


Genetic Diversity and Differences among Three F1 Families and Two Wild Populations of Genus Scylla Using Microsatellite Markers
 , , and
, , and 
Abstract
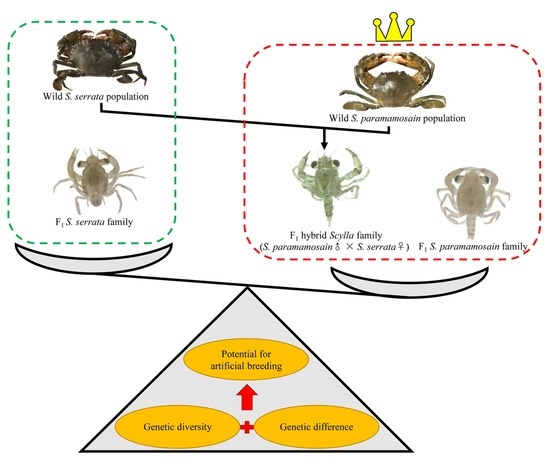
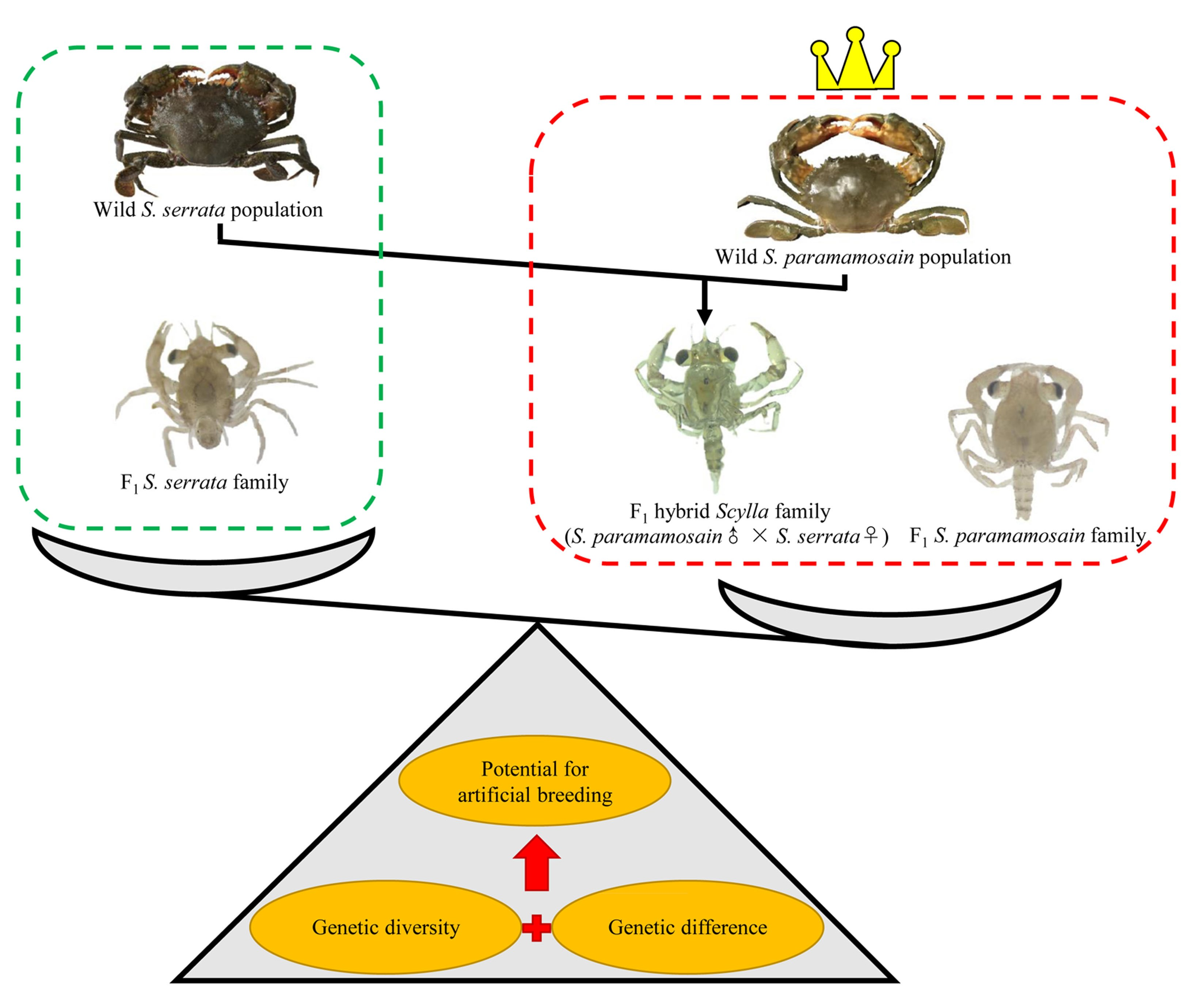
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. PCR Amplification and Electrophoresis
2.3. Genetic Analysis
3. Results
3.1. Comparative Analysis of Genetic Diversity
3.2. Population Structure and Deviations from HWE
3.3. Genetic Distance and Genetic Identity among Distinct Populations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naim, D.M.; Nor, S.A.M.; Mahboob, S. Reassessment of species distribution and occurrence of mud crab (Scylla spp., Portunidae) in Malaysia through morphological and molecular identification. Saudi. J. Biol. Sci. 2019, 27, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Yang, Q.; Zhang, Y.; Farhadi, A.; Zheng, H.; Li, S.; Zhang, Y.; Ikhwanuddin, M.; Ma, H. Integrative transcriptome sequencing reveals the molecular difference of maturation process of ovary and testis in mud crab Scylla paramamosain. Front. Mar. Sci. 2021, 8, 658091. [Google Scholar] [CrossRef]
- Meynecke, J.O.; Grubert, M.; Gillson, J. Giant mud crab (Scylla serrata) catches and climate drivers in Australia—A large scale comparison. Mar. Freshw. Res. 2012, 63, 84–94. [Google Scholar] [CrossRef]
- Ma, H.; Ma, C.; Ma, L.; Cui, H. Novel polymorphic microsatellite markers in Scylla paramamosain and cross-species amplification in related crab species. J. Crustacean Biol. 2010, 30, 441–444. [Google Scholar] [CrossRef]
- Wang, W.; Ma, C.; Chen, W.; Jin, Z.; Zhao, M.; Zhang, F.; Liu, Z.; Ma, L. Population genetic diversity of mud crab (Scylla paramamosain) from southeast coastal regions of China based on mitochondrial COI gene sequence. Gene 2020, 751, 144763. [Google Scholar] [CrossRef]
- Lin, Q. Species Composition of Genus Scylla and Genetic Diversity of Scylla paramamosain (Estampador, 1949) Population in China. Ph.D. Thesis, Xiamen University, Xiamen, China, 2009. [Google Scholar]
- Hulata, G. A review of genetic improvement of the common carp (Cyprinus carpio L.) and other cyprinids by crossbreeding, hybridization and selection. Aquaculture 1995, 129, 143–155. [Google Scholar] [CrossRef]
- Han, Z.; Li, Q.; Liu, S.; Kong, L. Crossbreeding of three different shell color lines in the Pacific oyster reveals high heterosis for survival but low heterosis for growth. Aquaculture 2020, 529, 735621. [Google Scholar] [CrossRef]
- Cui, W.; Guan, M.; Abu Sadek, M.; Wu, F.; Wu, Q.; Tan, H.; Shi, X.; Ikhwanuddin, M.; Ma, H. Construction of a genetic linkage map and QTL mapping for sex indicate the putative genetic pattern of the F1 hybrid Scylla (Scylla serrata ♀ × S. paramamosain ♂). Aquaculture 2021, 545, 737222. [Google Scholar] [CrossRef]
- Zheng, G.D.; Guo, D.D.; Wu, C.B.; Chen, J.; Jiang, X.Y.; Zou, S.M. The obvious heterosis and genetic characters of intergeneric cross and backcross juveniles between blunt snout bream (Megalobrama amblycephala) and topmouth culter (Culter alburnus). Aquac. Res. 2019, 50, 1634–1643. [Google Scholar] [CrossRef]
- Min, J.J.; Ye, R.H.; Zhang, G.F.; Zheng, R.Q. Microsatellite analysis of genetic diversity and population structure of freshwater mussel (Lamprotula leai). Zool. Res. 2015, 36, 34–40. [Google Scholar]
- Feng, Y.; Zhang, D.; Lv, J.; Gao, B.; Liu, P. Identification of SNP markers correlated with the tolerance of low-salinity challenge in swimming crab (Portunus trituberculatus). Acta Oceanol. Sin. 2019, 38, 41–47. [Google Scholar] [CrossRef]
- Kindie, B. Assess molecular marker applications for genetic variety analysis in biodiversity conservation status. Mol. Biol. 2021, 10, 287. [Google Scholar]
- Gross, R.; Kõiv, K.; Pukk, L.; Kaldre, K. Development and characterization of novel tetranucleotide microsatellite markers in the noble crayfish (Astacus astacus) suitable for highly multiplexing and for detecting hybrids between the noble crayfish and narrow-clawed crayfish (A. leptodactylus). Aquaculture 2017, 472, 50–56. [Google Scholar] [CrossRef]
- Souza, F.; Ruas, C.; Urrea-Rojas, A.M.; Lima, E.; Povh, J.A.; Ribeiro, R.P.; Ruas, E.A.; Benicio, L.M.; Furlan-Murari, P.J.; Lopera-Barrero, N.M. Novel microsatellite markers for the invasive golden mussel Limnoperna fortunei. J. Shellfish Res. 2018, 37, 485–489. [Google Scholar] [CrossRef]
- Mondal, D.; Dutta, S.; Chakrabarty, U.; Mallik, A.; Mandal, N. Development and characterization of white spot disease linked microsatellite DNA markers in Penaeus monodon, and their application to determine the population diversity, cluster and structure. J. Invertebr. Pathol. 2019, 168, 107275. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, U.; Paulraj, R. Dietary protein requirement of giant mud crab Scylla serrata juveniles fed iso-energetic formulated diets having graded protein levels. Aquac. Res. 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Farhadi, A.; Fang, S.; Zhang, Y.; Cui, W.; Fang, H.; Ikhwanuddin, M.; Ma, H. The significant sex-biased expression pattern of Sp-Wnt4 provides novel insights into the ovarian development of mud crab (Scylla paramamosain). Int. J. Biol. Macromol. 2021, 183, 490–501. [Google Scholar] [CrossRef]
- Azra, M.N.; Ikhwanuddin, M. A review of maturation diets for mud crab genus Scylla broodstock: Present research, problems and future perspective. Saudi J. Biol. Sci. 2016, 23, 257–267. [Google Scholar] [CrossRef]
- Ma, H.Y.; Ma, C.Y.; Ma, L.B. Population genetic diversity of mud crab (Scylla paramamosain) in Hainan Island of China based on mitochondrial DNA. Biochem. Syst. Ecol. 2011, 39, 434–440. [Google Scholar] [CrossRef]
- Ma, C.; Ma, H.; Ma, L.; Jiang, K.; Zhang, F.; Song, W. Isolation and characterization of polymorphic microsatellite loci from cDNA library of Scylla paramamosain. Afr. J. Biotechnol. 2011, 10, 11142–11148. [Google Scholar]
- Ma, H.; Wu, Q.; Tan, H.; Wu, F.; Lin, F. Establishment of inter-specific hybridization technique and identification of phenotypic and genotypic characters of hybrids in mud crab (Scylla paramamosain and S. serrata). J. Shantou Univ. 2021, 36, 59–66. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F. Genepop’007 A Complete Re-Implementation of the Genepop Software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin Ver. 3.1: An Integrated Software Package for Population Genetics Data Analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; And Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Liu, F.; Yao, J.; Wang, X.; Repnikova, A.; Galanin, D.A.; Duan, D. Genetic diversity and structure within and between wild and cultivated Saccharina japonica (Laminariales, Phaeophyta) revealed by SSR markers. Aquaculture 2012, 358–359, 139–145. [Google Scholar] [CrossRef]
- Nayak, S.N.; Song, J.; Villa, A.; Pathak, B.; Ayala-Silva, T.; Yang, X.; Todd, J.; Glynn, N.C.; Kuhn, D.N.; Glaz, B.; et al. Promoting utilization of Saccharum spp. genetic resources through genetic diversity analysis and core collection construction. PLoS ONE 2014, 9, e110856. [Google Scholar] [CrossRef]
- Singh, R.B.; Singh, B.; Singh, R.K. Evaluation of genetic diversity in Saccharum species clones and commercial varieties employing molecular (SSRs) and physiological markers. Indian J. Plant. Genet. Resour. 2018, 31, 17–20. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–333. [Google Scholar] [PubMed]
- Cui, H.; Ma, Y.; Ma, C.; Qiao, Z.; Ma, Q.; Qi, L.; Lv, S.A.; Ma, L.B. Genetic diversity among different families of mud crab Scylla paramamosain by microsatellite markers. Mar. Fish 2011, 33, 274–281, (In Chinese with English Abstract). [Google Scholar]
- Ma, H.; Ma, C.; Li, X.; Xu, Z.; Feng, N.; Ma, L. The complete mitochondrial genome and gene organization of the mud crab (Scylla paramamosain) with phylogenetic consideration. Gene 2013, 519, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Hu, L.; Guo, L.; Zhang, J.; Tang, L.; Zhang, E.; Zhang, J.; Luo, S.; Tang, J.; Chen, X. Preservation of the genetic diversity of a local common carp in the agricultural heritage rice-fish system. Proc. Natl. Acad. Sci. USA 2018, 115, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Park, C.; Endo, T.; Kijima, A. Loss of genetic variation at microsatellite loci in hatchery strains of the Pacific abaione (Haliotis discus hannai). Aquaculture 2004, 235, 207–222. [Google Scholar] [CrossRef]
- Li, W.; Liu, L.; Wang, Y.; Zhang, Q.; Fan, G.; Zhang, S.; Wang, Y.; Liao, K. Genetic diversity, population structure, and relationships of apricot (Prunus) based on restriction site-associated DNA sequencing. Hortic. Res. 2020, 7, 69. [Google Scholar] [CrossRef]
- Kovach, R.P.; Gharrett, A.J.; Tallmon, D.A. Genetic change for earlier migration timing in a pink salmon population. Proc. R. Soc. B Biol. Sci. 2012, 279, 3870–3878. [Google Scholar] [CrossRef]
- Bi, X.; Yang, Q.; Gao, T.; Li, C. The loss of genetic diversity during captive breeding of the endangered sculpin, Trachidermus fasciatus, based on ISSR markers: Implications for its conservation. Chin. J. Oceanol. Limnol. 2011, 29, 958–966. [Google Scholar] [CrossRef]
- He, L.J.; Zhang, A.B.; Weese, D.; Zhu, C.D.; Jiang, C.J.; Qiao, Z.G. Late Pleistocene population expansion of Scylla paramamosain along the coast of China: A population dynamic response to the Last Interglacial sea level highstand. J. Exp. Mar. Biol. Ecol. 2010, 385, 20–28. [Google Scholar] [CrossRef]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequence of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Hill, W.G. Artificial selection. In Encyclopedia of Genetics; Academic Press: New York, NY, USA, 2001; pp. 96–101. [Google Scholar]
- Svardal, H.; Rueffler, C.; Hermisson, J. Comparing environmental and genetic variance as a response to fluctuating selection. Evolution 2011, 65, 2492–2513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Zhang, Y.; Ma, H.; Xiao, S.; Xiang, Z.; Yu, Z. Performance evaluation of reciprocal hybrids derived from the two brackish oysters, Crassostrea hongkongensis and Crassostrea sikamea in southern China. Aquaculture 2017, 473, 310–316. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Population | Code | Sample Size |
|---|---|---|
| Wild paternal S. paramamosain population | WSp | 36 |
| F1 paternal S. paramamosain family | FSp | 41 |
| Wild maternal S. serrata population | WSs | 40 |
| F1 maternal S. serrata family | FSs | 45 |
| F1 hybrid Scylla family (S. paramamosain ♂ × S. serrata ♀) | Hy | 39 |
| Locus | Repeat Motif | Primer Sequences (5′-3′) | Ta (°C) | Size (bp) | GenBank Accession No. |
|---|---|---|---|---|---|
| Scpa72 | (ATT)14 | F: TGCCGTCATTACTTCGTC | 53 °C | 240–390 | KC793964 |
| R: GAGAGTGCTTTGCATTATCA | |||||
| Scpa77 | (CA)13 | F: GTCTTTCAGGGTAGAGGTTATTA | 52 °C | 280–344 | KC793969 |
| R: CACAACAGATGAAGAACGCT | |||||
| Scpa02 | (CCA)3A(CGC)3N4(ACC)4N94(CT)27 | F: CAGCAGCTTCACCGCCAATA | 60 °C | 262–336 | GU182882 |
| R: TCAAAGAAAGAGGGACGGGAGA | |||||
| Scpa51 | (GT)8A(TG)6TAC(GT)7 | F: AAGTACCACCAATAAATATGAATAA | 46 °C | 218–242 | KC793943 |
| R: TGGAAAAAATAACATAAATAAAAGT | |||||
| Scpa07 | (CT)3(TCCT)6N49(AC)17 | F: CAGACCCTGAATTTACCTGT | 49 °C | 170–216 | GU18288 |
| R: TAGAATAAGAGTCAAAGAAAGCT | |||||
| Scpa37 | (TG)17 | F: AGATGAGATCGCCAAGGGT | 53 °C | 190–218 | KC793929 |
| R: CTGACCTGATTGTGTAACCCA | |||||
| Scpa33 | (CA)13 | F: GTTCCTGCGTGGGTGCTC | 57 °C | 190–274 | KC793925 |
| R: ACATCTAATCCAACCATGCGTAC | |||||
| Scpa24 | (AGT)12 | F: TAGGGCTGAAATGTATGAAA | 52 °C | 275–332 | KC793916 |
| R: AATGAGGCTAAGAATGTGAAT | |||||
| Scpa23 | (TAG)10 | F: AAGCACCAAGAGGAGAGT R: CTCATGTCTCGCTACACC | 54 °C | 218–272 | KC793915 |
| Locus | Na | Ne | Ho | He | PIC |
|---|---|---|---|---|---|
| Scpa72 | 15 | 8.008 | 0.840 | 0.875 | 0.863 |
| Scpa77 | 13 | 7.894 | 0.927 | 0.873 | 0.861 |
| Scpa02 | 9 | 4.587 | 0.870 | 0.782 | 0.753 |
| Scpa51 | 9 | 5.071 | 0.725 | 0.803 | 0.774 |
| Scpa07 | 10 | 5.031 | 0.920 | 0.801 | 0.778 |
| Scpa37 | 9 | 4.154 | 0.738 | 0.759 | 0.726 |
| Scpa33 | 14 | 8.053 | 0.838 | 0.876 | 0.865 |
| Scpa24 | 6 | 4.001 | 0.405 | 0.750 | 0.706 |
| Scpa23 | 13 | 7.001 | 0.737 | 0.857 | 0.842 |
| Mean | 10.889 | 5.978 | 0.778 | 0.819 | 0.796 |
| Locus | Na | Ne | Ho | He | PIC | HWE | Fis | Fua |
|---|---|---|---|---|---|---|---|---|
| WSp | ||||||||
| Scpa72 | 9 | 6.964 | 0.853 | 0.856 | 0.840 | 0.192 | 0.019 | 0.002 |
| Scpa77 | 6 | 5.312 | 0.879 | 0.812 | 0.785 | 0.631 | −0.067 | −0.037 |
| Scpa02 | 7 | 5.642 | 0.813 | 0.823 | 0.799 | 0.108 | 0.028 | 0.006 |
| Scpa51 | 6 | 3.580 | 0.563 | 0.721 | 0.671 | 0.032 | 0.235 | 0.092 |
| Scpa07 | 7 | 4.734 | 0.871 | 0.789 | 0.764 | 0.119 | −0.088 | −0.046 |
| Scpa37 | 6 | 4.253 | 0.800 | 0.765 | 0.729 | 0.074 | −0.031 | −0.020 |
| Scpa33 | 8 | 6.135 | 0.879 | 0.837 | 0.818 | 0.013 | −0.035 | −0.023 |
| Scpa24 | 6 | 4.063 | 0.697 | 0.754 | 0.718 | 0.044 | 0.091 | 0.033 |
| Scpa23 | 8 | 6.205 | 0.758 | 0.839 | 0.819 | 0.084 | 0.112 | 0.044 |
| FSp | ||||||||
| Scpa72 | 4 | 3.081 | 0.576 | 0.675 | 0.628 | 0.003 * | 0.163 | 0.060 |
| Scpa77 | 4 | 3.129 | 0.946 | 0.680 | 0.620 | 0.000 * | −0.379 | −0.158 |
| Scpa02 | 4 | 3.906 | 0.730 | 0.744 | 0.696 | 0.138 | 0.033 | 0.008 |
| Scpa51 | 3 | 2.652 | 0.795 | 0.623 | 0.552 | 0.009 | −0.264 | −0.106 |
| Scpa07 | 4 | 3.726 | 1.000 | 0.732 | 0.682 | 0.000 * | −0.355 | −0.155 |
| Scpa37 | 3 | 2.490 | 0.585 | 0.598 | 0.513 | 0.197 | 0.034 | 0.008 |
| Scpa33 | 4 | 3.497 | 0.865 | 0.714 | 0.661 | 0.000 * | −0.198 | −0.088 |
| Scpa24 | 2 | 1.730 | 0.395 | 0.422 | 0.333 | 0.702 | 0.078 | 0.019 |
| Scpa23 | 4 | 3.723 | 0.902 | 0.731 | 0.681 | 0.076 | −0.222 | −0.099 |
| WSs | ||||||||
| Scpa72 | 5 | 3.941 | 0.850 | 0.746 | 0.703 | 0.046 | −0.127 | −0.059 |
| Scpa77 | 6 | 2.999 | 0.868 | 0.667 | 0.616 | 0.0013 * | −0.291 | −0.121 |
| Scpa02 | 5 | 2.629 | 0.795 | 0.620 | 0.578 | 0.407 | −0.271 | −0.108 |
| Scpa51 | 3 | 2.027 | 0.632 | 0.507 | 0.421 | 0.348 | −0.234 | −0.083 |
| Scpa07 | 5 | 3.146 | 0.763 | 0.682 | 0.640 | 0.013 | −0.106 | −0.048 |
| Scpa37 | 4 | 1.994 | 0.568 | 0.499 | 0.419 | 0.579 | −0.125 | −0.046 |
| Scpa33 | 4 | 2.373 | 0.730 | 0.579 | 0.528 | 0.146 | −0.249 | −0.096 |
| Scpa24 | 2 | 1.651 | 0.324 | 0.394 | 0.317 | 0.400 | 0.191 | 0.050 |
| Scpa23 | 3 | 2.880 | 0.472 | 0.653 | 0.579 | 0.111 | 0.290 | 0.109 |
| FSs | ||||||||
| Scpa72 | 3 | 2.667 | 1.000 | 0.625 | 0.555 | 0.000 * | −0.593 | −0.231 |
| Scpa77 | 3 | 2.113 | 0.933 | 0.527 | 0.419 | 0.000 * | −0.768 | −0.266 |
| Scpa02 | 4 | 2.664 | 1.000 | 0.625 | 0.556 | 0.000 * | −0.593 | −0.231 |
| Scpa51 | 4 | 2.980 | 0.750 | 0.664 | 0.602 | 0.000 * | −0.116 | −0.051 |
| Scpa07 | 3 | 2.559 | 0.953 | 0.609 | 0.527 | 0.000 * | −0.557 | −0.214 |
| Scpa37 | 3 | 2.608 | 0.775 | 0.617 | 0.546 | 0.042 | −0.245 | −0.098 |
| Scpa33 | 4 | 2.738 | 0.800 | 0.635 | 0.580 | 0.000 * | −0.250 | −0.101 |
| Scpa24 | 2 | 1.953 | 0.489 | 0.488 | 0.369 | 1.000 | 0.009 | −0.001 |
| Scpa23 | 2 | 1.939 | 0.556 | 0.484 | 0.367 | 0.536 | −0.136 | −0.048 |
| Hy | ||||||||
| Scpa72 | 3 | 2.947 | 0.865 | 0.661 | 0.587 | 0.017 | −0.297 | −0.123 |
| Scpa77 | 4 | 3.851 | 1.000 | 0.740 | 0.692 | 0.000 * | −0.339 | −0.149 |
| Scpa02 | 3 | 2.992 | 1.000 | 0.666 | 0.592 | 0.000 * | −0.492 | −0.201 |
| Scpa51 | 4 | 3.896 | 0.879 | 0.743 | 0.695 | 0.000 * | −0.167 | −0.078 |
| Scpa07 | 4 | 3.951 | 1.000 | 0.747 | 0.700 | 0.000 * | −0.327 | −0.145 |
| Scpa37 | 4 | 3.682 | 1.000 | 0.728 | 0.679 | 0.000 * | −0.360 | −0.157 |
| Scpa33 | 3 | 2.504 | 0.923 | 0.601 | 0.532 | 0.000 * | −0.528 | −0.202 |
| Scpa24 | 2 | 1.911 | 0.135 | 0.477 | 0.363 | 0.000 * | 0.723 | 0.231 |
| Scpa23 | 3 | 2.638 | 1.000 | 0.621 | 0.549 | 0.000 * | −0.602 | −0.234 |
| Mean | ||||||||
| WSp | 7.000 | 5.210 | 0.790 | 0.799 | 0.771 | 0.144 | 0.029 | 0.006 |
| FSp | 3.556 | 3.104 | 0.755 | 0.658 | 0.596 | 0.125 | −0.123 | −0.057 |
| WSs | 4.111 | 2.627 | 0.667 | 0.594 | 0.533 | 0.228 | −0.102 | −0.045 |
| FSs | 3.111 | 2.469 | 0.806 | 0.586 | 0.502 | 0.175 | −0.361 | −0.138 |
| Hy | 3.333 | 3.152 | 0.867 | 0.665 | 0.599 | 0.002 | −0.265 | −0.117 |
| Population | WSp | FSp | WSs | FSs | Hy |
|---|---|---|---|---|---|
| WSp | 0.000 | 0.000 | 0.000 | 0.000 | |
| FSp | 0.179 | 0.000 | 0.000 | 0.000 | |
| WSs | 0.175 | 0.291 | 0.000 | 0.000 | |
| FSs | 0.223 | 0.362 | 0.094 | 0.000 | |
| Hy | 0.247 | 0.257 | 0.337 | 0.386 |
| Source of Variation | Variance Components | Percentage of Variation (%) | Fixation Index | p Value |
|---|---|---|---|---|
| Among populations | 0.34 | 26.72 | Φst = 0.26716 | 0.00000 |
| Within populations | 0.93 | 73.28 |
| Population | WSp | FSp | WSs | FSs | Hy |
|---|---|---|---|---|---|
| WSp | 0.480 | 0.374 | 0.447 | 0.370 | |
| FSp | 0.734 | 0.344 | 0.234 | 0.543 | |
| WSs | 0.985 | 1.068 | 0.849 | 0.214 | |
| FSs | 0.805 | 1.453 | 0.164 | 0.160 | |
| Hy | 0.995 | 0.611 | 1.540 | 1.835 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, W.; Cui, W.; Wu, F.; Chen, H.; Liu, S.; Guan, M.; Saqib, H.S.A.; Ye, S.; Ikhwanuddin, M.; Ma, H. Genetic Diversity and Differences among Three F1 Families and Two Wild Populations of Genus Scylla Using Microsatellite Markers. Fishes 2023, 8, 18. https://doi.org/10.3390/fishes8010018
Gao W, Cui W, Wu F, Chen H, Liu S, Guan M, Saqib HSA, Ye S, Ikhwanuddin M, Ma H. Genetic Diversity and Differences among Three F1 Families and Two Wild Populations of Genus Scylla Using Microsatellite Markers. Fishes. 2023; 8(1):18. https://doi.org/10.3390/fishes8010018
Chicago/Turabian StyleGao, Weifeng, Wenxiao Cui, Fangchun Wu, Huiying Chen, Simin Liu, Mengyun Guan, Hafiz Sohaib Ahmed Saqib, Shaopan Ye, Mhd Ikhwanuddin, and Hongyu Ma. 2023. "Genetic Diversity and Differences among Three F1 Families and Two Wild Populations of Genus Scylla Using Microsatellite Markers" Fishes 8, no. 1: 18. https://doi.org/10.3390/fishes8010018
APA StyleGao, W., Cui, W., Wu, F., Chen, H., Liu, S., Guan, M., Saqib, H. S. A., Ye, S., Ikhwanuddin, M., & Ma, H. (2023). Genetic Diversity and Differences among Three F1 Families and Two Wild Populations of Genus Scylla Using Microsatellite Markers. Fishes, 8(1), 18. https://doi.org/10.3390/fishes8010018