfish, leading to high energy consumption, stress, and toxic effects on crayfish [
54]. Compared with the control group, dietary treatment of 1 g dw ELE kg (dw)
−1 diet increased crude protein content and decreased crude lipid content in crayfish muscle. The results are consistent with previous studies on plant extracts [
54,
55]. Sufficient nutrient utilization improves the physiological metabolism of crayfish, affecting the metabolism of amino acids, lipids, and glycogen in the tissue. Crayfish fed diets with proper ELE levels alter protein and lipid utilization, causing lipid consumption and protein deposition. Our results indicated that dietary ELE could optimize the utilization of nutrients and increase protein synthesis in crayfish.
The hepatopancreas is an important organ in crustaceans and plays an important role in digestion, immune defense, nutrient absorption, and metabolism [
56,
57]. The nutritional status can be assessed by observing changes in the morphology of the hepatopancreas in crustaceans [
58,
59]. Previous studies reported that dietary ELE could significantly reduce an increase in lipid accumulation and preserve liver function in chickens and rats [
60,
61]. In this study, crayfish fed Diet 3 and Diet 4 showed regular hepatocyte morphology. However, hepatocytes of crayfish in the Diet 6 group showed cell swelling and mild vacuolation. The increased vacuolation of the crayfish hepatopancreas may be caused by the complex composition of ELE, suggesting that extra ELE may damage the crayfish hepatopancreas, which may be the reason for not improving the growth performance of crayfish fed Diet 6 [
62]. Inadequate crayfish feed formulation may cause mild vacuolation of the hepatopancreas in the Diet 1 and Diet 2 groups. These results indicated that dietary supplementation with 1–2 g dw ELE kg (dw)
−1 diet could improve the morphology of the hepatopancreas with the improvement of absorptive function for various nutrients.
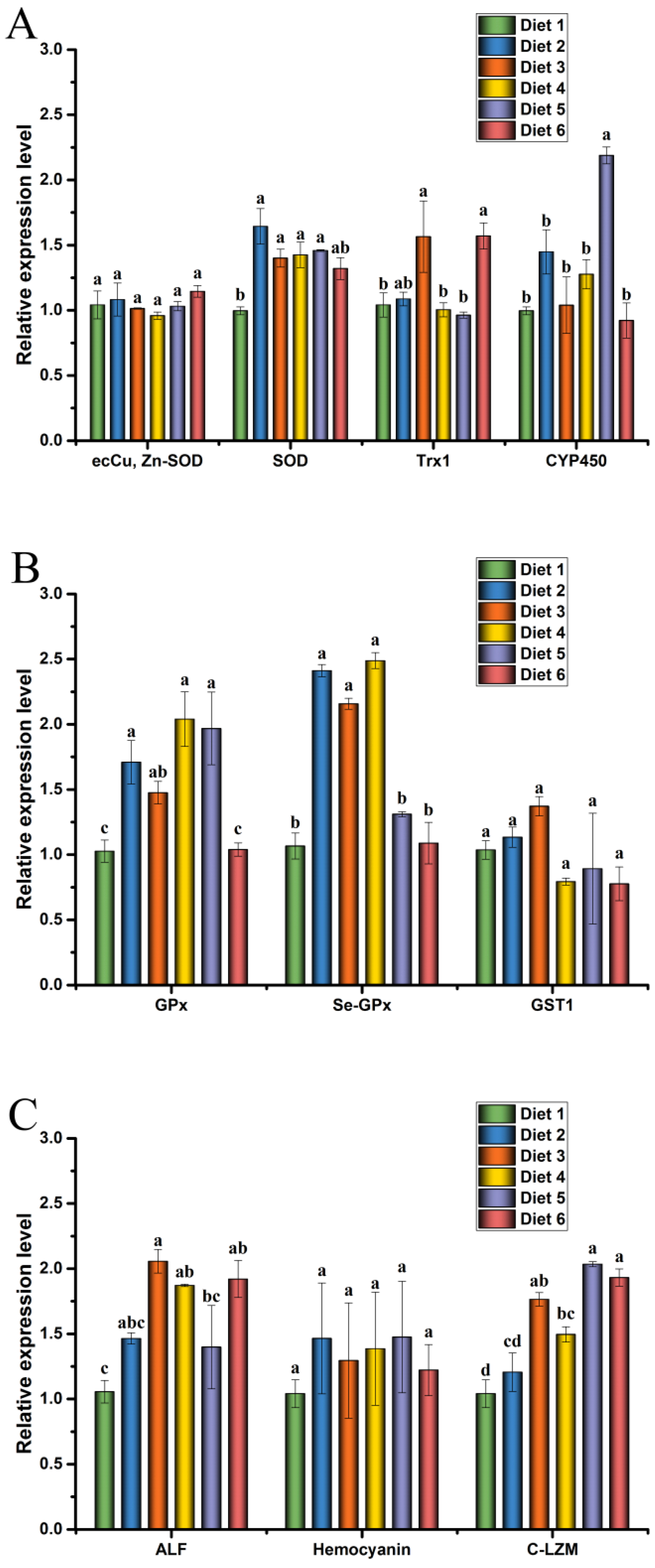
The antioxidant system plays a crucial part in protecting crayfish from oxidative stress [
18]. Antioxidant enzymes are important components of the antioxidant system in crustaceans. The T-AOC refers to the antioxidant capacity of the enzymatic and non-enzymatic antioxidants of crustaceans [
63]. The SOD, GPx, and CAT are important antioxidant enzymes of aquatic animals and can eliminate unnecessary free superoxide anion radicals [
34,
64]. MDA can induce tissue damage, reflecting the degree of lipid peroxidation and cell damage [
65]. The results showed that T-AOC, GPx and SOD activities significantly increased, and the MDA content decreased in the hemolymph and the hepatopancreas of crayfish fed the optimal ELE level, which is similar to the findings in grass carp [
66]. Yang et al. reported that the supplementation of ELE in diets significantly increased the activities of SOD and GPx in grass carp [
66]. Previous studies reported that plant extract supplements could improve the antioxidant capacity of red swamp crayfish (
Procambarus clarkii) [
67,
68]. The trends in the change expression levels of antioxidant genes were consistent with the enzyme activity analysis in this study. Our results showed that dietary ELE significantly up-regulated the expression levels of SOD, GPx, and Se-GPx in the hepatopancreas of crayfish. Tan et al. reported that dietary Ginkgo (
Ginkgo biloba) leaf extract could improve antioxidant ability in the head kidney of hybrid grouper (
Epinephelus lanceolatus
×
Epinephelus fuscoguttatus
) by increasing antioxidant gene expression [
62]. Consistent with these studies, our findings indicated that ELE supplementation in the diet could protect the hepatocytes and hemocytes from oxidative protein carbonation and lipid peroxidation by activating the antioxidant response in red claw crayfish [
67].
Phosphatases are crucial enzymes in the biological processes of aquatic animals and shape immune responses and reflect the immune system in aquatic animals [
69,
70]. ACP plays an essential role in substance metabolism. It is related to lysosomes and can be induced by exogenous substances [
71,
72]. AKP is the most important metabolic enzyme and plays a crucial role in the absorption and utilization of nutrients in aquatic animals, enhancing immunity of aquatic animals [
73,
74]. PO is an important enzyme in the prophenoloxidase (proPO) system, a key systemic enzyme for recognizing non-self substances in invertebrates. It is closely related to the immunity of crustaceans [
75]. In this study, dietary ELE increased the activities of ACP, AKP, and PO of crayfish. A previous study observed that the administration of Mojave yucca (
Yucca schidigera) extract enhanced ACP, AKP, and PO activities in whiteleg shrimp (
Litopenaeus vannamei) [
76]. Many plant extracts can improve the non-specific immunity of aquatic animals [
54,
68,
77]. Yang et al. reported that dietary ELE significantly improved the muscle antioxidant capacity and flesh quality of grass carp [
66]. No studies about the effects of dietary ELE on the non-specific immunity in crayfish have been reported. Further studies are needed to determine if the mechanisms underlying ELE-enhanced non-specific immunity are comparable with those found in other terrestrial animals.
Antimicrobial peptides (AMPs), such as HEM, lysozymes (LZM), and ALF, play important roles in the innate immunity of aquatic animals [
78,
79]. The regulation of immune genes is closely related to the regulation of immunity in aquatic animals [
62]. Previous studies reported that
E. ulmoides could promote immunity by regulating the secretion of proinflammatory cytokines [
32,
42]. Similarly, the high expression of AMPs implies an increase in disease resistance in aquatic animals [
80,
81,
82]. In the present study, the results suggested that diet supplement ELE significantly up-regulated the mRNA levels of C-LZM and ALF in the hepatopancreas of crayfish compared with the control group. However, the regulation of AMP gene expression by dietary ELE at the level of mRNA translation was not reported previously in aquatic animals. We demonstrated that ELE induced the expression of the AMP gene. Thus, more research needs to be done to clarify the molecular mechanism.
Plants or plant extracts can improve the resistance of aquatic animals to environmental stress, such as low temperature, toxins, and heavy metals [
73,
83,
84]. In the present study, supplementation of ELE to diets significantly improved the resistance of red claw crayfish against MC-LR stress. The increased SR of crayfish to environmental stress, as described for other aquaculture species, can result in increased health status and positive immune response of crayfish [
85]. However, few studies have reported the effects of plant extracts on immune-related genes in crayfish under MC-LR stress. Therefore, we analyzed the immune status of crayfish in each feed group under MC-LR stress. In the current study, supplementation of ELE to diets significantly up-regulated the mRNA levels of SOD, GPx, Se-GPx, GST1, ALF, and HEM in the hepatopancreas of crayfish under MC-LR stress compared with Diet 1. The antioxidant system of crayfish neutralizes the damaging effects of ROS, and the antioxidant system with GSH plays a vital role in MC-LR depuration in crayfish [
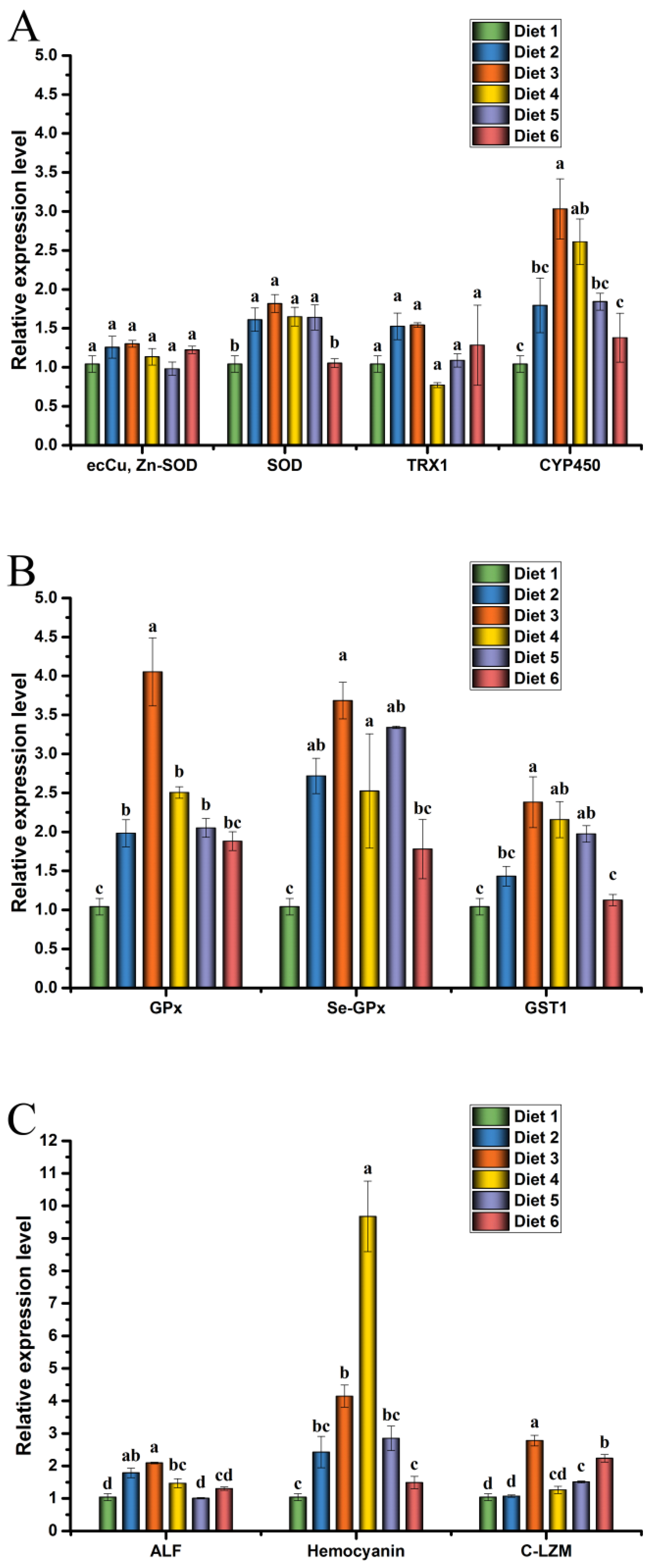
18]. In MC-LR stress experiments, the expression levels of SOD, CYP450, GPx, Se-GPx, and GST1 in crayfish were significantly increased in Diet 3 and Diet 4 groups. After a challenge with MC-LR, crayfish survival was significantly higher in Diet 3 and Diet 4 groups than in the Diet 1, which may be due to the increased effects of ELE on the immune system of crayfish [
86]. These results are similar to those reported previously [
87,
88]. Plant extracts can improve the survival of aquatic animals in stressful environments by enhancing immunity [
54,
67,
85,
89]. Xie et al. reported that Tian-Dong-Tang-Gan powder could improve the resistance of whiteleg shrimp by inducing the expression levels of GPx, Mn-SOD, and ACP in the hepatopancreas of nitrite exposure [
88]. Changes in SOD, GPx, GST, ALF, HEM, and LZM gene expression levels suggested that MC-LR induced ROS production and crayfish made the corresponding reaction by regulating these immune-related factors in vivo [
18,
87]. These results indicated that ELE could increase or maintain the mRNA expression levels of SOD, GPx, Se-GPx, GST1, ALF, HEM, and C-LZM in the hepatopancreas of juvenile red claw crayfish to regulate the ability of crayfish and resist MC-LR stress, reducing the damage of hepatopancreas caused by MC-LR in juvenile red claw crayfish.



 ,
, 

 × Epinephelus fuscoguttatus
× Epinephelus fuscoguttatus ) by increasing antioxidant gene expression [62]. Consistent with these studies, our findings indicated that ELE supplementation in the diet could protect the hepatocytes and hemocytes from oxidative protein carbonation and lipid peroxidation by activating the antioxidant response in red claw crayfish [67].
) by increasing antioxidant gene expression [62]. Consistent with these studies, our findings indicated that ELE supplementation in the diet could protect the hepatocytes and hemocytes from oxidative protein carbonation and lipid peroxidation by activating the antioxidant response in red claw crayfish [67].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}