
Improving Growth, Digestive and Antioxidant Enzymes and Immune Response of Juvenile Grass Carp (Ctenopharyngodon idella) by Using Dietary Spirulina platensis

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Culture and Diet Preparation
2.2. Experimental Setup
2.3. Analysis of Growth
2.4. Analysis of Mucus Immunity
2.5. Oxidative Stress and Antioxidant Defense Markers Assessment
2.6. Analysis of Digestive Enzymes
2.7. Expression Analysis of Immune-Related Genes
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Mucus Immunity
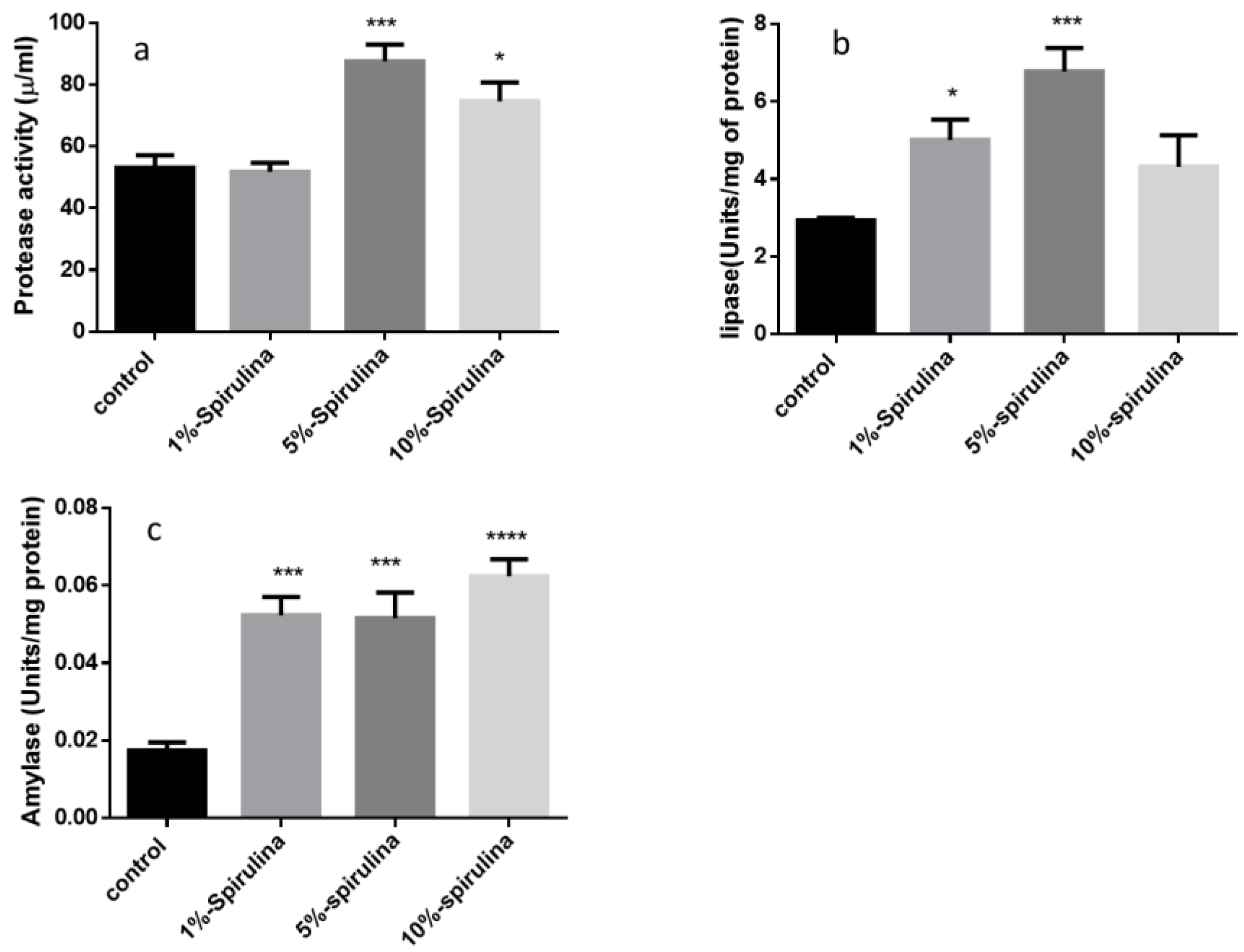
3.3. Digestive Enzyme Activity
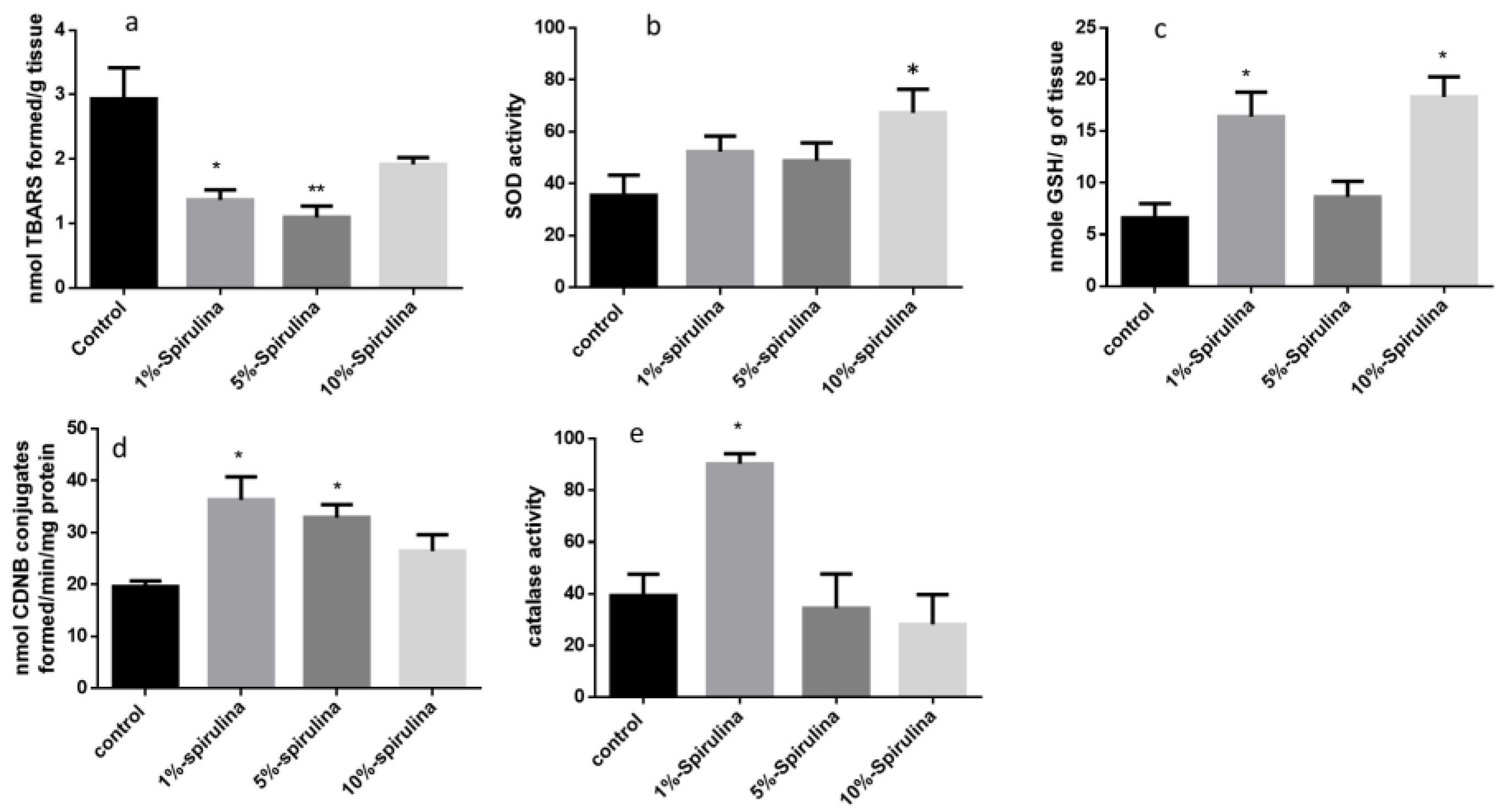
3.4. Hepatic Anti-Oxidants
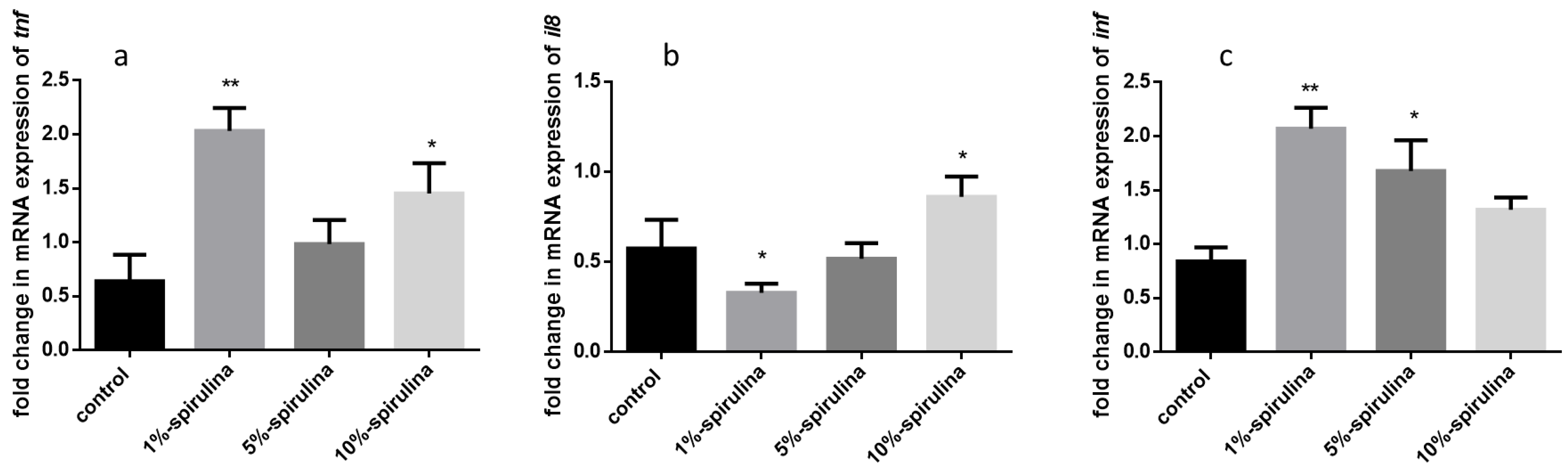
3.5. Gene Expression
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoseinifar, S.H.; Sun, Y.-Z.; Zhou, Z.; Van Doan, H.; Davies, S.J.; Harikrishnan, R. Boosting immune function and disease bio-control through environment- friendly and sustainable approaches in finfish aquaculture: Herbal therapy scenarios. Rev. Fish. Sci. Aquac. 2020, 28, 303–321. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar]
- Pradeepkiran, J.A. Aquaculture role in global food security with nutritional value: A review. Transl. Anim. Sci. 2019, 3, 903–910. [Google Scholar] [PubMed]
- FAO—Food and Agricultural Organization. The State of World Fisheries and Aquaculture 2018-Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Al-Deriny, S.H.; Dawood, M.A.O.; Abou Zaid, A.; El-Trasa, W.F.; Paray, B.A.; Van Doan, H.; Mohamed, R.A. The synergistic effects of Spirulina platensis and Bacillus amyloliquefaciens on the growth performance, intestinal histomorphology, and immune response of Nile tilapia (Oreochromis niloticus). Aquac. Rep. 2020, 17, 100390. [Google Scholar]
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [PubMed]
- Bosila, M.A.; Mekky, H.M.; Fedawy, H.S.; Elbayomi, K.M.; Amer, M.M. Histopathological lesion of arthritis in Mycoplasma synoviae naturally infected breeder chicken in Egypt. Int. J. Vet. Sci 2021, 10, 72–74. [Google Scholar]
- Saeed, N.M. Sequence analysis and comparison of infectious bursal disease virus affecting indigenous Kurdish breed and broiler chickens in Sulaymaniyah, Kurdistan Region of Iraq. Pak. Vet. J. 2021, 41, 249–253. [Google Scholar]
- Tahir, A.; Khan, M.A.; Bibi, K.; Bibi, S.; Rauf, F.; Ayaz, F. Prevalence of Colibacillosis in Young Broiler Chicks and Antibiogram of Escherichia coli in Different Areas of Hazara Region. Adv. Life Sci. 2021, 8, 238–240. [Google Scholar]
- Abu, S.M.; Monira, N.; Asmaul, H.; E-Elahi ATM Mahbub, A.T.M.; Mustafa, K.A.H.; Masudur, R.M. Seroprevalence of Newcastle disease in layer chickens and pathology in clinically affected chickens at Gazipur, Bangladesh. Cont. Vet. J. 2022, 2, 35–41. [Google Scholar]
- Ul-Rahman, A.; Shabbir, M.A.B.; Mehmood, A.; Shabbir, M.Z. Genotypic and subgenotypic diversity of avian paramyxoviruses 2, 4 and 6. Pak. Vet. J. 2021, 41, 156–159. [Google Scholar]
- Elazab, S.T.; Elshater, N.S.; Hashem, Y.H.; Abdelaziz, A.S. Pharmacokinetics of tildipirosin in healthy and Mycoplasma gallisepticum infected chickens. Int. J. Vet. Sci. 2021, 10, 119–123. [Google Scholar]
- Iraqi, M.; Nasef, S.A.; El-Enbaawy, M. Phenotypic and genotypic characteristics of antimicrobial and disinfectant resistance gram-negative bacteria involved in early broiler chick mortality. Int. J. Vet. Sci. 2021, 10, 129–134. [Google Scholar]
- Sharif, M.; Tunio, S.A.; Bano, S. Synergistic effects of Zinc oxide nanoparticles and conventional antibiotics against methicillin resistant Staphylococcus aureus. Adv. Life Sci. 2021, 8, 167–171. [Google Scholar]
- Betelhem, T.; Shubisa, A.L.; Bari, F.D. Isolation, identification and antimicrobial resistance of Staphylococcus aureus isolates from mastitis cases of lactating dairy cows found in Sululta and Holleta Towns, Oromia, Ethiopia. Agrobiol. Rec. 2022, 8, 27–34. [Google Scholar]
- Al-Sarraj, F.M.B. A Review on the impacts of Azadirachta indica on Multi-drug Resistant Extended Spectrum Beta Lactamase positive of Escherichia coli and Klebsiella pneumonia. Adv. Life Sci. 2021, 8, 228–232. [Google Scholar]
- Mohamed, H.F.; Atta, A.H.; Darwish, A.S.; Atef, M. Effect of probiotics on the pharmacokinetic aspects and tissue residues of difloxacin in broiler chickens. Pak. Vet. J. 2021, 41, 269–273. [Google Scholar]
- Mohsin, M.; Li, L.; Huang, X.; Aleem, M.T.; Habib, Y.J.; Shehata, A.I.; Afzal, M.Z.; Abbas, R.Z.; Abbas, A.; Yin, G. Immunogenicity and protective efficacy of probiotics with EtIMP1C against Eimeria tenella challenge. Pak. Vet. J. 2021, 41, 274–278. [Google Scholar]
- Rafay, M.; Ghaffar, M.U.; Abid, M.; Malik, Z.; Madnee, M. Phytochemicals analysis and antimicrobial activities of chinops echinatus from Cholistan desert, Pakistan. Agrobiol. Rec. 2021, 5, 21–27. [Google Scholar]
- Rani, Z.; Abbas, R.Z.; Abbas, A.; Saeed, Z.; Rehman, T.; Mehmood, K.; Rehman, A.; Hussain, K. In vitro and in vivo anticoccidial effects of butyric acid and its impact on blood and serum chemistry of broiler chickens. Kafkas. Univ. Vet. Fak. Derg. 2021, 27, 583–588. [Google Scholar]
- Rehman, K.; Hamayun, M.; Khan, S.S.; Ahmad, W.S. Efficiency of virgin’s mantle (Fagonia cretica L.) as an antibacterial and antifungal agent. Adv. Life Sci. 2021, 8, 233–237. [Google Scholar]
- Ma, K.; Bao, Q.; Wu, Y.; Chen, S.; Zhao, S.; Wu, H.; Fan, J. Evaluation of Microalgae as Immunostimulants and Recombinant Vaccines for Diseases Prevention and Control in Aquaculture. Front. Bioeng. Biotechnol. 2020, 8, 590431. [Google Scholar] [PubMed]
- Ahmadifar, E.; Fallah, H.P.; Yousefi, M.; Dawood, M.A.O.; Hoseinifar, S.H.; Adineh, H.; Yilmaz, S.; Paolucci, M.; Van Doan, V. The gene regulatory roles of herbal extracts on the growth, immune system, and reproduction of fish. Animals 2021, 11, 2167. [Google Scholar]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Impact of plant products on innate and adaptive immune system of cultured finfish and shellfish. Aquaculture 2011, 317, 1–15. [Google Scholar]
- Harikrishnan, R.; Kim, J.S.; Kim, M.C.; Balasundaram, C.; Heo, M.S. Lactuca indica extract as feed additive enhances immunological parameters and disease resistance in Epinephelus bruneus to Streptococcus iniae. Aquaculture 2011, 318, 43–47. [Google Scholar]
- Van Doan, H.; Hoseinifar, S.H.; Chitmanat, C.; Jaturasitha, S.; Paolucci, M.; Ashouri, G.; Dawood, M.A.O.; Esteban, M.A. The effects of Thai ginseng, Boesenbergia rotunda powder on mucosal and serum immunity, disease resistance, and growth performance of Nile tilapia (Oreochromis niloticus) fingerlings. Aquaculture 2019, 513, 734388. [Google Scholar]
- Van Doan, H.; Lumsangkul, C.; Hoseinifar, S.H.; Hung, T.Q.; Stejskal, V.; Ringø, E.; Dawood, M.A.O.; Esteban, M.A. Administration of watermelon rind powder to Nile tilapia (Oreochromis niloticus) culture under biofloc system: Effect on growth performance, innate immune response, and disease resistance. Aquaculture 2020, 528, 735574. [Google Scholar]
- Mohammadi, G.; Rafiee, G.; El Basuini, M.F.; Van Doan, H.; Ahmed, H.A.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Oregano (Origanum vulgare), St John’s-wort (Hypericum perforatum), and lemon balm (Melissa officinalis) extracts improved the growth rate, antioxidative, and immunological responses in Nile tilapia (Oreochromis niloticus) infected with Aeromonas hydrophila. Aquac. Rep. 2020, 18, 100445. [Google Scholar]
- Abdel-Tawwab, M.; El-Araby, D.A. Immune and antioxidative effects of dietary licorice (Glycyrrhiza glabra L.) on performance of Nile tilapia, Oreochromis niloticus (L.) and its susceptibility to Aeromonas hydrophila infection. Aquaculture 2021, 530, 735828. [Google Scholar]
- Stratev, D.; Zhelyazkov, G.; Noundou, X.S.; Krause, R.W.M. Beneficial effects of medicinal plants in fish diseases. Aquac. Int. 2018, 26, 289–308. [Google Scholar]
- Bilen, S.; Altunoglu, Y.C.; Ulu, F.; Biswas, G. Innate immune and growth promoting responses to caper (Capparis spinosa) extract in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016, 57, 206–212. [Google Scholar]
- Van Doan, H.; Hoseinifar, S.H.; Naraballobh, W.; Jaturasitha, S.; Tongsiri, S.; Chitmanat, C.; Ringø, E. Dietary inclusion of orange peels derived pectin and Lactobacillus plantarum for Nile tilapia (Oreochromis niloticus) cultured under indoor biofloc systems. Aquaculture 2019, 508, 98–105. [Google Scholar]
- Srichaiyo, N.; Tongsiri, S.; Hoseinifar, S.H.; Dawood, M.A.O.; Jaturasitha, S.; Esteban, M.A.; Ringø, E.; Van Doan, H. The effects gotu kola (Centella asiatica) powder on growth performance, skin mucus, and serum immunity of Nile tilapia (Oreochromis niloticus) fingerlings. Aquac. Rep. 2020, 16, 100239. [Google Scholar]
- Gabriel, N.N.; Qiang, J.; Ma, X.Y.; He, J.; Xu, P.; Liu, K. Dietary Aloe vera improves plasma lipid profile, antioxidant, and hepatoprotective enzyme activities in GIFT-tilapia (Oreochromis niloticus) after Streptococcus iniae challenge. Fish Physiol. Biochem. 2015, 41, 1321–1332. [Google Scholar] [PubMed]
- Kuebutornye, F.K.; Abarike, E.D. The contribution of medicinal plants to tilapia aquaculture: A review. Aquac. Int. 2020, 28, 965–983. [Google Scholar]
- Sonmez, A.Y.; Bilen, S.; Alak, G.; Hisar, O.; Yanık, T.; Biswas, G. Growth performance and antioxidant enzyme activities in rainbow trout (Oncorhynchus mykiss) juveniles fed diets supplemented with sage, mint and thyme oils. Fish Physiol. Biochem. 2015, 41, 165–175. [Google Scholar] [PubMed]
- Awed, E.M.; Sadek, K.M.; Soliman, M.K.; Khalil, R.H.; Younis, E.M.; Abdel-Warith, A.A.-W.; Van Doan, H.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Spirulina platensis Malleviated the oxidative damage in the gills, liver, and kidney organs of Nile tilapia intoxicated with sodium sulphate. Animals 2020, 10, 2423. [Google Scholar]
- Yousefi, M.; Abtahi, B.; Adineh, H.; Hoseinifar, S.H.; Mirghaed, A.T.; Paolucci, M.; Van Doan, H. Effects of dietary arginine supplementation on cytokine- and antioxidant-related gene expressions in common carp (Cyprinus carpio) fingerling during ammonia toxicity. Aquac. Res. 2021, 52, 2751–2758. [Google Scholar]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2010, 18, 403–414. [Google Scholar]
- Lee, J.Y.; Gao, Y. Review of the application of garlic, Allium sativum, in aquaculture. J. World Aquac. Soc. 2012, 43, 447–458. [Google Scholar]
- Xu, A.; Shang-Guan, J.; Li, Z.; Gao, Z.; Huang, Y.C.; Chen, Q. Effects of dietary Chinese herbal medicines mixture on feeding attraction activity, growth performance, nonspecific immunity and digestive enzyme activity of Japanese seabass (Lateolabrax japonicus). Aquac. Rep. 2020, 17, 100304. [Google Scholar]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar]
- Egamberdieva, D.; Wirth, S.; Behrendt, U.; Ahmad, P.; Berg, G. Antimicrobial activity of medicinal plants correlates with the proportion of antagonistic endophytes. Front. Microbiol. 2017, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Tang, H.; Lyles, J.T.; Pineau, R.; Mashwani, Z.-U.-R.; Quave, C.L. Antibacterial properties of medicinal plants from Pakistan against multidrug-resistant ESKAPE pathogens. Front. Pharmacol. 2018, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Dada, A.A. Improvement of tilapia (Oreochromis niloticus Linnaeus, 1758) growth performance fed three commercial feed additives in diets. J. Aquac. Res. Dev. 2015, 6, 2. [Google Scholar]
- Radhakrishnan, S.; Bhavan, P.S.; Seenivasan, C.; Shanthi, R.; Muralisankar, T. Replacement of fishmeal with Spirulina platensis, Chlorella vulgaris and Azolla pinnata on non-enzymatic and enzymatic antioxidant activities of Macrobrachium rosenbergii. J. Basic Appl. Zool. 2014, 67, 25–33. [Google Scholar]
- Sheikhzadeh, N.; Mousavi, S.; Oushani, A.K.; Firouzamandi, M.; Mardani, K. Spirulina platensis in rainbow trout (Oncorhynchus mykiss) feed: Effects on growth, fillet composition, and tissue antioxidant mechanisms. Aquac. Int. 2019, 27, 1613–1623. [Google Scholar]
- Raji, A.A.; Jimoh, W.A.; Abu Bakar, N.H.; Mohd Taufek, N.H.; Muin, H.; Alias, Z.; Milow, P.; Abdul Razak, S. Dietary use of Spirulina (Arthrospira) and Chlorella instead of fish meal on growth and digestibility of nutrients, amino acids and fatty acids by African catfish. J. Appl. Phycol. 2020, 32, 1763–1770. [Google Scholar]
- Xia, Y.; Liu, C.; Fei, S.; Liu, H.; Han, D.; Jin, J.; Yang, Y.; Zhu, X.; Xie, S. Arthrospira platensis additive enhances the growth performance and antioxidant response in hybrid yellow catfish (Pelteobagrus fulvidraco♀ × Pelteobagrus vachelli♂). Aquac. Rep. 2021, 20, 100721. [Google Scholar]
- Ragap, H.M.; Khalil, R.H.; Mutawie, H. Immunostimulant effects of dietary Spirulina platensis on tilapia Oreochromis niloticus. J. App. Pharm. Sci. 2012, 2, 26–31. [Google Scholar]
- Watanuki, H.; Ota, K.; Tassakka, A.C.M.A.; Kato, T.; Sakai, M. Immunostimulant effects of dietary Spirulina platensis on carp, Cyprinus carpio. Aquaculture 2006, 258, 157–163. [Google Scholar]
- Adel, M.; Yeganeh, S.; Dadar, M.; Sakai, M.; Dawood, M.A.O. Effects of dietary Spirulina platensis on growth performance, humoral and mucosal immune responses and disease resistance in juvenile great sturgeon (Huso huso Linnaeus, 1754). Fish Shellfish Immuno. 2016, 56, 436–444. [Google Scholar]
- Teimouri, M.; Yeganeh, S.; Mianji, G.R.; Najafi, M.; Mahjoub, S. The effect of Spirulina platensis meal on antioxidant gene expression, total antioxidant capacity, and lipid peroxidation of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2019, 45, 977–986. [Google Scholar] [CrossRef]
- Yu, W.; Wen, G.; Lin, H.; Yang, Y.; Huang, X.; Zhou, C.; Zhang, Z.; Duan, Y.; Huang, Z.; Li, T. Effects of dietary Spirulina platensis on growth performance, hematological and serum biochemical parameters, hepatic antioxidant status, immune responses and disease resistance of Coral trout Plectropomus leopardus (Lacepede, 1802). Fish Shellfish Immunol. 2018, 74, 649–655. [Google Scholar] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Latif, M.; Faheem, M.; Asmatullah. Study of oxidative stress and histo-biochemical biomarkers of diethyl phthalate induced toxicity in a culturable fish, Labeo rohita. Pak. Vet. J. 2020, 40, 202–208. [Google Scholar] [CrossRef]
- Faheem, M.; Khaliq, S.; Mustafa, N.; Rani, S.; Lone, K.P. Dietary Moringa oleferia leaf meal induce growth, innate immunity and cytokine expression in grass carp, Ctenopharyngodon Idella. Aquac. Nut. 2020, 26, 1164–1172. [Google Scholar]
- Faheem, M.; Lone., K.P. Oxidative stress and histopathologic biomarkers of exposure to bisphenol-A in the freshwater fish, Ctenopharyngodon idella. Braz. J. Pharm. Sci. 2017, 53, e17003. [Google Scholar]
- Faheem, M.; Zahid, Z.; Ferreira, N.G.C. Toxicity assessment of dibutyl phthalate in grass carp: An integrated biomarker approach. Pak. Vet. J. 2021, 41, 365–371. [Google Scholar]
- Allameh, S.K.; Ringø, E.; Yusoff, F.M.; Daud, H.M.; Ideris, A. Dietary supplement of Enterococcus faecalis on digestive enzyme activities, short-chain fatty acid production, immune system response and disease resistance of Javanese carp (Puntius gonionotus, Bleeker 1850). Aquac. Nutr. 2017, 23, 331–338. [Google Scholar]
- Dar, S.A.; Srivastava, P.P.; Nazir, M.I.; Jahan, I.; Varghese, T.; Hajam, I.A.; Gupta, S. Modulations of digestive and metabolic enzymes profiles during restriction feeding in Labeo rohita (Hamilton, 1822) fingerlings. N. Am. J. Aquac. 2021, 83, 58–66. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalil, M.; Tabarraei, A.; Van Doan, V.H.; Span, N.; Faggio, C. Mucosal immune parameters, immune and antioxidant defence related genes expression and growth performance of zebrafish (Danio rerio) fed on Gracilaria gracilis powder. Fish Shellfish Immunol. 2018, 83, 232–237. [Google Scholar] [PubMed]
- Van Doan, H.; Hoseinifar, S.H.; Sringarm, K.; Jaturasitha, S.; Khamlor, T.; Dawood, M.A.O.; Esteban, M.A.; Soltani, M.; Mustha, M.S. Effects of elephant’s foot (Elephantopus scaber) extract on growth performance, immune response, and disease resistance of Nile tilapia (Oreochromis niloticus) fingerlings. Fish Shellfish Immunol. 2019, 93, 328–335. [Google Scholar]
- Harikrishnan, R.; Devi, G.; Van Doan, H.V.; Balasundaram, C.; Thamizharasan, S.; Hoseinifar, S.H.; Abdel-Tawwab, M. Effect of diet enriched with Agaricus bisporus polysaccharides (ABPs) on antioxidant property, innate-adaptive immune response and pro-anti inflammatory genes expression in Ctenopharyngodon idella against Aeromonas hydrophila. Fish Shellfish Immunol. 2021, 114, 238–252. [Google Scholar] [PubMed]
- Ghafarifarsani, H.; Rashidian, G.; Sheikhlar, A.; Farsani, M.N.; Hoseinifar, S.H.; Van Doan, H.V. The use of dietary oak acorn extract to improve haematological parameters, mucosal and serum immunity, skin mucus bactericidal activity, and disease resistance in rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2021, 52, 2518–2527. [Google Scholar]
- Rufchaei, R.; Nedaei, S.; Hoseinifar, S.H.; Hassanpour, S.; Golshan, M.; Bourani, M.S. Improved growth performance, serum and mucosal immunity, haematology and antioxidant capacity in pikeperch (Sander lucioperca) using dietary water hyacinth (Eichhornia crassipes) leaf powder. Aquac. Res. 2021, 52, 2194–2204. [Google Scholar]
- Latif, M.; Faheem, M.; Asmatullah; Hoseinifar, S.H.; Van Doan, H. Protective efficacy of Nigella sativa seeds against diethyl phthalate induced growth retardation, oxidative stress and histo-biochemical damages in Labeo rohita. Aquaculture 2021, 533, 736065. [Google Scholar]
- Akintunde, E.A. Digestive enzymes in the digestive enzymes in the gut of Sarotherodon galilaeus (syn. Tilapia galilaea, family cichlidae) of the Lake Kainji, Nigeria. Niger. J. Sci. 1985, 18, 22–25. [Google Scholar]
- Guzman, C.; Gaxiola, G.; Rosa, C.; Torre-Blanco, A. The effect of dietary protein and total energy content on digestive enzyme activities, growth and survival of Litopenaeus setiferus post larvae. Aquac. Nut. 2005, 7, 113–122. [Google Scholar]
- Hoseinifar, S.H.; Dadar, M.; Khalil, M.; Cerezuela, R.; Esteban, M.Á. Effect of dietary supplementation of palm fruit extracts on the transcriptomes of growth, antioxidant enzyme and immune-related genes in common carp (Cyprinus carpio) fingerlings. Aquac. Res. 2017, 48, 3684–3692. [Google Scholar]
- Takeuchi, T.; Lu, J.; Yoshizaki, G.; Satoh, S. Effect on the growth and body composition of juvenile tilapia Oreochromis niloticus fed raw Spirulina. Fish Sci. 2002, 68, 34–40. [Google Scholar]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar]
- Esteban, M.A. An overview of the immunological defenses in fish skin. ISRN Immunol. 2012, 2012, 853470. [Google Scholar] [CrossRef]
- Renugadevi, K.; Valli Nachiyar, C.; Sowmiya, P.; Sunkar, S. Antioxidant activity of phycocyanin pigment extracted from marine filamentous cyanobacteria Geitlerinema sp TRV57. Biocatal. Agric. Biotechnol. 2018, 16, 237–242. [Google Scholar]
- Promya, J.; Chitmanat, C. The effects of Spirulina platensis and Cladophora algae on the growth performance, meat quality and immunity stimulating capacity of the African sharptooth catfish (Clarias gariepinus). Int. J. Agric. Biol. 2011, 13, 77–82. [Google Scholar]
- Mahmoud, M.M.; El-Lamie, M.M.; Kilany, O.E.; Dessouki, A.A. Spirulina (Arthrospira platensis) supplementation improves growth performance, feed utilization, immune response, and relieves oxidative stress in Nile tilapia (Oreochromis niloticus) challenged with Pseudomonas fluorescens. Fish Shellfish Immunol. 2018, 72, 291–300. [Google Scholar] [PubMed]
- Wink, M. Evolution of toxins and antinutritional factors in plants with special emphasis on Leguminosae. In Poisonous Plants and Related Toxins; CABI Publishing: Oxford, UK, 2004; pp. 1–25. [Google Scholar]
- Galindo-Villegas, J.; Mulero, I.; García-Alcazar, A.; Muñoz, I.; Peñalver-Mellado, M.; Streitenberger, S.; Scapigliati, G.; Meseguer, J.; Mulero, V. Recombinant TNFα as oral vaccine adjuvant protects European sea bass against vibriosis: Insights into the role of the CCL25/CCR9 axis. Fish Shellfish Immunol. 2013, 35, 1260–1271. [Google Scholar]
- Abdellatief, S.A.; Abdel Rahman, A.N.; Abdallah, F.D. Evaluation of Immunostimulant Activity of Spirulina platensis (Arthrospira platensis) and Sage (Salvia officinalis) in Nile Tilapia (Oreochromis niloticus). Zagazig Vet. J. 2018, 46, 25–36. [Google Scholar]
- Shih, C.M.; Cheng, S.N.; Wong, C.S.; Kuo, Y.L.; Chou, T.C. Antiinflammatory and antihyperalgesic activity of C-phycocyanin. Anesth. Analg. 2009, 108, 1303–1310. [Google Scholar]
- Liu, Q.; Huang, Y.; Zhang, R.; Cai, T.; Cai, Y. Medical Application of Spirulina platensis Derived C-Phycocyanin. Evid. Based Complement. Alternat. Med. 2016, 7803846. [Google Scholar]
- Mao, T.K.; Van De Water, J.; Gershwin, M.E. Effect of Spirulina on the secretion of cytokines from peripheral blood mononuclear cells. J. Med. Food. 2000, 3, 135–140. [Google Scholar]
- Mohan, k.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Rajan, D.K. Potential uses of fungal polysaccharides as immunostimulants in fish and shrimp aquaculture: A review. Aquaculture 2019, 500, 250–263. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control | 1% Spirulina | 5% Spirulina | 10% Spirulina |
|---|---|---|---|---|
| Fish meal | 10 | 10 | 10 | 10 |
| Wheat flour | 20 | 19.5 | 18 | 15 |
| Soybean meal | 21.5 | 21 | 19.5 | 18.5 |
| Cottonseed meal | 6.5 | 6.5 | 6 | 6 |
| Mustard cake | 21.5 | 21.5 | 21 | 20 |
| Fish oil | 3 | 3 | 3 | 3 |
| Soya oil | 4 | 4 | 4 | 4 |
| Mineral premix a | 3 | 3 | 3 | 3 |
| Vitamin premix b | 3 | 3 | 3 | 3 |
| Spirulina | 0 | 1 | 5 | 10 |
| cellulose | 4 | 4 | 4 | 4 |
| Salt | 0.5 | 0.5 | 0.5 | 0.5 |
| Molasses | 3 | 3 | 3 | 3 |
| Crude protein (%) | 28 | 28 | 28.5 | 29 |
| Fat | 5.5 | 5.6 | 5.5 | 5.7 |
| Ash | 20.37 | 20.34 | 20.45 | 20.39 |
| Moisture | 12.89 | 12.77 | 12.6 | 12.79 |
| Genes | Primer Sequence | Annealing Temperature (°C) | Product Size | Accession Number |
|---|---|---|---|---|
| tnf-α | GGTGCATACGACCCTGAAGT TTTTGCCTCCATAGGAATCG | 60 | 244 | JQ040498.1 |
| il-8 | ATGAGTCTTAGAGGTCTGGGTG ACAGTGAGGGCTAGGAGGG | 60 | 118 | JN663841 |
| inf-γ | TGCATGTAGGCGGATATCAA GAGGGCGCATAAGTCTGAAG | 60 | 192 | FJ695520.1 |
| actin-β | ACCCACACTGTGCCCATCTACG ATTTCCCTCTCGGCTGTGGTGG | 60 | 146 | JQ991014.1 |
| Parameters | Control | 1% Spirulina | 5% Spirulina | 10% Spirulina |
|---|---|---|---|---|
| Initial body weight (g) | 4.9 ± 0.125 | 4.975 ± 0.182 | 4.65 ± 0.175 | 4.75 ± 0.153 |
| Final body weight (g) | 7.393 ± 0.4362 | 10.19 ± 0.7686 * | 11.11 ± 0.7644 *** | 7.948 ± 0.4511 |
| %-weight gain | 43.67 ± 8.310 | 92.43 ± 17.92 | 117.8 ± 16.32 ** | 89.09 ± 13.07 |
| SGR | 0.2874 ± 0.036 | 0.5035 ± 0.056 | 0.6154 ± 0.037 | 0.4401 ± 0.054 |
| Thermal condition factor | 0.1498 ± 0.020 | 0.2772 ± 0.034 * | 0.3397 ± 0.024 *** | 0.2261 ± 0.028 |
| Survival rate (100%) | 100 | 100 | 100 | 100 |
| Parameters | Control | 1%-Spirulina | 5%-Spirulina | 10%-Spirulina |
|---|---|---|---|---|
| Protease (%) | 51.91 ± 6.14 | 61.86 ± 6.480 | 80.53 ± 6.57 * | 63.17 ± 7.26 |
| Antiprotease (%) | 66.78 ± 14.4 | 120.2 ± 13.84 | 152.2 ± 15.36 ** | 130.6 ± 16.77 * |
| Lysozyme (U/L) | 2.056 ± 0.318 | 4.5 ± 0.606 | 7.00 ± 1.069 *** | 5.33 ± 0.991 * |
| Peroxidase (U/mL) | 0.2611 ± 0.030 | 0.2951 ± 0.028 | 0.4390 ± 0.057 * | 0.248 ± 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faheem, M.; Jamal, R.; Nazeer, N.; Khaliq, S.; Hoseinifar, S.H.; Van Doan, H.; Paolucci, M. Improving Growth, Digestive and Antioxidant Enzymes and Immune Response of Juvenile Grass Carp (Ctenopharyngodon idella) by Using Dietary Spirulina platensis. Fishes 2022, 7, 237. https://doi.org/10.3390/fishes7050237
Faheem M, Jamal R, Nazeer N, Khaliq S, Hoseinifar SH, Van Doan H, Paolucci M. Improving Growth, Digestive and Antioxidant Enzymes and Immune Response of Juvenile Grass Carp (Ctenopharyngodon idella) by Using Dietary Spirulina platensis. Fishes. 2022; 7(5):237. https://doi.org/10.3390/fishes7050237
Chicago/Turabian StyleFaheem, Mehwish, Rafia Jamal, Nadia Nazeer, Saba Khaliq, Seyed Hossein Hoseinifar, Hien Van Doan, and Marina Paolucci. 2022. "Improving Growth, Digestive and Antioxidant Enzymes and Immune Response of Juvenile Grass Carp (Ctenopharyngodon idella) by Using Dietary Spirulina platensis" Fishes 7, no. 5: 237. https://doi.org/10.3390/fishes7050237
APA StyleFaheem, M., Jamal, R., Nazeer, N., Khaliq, S., Hoseinifar, S. H., Van Doan, H., & Paolucci, M. (2022). Improving Growth, Digestive and Antioxidant Enzymes and Immune Response of Juvenile Grass Carp (Ctenopharyngodon idella) by Using Dietary Spirulina platensis. Fishes, 7(5), 237. https://doi.org/10.3390/fishes7050237